
Abstract
The bacterium Photorhabdus luminescens produces a number of insecticidal proteins to kill its larval prey. In this study, we cloned the gene coding for a binary toxin PirA1B1 and purified the recombinant protein using affinity chromatography combined with desalination technology. Furthermore, the cytotoxicity of the recombinant protein against the haemocytes of Galleria mellonella larvae was investigated. We found that the protein had haemocoel insecticidal activity against G. mellonella with an LD50 of 131.5 ng/larva. Intrahaemocoelic injection of PirA1B1 into G. mellonella resulted in significant decreases in haemocyte number and phagocytic ability. In in vitro experiments, PirA1B1 inhibited the spreading behaviour of the haemocytes of G. mellonella larvae and even caused haemocyte degeneration. Fluorescence microscope analysis and visualization of haemocyte F-actin stained with phalloidin-FITC showed that the PirA1B1 toxin disrupted the organization of the haemocyte cytoskeleton. Our results demonstrated that the PirA1B1 toxin disarmed the insect cellular immune system.
Similar content being viewed by others
Introduction
Photorhabdus luminescens, a Gram-negative bacterium, resides as a symbiont in the gut of entomopathogenic nematodes (EPNs) of the genus Heterorhabditis1. Upon entering an insect host, EPNs release the symbiotic bacteria directly into the insect haemocoel. To infect its host and survive, bacteria must be capable of producing a wide range of proteins, including toxins2. To date, four primary classes of toxins are characterized in P. luminescens. The first class, toxin complexes (Tcs), shows both oral and injectable activity against the Colorado potato beetle3. The second class is Photorhabdus Virulence Cassettes (PVCs), and the injection of PVCs destroys insect haemocytes, which undergo dramatic actin cytoskeleton condensation4. Making caterpillars floppy (Mcf), the third class of toxins promotes apoptosis in the midgut and produces a characteristic “floppy” phenotype in the infected insect5. The fourth class, Photorhabdus insect-related toxins (PirAB), are binary toxins that exhibit injectable and oral toxicity against mosquitos and lepidopterans6.
To combat infection, insects rely on multiple immune responses that encompass both humoural and cellular defence reactions. Humoral reactions include the production of antimicrobial peptides (AMPs), reactive oxygen and nitrogen species, and the prophenoloxidase (proPO) activating system that regulates coagulation or melanization of haemolymph7,8,9. Cellular responses include the phagocytosis of small pathogens such as bacteria and fungi and the encapsulation of parasites such as parasitoids and nematodes by haemocytes10,11. The symbiotic bacteria of entomopathogenic nematodes that enter the insect haemocoel must fight with the haemocytes and AMPs. Many toxic proteins produced by symbiotic bacteria are reported to target haemocytes, which affects the host immune system by reducing vitality12 and by inducing apoptosis and cytolysis13,14. Moreover, in a recent study, Tc toxins inhibited the phagocytic activity of haemocytes from Galleria mellonella, and the biologically active components of the Tc toxins were characterized as adenosine diphosphate (ADP)-ribosyltransferases, which modify unusual amino acids and cause actin polymerization15.
PirAB toxin proteins, PirA1B1 and PirA2B2, are encoded at two distinct loci, plu4093 to plu4092 and plu4437 to plu4436, in the TT01 genome, respectively, and these toxins show similarity to both δ-endotoxins from Bacillus thuringiensis and a developmentally regulated protein from the Colorado potato beetle6,16. PirA1B1 has documented oral insecticidal activity against Plutella xylostella, Aedes aegypti, Culex pipiens and Gangya anopheles, and the insecticidal activity is eliminated when pirB1 is knockedout16,17. Moreover, in a recent study, we demonstrated that the PirA2B2 toxin attacks haemocytes and decreases the cellular immunity of G. mellonella larvae following a haemocoel injection of the toxin18. Therefore, whether the PirA1B1 toxins also suppress the immune system of insects is a question to be investigated.
To determine the changes in the immune system of a host at the cellular level and reveal the potential role of PirA1B1 during microbial infection, we analysed the effect of the PirA1B1 toxin on the immune activity of haemocytes in G. mellonella larvae and investigated the most likely mechanisms. Our hope is that this study will reveal the biological role played by PirAB binary toxins in the infection process and determine their potential use in agriculture as alternatives to toxins from Bacillus thuringiensis.
Materials and Methods
Bacterial strains and insects
Escherichia coli DH5α was used as the host for recombinant DNA cloning, and Escherichia coli M15 was used as the host for expression of the PirA1B1 toxin protein. Photorhabdus luminescens TT01 was grown in Luria–Bertani (LB) medium at 28 °C, and the E. coli strains were grown in LB medium at 37 °C.
G. mellonella (Lepidoptera, Pyralidae) larvae were reared on an artificial diet as described in our previous study19. Last instar G. mellonella larvae with a weight of approximately 250–300 mg were used in all experiments.
Cloning, expression and fusion protein solubility analysis
The following primers were used for amplification of PirA1B1 gene based on the genome sequence of P. luminescens TT01: PirA1B1-Antisense: TTTAAGCTTGTGGTGGTGGTGGTGGTGCTCAACTAATTGGTG and PirA1B1-Sense: ATAGGATCCATGCCAGTCAATCAGATTGGCTTAC (the underlined sequences denote the restriction sites). The resultant PCR products were gel purified and ligated into pMD18-T (Takara, Japan). The resultant plasmids were named pMDpirA1B1 and were transformed into E. coli DH5α. Three clones were sampled randomly, and the inserted DNA fragment was sequenced.
The pMDpirA1B1 was digested with BamHI and HindIII (Takara, Japan). The inserted fragment was cloned into expression vector pQE (Qiagen, German), named pQE-pirA1B1, and then transformed into E. coli M15. The pQE vector without an insert fragment was selected as the control, which expressed the 6×His-tagged proteins in the prokaryotic expression system. The inserted DNA fragment was sequenced again. Cells of E. coli M15 harbouring pQE-pirA1B1 were grown in LB medium supplemented with ampicillin (100 mg/ml) at 37 °C to an OD600 nm of 0.6–0.8. Then, isopropyl-β-D-thiogalactopyranoside (IPTG) was added at a concentration of 1 mM to induce the expression of the protein. After IPTG induction for 4 h, aliquots of 1 ml of culture were sampled, and the cells were harvested by centrifugation (10,000 g, 1 min). Pellet were washed 3 times with distilled water and suspended in 0.1 ml of lysis buffer (50 mMNaH2PO4, 300 mM NaCl, 10 mM imidazole, pH 8.0), and then the cells were lysed by sonication (200–300 W, 6 × 10 s,10 s pause) and centrifuged at 10,000 g for 2 min. The supernatant was collected and 10μl aliquots were taken for SDS-PAGE.
Purification of the fusion proteins and western blotting
Soluble proteins were directly purified by nickel affinity chromatography using a MagExtractor His-Tag NPK-700 (Toyobo, Japan), as described by the manufacturer. The resulting E. coli expression samples and the purified proteins were analysed by SDS-PAGE. The protein content was determined using the Bio-Rad Bradford reagent. Western blotting was then performed as described previously18.
Injection and bioassays
Injections were performed directly into the insect haemocoel with a sterilized 50-μl Hamilton syringe. G. mellonella larvae were injected with 10 μl of PBS alone (control) or with the identical volume of PBS containing a designated amount (90, 120, 150, 180 or 240 ng/larva) of toxin protein into the haemocoel of each larva via the last left proleg. After injection, the larvae were reared on an artificial diet. Bioassays were repeated at least three times and were performed on 30 larvae for each concentration.
Phagocytosis assays, total haemocyte counts and actin morphology
For each repeat of the experiment, two groups of G. mellonella larvae were injected with 10 μl of PBS containing 131.5 ng (the 50% lethal dose, LD50) of PirA1B1 or BSA per larva, and a third group were injected with PBS alone and used as a negative control. At 6 and 12 h after injection, fifteen live larvae were sampled from each group for collection of haemolymph. In each group, 40 μl of fresh haemolymph was collected from each larva by pricking the larva with an insect needle. The haemolymph from each individual was divided into two aliquots and processed as previously described for phagocytosis and total haemocyte count assays19.
As targets for phagocytosis, FITC-labelled E. coli cells were used. An aliquot of 30 μl of a haemolymph-anticoagulant solution was added to the well of a 24-well cell culture plate containing a 12-mm-diameter round glass cover slips prefilled with 300 μl of ice-cold Grace’s tissue culture medium (GIM), and incubated at 28 °C for 30 min. After several washes with GIM, the FITC-labelled E. coli cell suspension solution and 380 μl of GIM were added to each well and incubated for 2 h in the dark at 28 °C. Afterwards, the culture medium was removed and 300 μl of a 0.4% trypan blue solution was added to quench the nonphagocytosed bacteria. The phagocytic activity was determined by counting haemocytes with or without ingested bacterial cells under a fluorescence microscope. Five photo-frames for each microscope slide were counted to determine the average.
To count the total haemocyte number, aliquots of the haemolymph suspension were transferred to a Neubauer haemocytometer. Total haemocytes were counted using a phase contrast microscope and expressed as the number of cells per ml of haemolymph.
F-actin staining with phalloidin-FITC was performed according to previous study. Briefly, haemolymph was collected from the larvae injected with PirA1B1 (131.5 ng per larva), BSA (131.5 ng per larva) or PBS at 6 h after injection. An aliquot of 50 μl of diluted haemolymph (20 μl of haemolymph diluted in 30 μl of PBS) from each treatment group was applied to a microscope slide. The slides were placed in a moist chamber for 30 min for haemocytes to attach to the glass surface and then were fixed for 5 min in 3.7% (w/v) formaldehyde in PBS, washed three times in PBS, and then permeabilized with 0.2% Triton X-100 in PBS for 10 min at room temperature. After washing the slides with PBS, haemocyte monolayers were overlaid with phalloidin-FITC (Sigma, USA) at a concentration of 0.05 mg/ml in PBS containing1% dimethyl sulfoxide for 40 min at room temperature in a humidified chamber. Then, the slides were washed with PBS several times and overlaid with a mixture of 30% glycerol and 70% PBS (v/v) and a cover slip. Haemocytes were examined using a fluorescence microscope.
In vitro toxicity experiments of PirA1B1 on haemocytes
The larvae were chilled, surface sterilized and bled into prechilled GIM, and the haemocyte density was adjusted accordingly to 1 × 107 cells/ml. The haemocyte suspension was exposed to 10 μl of PBS containing 131.5 ng of PirA1B1 toxins, BSA (positive control) or PBS alone (negative control). Haemocytes were then incubated in GIM and maintained at 28 °C. After incubation for the designated time (6 and 12 h), cell morphological changes were analysed using an inverted light microscope.
Statistical analyses
All results are expressed as the mean and standard deviation. Data were subjected to analysis of variance (ANOVA), and when the effects of ANOVA were significant, the factors that contributed to the significant differences were determined by means of least significant difference (LSD) tests using the DPS statistical software package. Significant differences were set at P < 0.05.
Results
Recombinant expression of PirA1B1 in E. coli
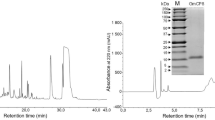
The recombinant plasmid pQE-pirA1B1 was transformed and expressed in E. coli M15. After IPTG induction for 4 h, the whole cell lysate analysed by SDS-PAGE revealed two distinct bands with molecular weights of 45 kDa (PirA1) and 14 kDa (PirB1) (Fig. 1A), which were consistent with the predicted molecular masses. Products were not found in either the uninduced cultures or in the control (lysate of cells transfected with empty vector pQE, data not shown). PirA1B1 was successfully purified using the protocols described (Fig. 1B). The objective band of recombinant PirA1B1 was confirmed by western blot analysis with anti-His6 monoclonal antibody. For the control, no visible reaction band was detected in the group of cell lysates without IPTG induction (Fig. 1C).

(A) SDS-PAGE analysis of PirA1B1 expression in E. coli M15 cells (pQE-pirA1B1). Lane M, markers (KDa); lane 1, uninduced pQE-pirA1B1; lanes 2 and 3, pQE-pirA1B1induced with IPTG. (B) SDS-PAGE analysis of PirA1B1 expressed in E. coli M15 (pQE-pirA1B1) as soluble proteins followed by the purification and desalination protocols described. Lane M, markers (KDa); lanes 1 and 2, unpurified PirA1B1; lane 3, purified and desalinated PirA1B1. (C) Western blot analysis of expressed protein PirA1B1 in E. coli M15 cells (pQE-pirA1B1). Lane M, markers (KDa); lane 1, uninduced pQE-pirA1B1; lanes 2 and 3, pQE-pirA1B1 induced with IPTG.
Insecticidal activity of PirA1B1
PirA1B1 effectively killed G. mellonella larvae, and the mortality rate was dose-dependent based on protein concentrations. The LD50 of PirA1B1 for G. mellonella was 131.5 ng per larva. The larvae developed to a deep brown body colour and showed swelling symptoms after injection with PirA1B1. Larvae of G. mellonella injected with a high dose of PirA1B1 (240 ng per larva) died primarily during the post injection period of 1 to 12 h, with a mortality rate of 100% at 24 h after injection. No dead larvae or external symptoms were observed in the control (injected with PBS; Fig. 2).

Mortality of Galleria mellonella larvae after injection with different doses of PirA1B1 protein.
Injection of PirA1B1 proteins into the larval haemocoel led to decreases in numbers and phagocytic ability of haemocytes
As shown in Fig. 3, only internalized bacteria retained their fluorescence (bright green) after quenching with trypan blue. The phagocytic rate of haemocytes in the group injected with PirA1B1 protein decreased significantly compared with that in the PBS control group (P < 0.05), whereas a significant increase was detected in the BSA treatment group at 6 h after injection (P < 0.05; Figs 3 and 4B).

Differential interference contrast and corresponding fluorescence microscope of Galleria mellonella larvae haemocytes with phagocytised FITC-labelled E. coli cells in vitro. Haemocytes collected from larvae injected with PBS, BSA or PirA1B1 at 6 and 12 h after injection.

Changes in total haemocyte counts and phagocytic rates of Galleria mellonella larvae at 6 and 12 h after injection with PBS, BSA or PirA1B1.
Values for different groups at the identical points in time followed by different letters are significantly different (P < 0.05) according to ANOVA and LSD tests.
There was no significant difference in the total haemocyte counts (THC) between the PBS and BSA treatment groups. However, after injecting PirA1B1 protein into the G. mellonella haemocoel, THC decreased to 58% of the original values after 6 h and ultimately, to 54% of the initial numbers after 12 h (Fig. 4A).
Effect of PirA1B1 protein on the cytoskeletons of the haemocytes of G. mellonella larvae
Staining of haemocytes from PirA1B1-injected larvae with phalloidin-FITC showed distinct differences in actin polymerization compared with the actin-mediated assembly of the cytoskeleton in haemocytes of larvae injected with PBS and BSA. Plasmatocytes in the PBS and BSA control larvae extended their filopodia over the surface of the slide. The granular cells spread to form rounder shapes, and the actin cytoskeleton was largely organized around the periphery of these cells. However, for both plasmatocytes and granular cells in the PirA1B1-injected larvae, the cytoskeleton appeared diffuse and not organized into bundles or stress fibres (Fig. 5).

Light and corresponding fluorescence micrographs of monolayers of haemocytes from Galleria mellonella larvae showing localization of cytoskeletal actin stained with FITC-labelled phalloidin.
Plasmatocytes (PL) and granular cells (GR) from the larvae injected with PBS or BSA showed pseudopods and an organized cytoskeleton; however, no pseudopods and spreading behaviour were observed in either plasmatocytes or granular cells from the larvae injected with PirA1B1.
Toxic effect of PirA1B1 protein on haemocytes of G. mellonella larvae in vitro
The haemocytes treated with PBS and BSA clearly spread normally and all the plasmatocytes were spindle-shaped. By contrast, most PirA1B1-treated haemocytes were small and failed to spread. After incubation for 12 h, the cytotoxic effects of PirA1B1 toxin on haemocytes were more severe, with the dead cells degenerating to form debris in the culture (Fig. 6).

Morphology of Galleria mellonella larvae haemocytes treated with PBS, BSA or PirA1B1 for 6 or 12 h.
Discussion
In this study, the gene coding for the binary toxin PirA1B1 was cloned from P. luminescens and expressed in E.coli M15. The recombinant protein was purified from E. coli M15 using nickel affinity chromatography. The toxin protein had injection activity against G. mellonella with an LD50 of 131.5 ng/larva. In a previous study, the protein PirA2B2 encoded by the loci plu4437 to plu4436 within P. luminescens TT01 had haemocoel insecticidal activity against the fifth instar larvae of both G. mellonella and Spodopteralitura, with an LD50 of 4.0 and 2.8 μg/larva, respectively20. Compared with PirA2B2, PirA1B1 had stronger toxicity against G. mellonella larvae. The causes of the difference in toxicity between PirA2B2 and PirA1B1 are not clear yet. In addition to the unique biological properties of these two toxin proteins, the purification process, which can affect the structure of toxin proteins, was also an important factor worth consideration.
Haemocyte-mediated immunity is activated immediately after the insect haemocoel is penetrated by entomopathogenic nematodes. However, P. luminescens released from the nematode escape the insect cellular response and proliferate successfully in the haemolymph before the insect dies. A successful outcome is primarily attributed to the toxin proteins secreted by the P. luminescens, which fight against the haemocyte-mediated cellular immunity and protect the bacteria from the insect immune responses21,22. Recombinant E. coli clones carrying plu4093–plu4092 (PirA1B1) of TT01 genes show oral activity against both mosquito larvae and the larvae of the moth P. xylostella16. In a recent study, PirAB-fusion protein encoded by plu4093 and plu4092 from P. luminescens TT01 also exhibited injectable insecticidal activity against Spodoptera exigua larvae and had cytotoxicity against insect midgut CF-203 cells23. However, whether the binary toxin PirA1B1 can destroy the immune function of the insect haemocytes is unknown.
Our results showed that injection of PirA1B1 into the haemocoel of G. mellonella larvae significantly decreased the number of circulating haemocytes and ability for phagocytosis. The phagocytic rate and total number of haemocytes decreased to 47% and 54% of their original values at 12 h after the toxin injection, respectively. In a previous study, we also demonstrated that another toxin, PirA2B2, had similar effects on the cellular immunity of G. mellonella larvae18; however, the phagocytic rate decreased to 71% of the PBS control at 12 h after injection and did not change afterwards, which indicated higher toxicity of the toxin PirA1B1 against G. mellonella larvae. Haemocytes play important roles in phagocytosis and capsule formation11,24, and with more haemocytes, insect have greater ability to clear invading pathogens25. Therefore, with the decrease in haemocytes, we believe that the immune strength and immune system coordination were weakened directly. The mechanism by which PirA1B1 causes this reduction in circulating haemocyte number is currently unknown. One possibility is the reduction in haemocyte number due to death and disintegration of haemocytes and/or reduced proliferation of haemocytes26. We performed in vitro experiments and directly treated haemocytes with PirA1B1, and the results showed that the toxin disrupted the spreading behaviour of haemocytes at 6 h after treatment. Furthermore, after incubation for another 12 h, the PirA1B1 toxin caused the haemocytes to lyse, which degenerated to form debris in the culture. We also observed that treatment with the PirA1B1 toxin caused a reduction in haemocyte pseudopod formation. Therefore, based on our results, the binary toxin PirA1B1 was cytotoxic against the haemocytes of G. mellonella larvae. These results were consistent with the in vivo experiments in which haemocoel injection of PirA1B1 caused significant reductions in number of circulating haemocytes in G. mellonella larvae.
To investigate the mechanisms responsible for the inhibition of phagocytosis by haemocytes after PirA1B1 treatment, we performed experiments to determine whether the toxin altered the haemocyte cytoskeleton. Staining the haemocytes with FITC-labelled phalloidin revealed major differences in the cytoskeletal architecture of cells following injection of the toxin protein compared with those in BSA or PBS control larvae. These assays indicated that PirA1B1 adversely affected the cytoskeleton of G. mellonella haemocytes, causing it to become diffuse and disorganized, and this result also correlated with the reduction in formation of pseudopods by the cells. Therefore, we concluded that the toxin PirA1B1 disrupted the cytoskeletons of haemocytes, which led to abnormal haemocyte spreading behaviour and pseudopod formation. Because haemocyte spreading behaviour and the ability to extend pseudopods are essential for phagocytosis27, with their disruption, phagocytosis was suppressed. Thus, PirA1B1 inhibited phagocytosis by affecting the cytoskeleton of the immunocytes. Numerous bacterial toxins reorganize the actin cytoskeleton of target cells. Moreover, Photorhabdus W14 bacterial supernatants cause marked changes in the actin cytoskeleton of specific haemocyte types22. Additionally, the injection of recombinant E. coli expressing Photorhabdus virulence cassettes (PVC) containing cosmids from Photorhabdus destroys insect haemocytes, which undergo dramatic actin cytoskeleton condensation4.
In conclusion, based on our preliminary results, following the injection of PirA1B1 toxin into the haemocoel of G. mellonella larvae, the host haemocyte number decreased and the ability of haemocyte phagocytosis was inhibited by the disruption of the cytoskeleton. Therefore, the toxin PirA1B1 disarmed the insect cellular immune system. As a ubiquitous protein in entomopathogenic nematode symbiotic bacteria, uncovering the cellular immune responses of a host insect to the injection of PirA1B1 helped to understand the interaction between nematode-symbiotic bacteria and their hosts.
Additional Information
How to cite this article: Wu, G. and Yi, Y. Haemocoel injection of PirA1B1 to Galleria mellonella larvae leads to disruption of the haemocyte immune functions. Sci. Rep. 6, 34996; doi: 10.1038/srep34996 (2016).
References
Forst, S., Dowds, B., Boemare, N. & Stackebrandt, E. Xenorhabdus and Photorhabdus spp.: bugs that kill bugs. Annu Rev Microbiol. 51, 47–72 (1997).
ffrench-Constant, R. et al. Photorhabdus: towards a functional genomic analysis of a symbiont and pathogen. FEMSMicrobiol Rev. 26, 433–456 (2003).
Blackburn, M. B., Domek, J. M., Gelman, D. B. & Hu, J. S. The broadly insecticidal Photorhabdus luminescens toxin complex a (Tca): activity against the Colorado potato beetle, Leptinotarsa decemlineata, and sweet potato whitefly, Bemisia tabaci. J Insect Sci. 5, 32 (2005).
Yang, G., Dowling, A. J., Gerike, U., ffrench-Constant, R. H. & Waterfield, N. R. Photorhabdus virulence cassettes confer injectable insecticidal activity against the wax moth. J Bacteriol. 188, 2254–2261 (2006).
Dowling, A. J. et al. The insecticidal toxin Makes caterpillars floppy (Mcf) promotes apoptosis in mammalian cells. Cell Microbiol. 6, 345–353 (2004).
Waterfield, N., Kamita, S. G., Hammock, B. D. & ffrench-Constant, R. The Photorhabdus Pir toxins are similar to a developmentally regulated insect protein but show no juvenile hormone esterase activity. FemsMicrobiol Lett. 245, 47–52 (2005).
Lu, A. et al. Insect prophenoloxidase: the view beyond immunity. Front Physiol. 5, 252 (2014).
Charroux, B. & Royet, J. Drosophila immune response: From systemic antimicrobial peptide production in fat body cells to local defense in the intestinal tract. Fly (Austin) 4, 40–47 (2010).
Giglio, A. et al. Immune challenges trigger cellular and humoral responses in adults of Pterostichus melas italicus (Coleoptera, Carabidae). Arthropod Struct Dev. 44, 209–217 (2015).
Williams, M. J. Drosophila hemopoiesis and cellular immunity. J Immunol. (2007).
Hillyer, J. F., Schmidt, S. L. & Christensen, B. M. Hemocyte-mediated phagocytosis and melanization in the mosquito Armigeres subalbatus following immune challenge by bacteria. Cell Tissue Res. 313, 117–127 (2003).
Khandelwal, P. et al. Insecticidal pilin subunit from the insect pathogen Xenorhabdus nematophila. J Bacteriol. 186, 6465–6476 (2004).
Ribeiro, C., Vignes, M. & Brehelin, M. Xenorhabdus nematophila (enterobacteriacea) secretes a cation-selective calcium-independent porin which causes vacuolation of the rough endoplasmic reticulum and cell lysis. J BiolChem. 278, 3030–3039 (2003).
Vigneux, F. et al. The xaxAB genes encoding a new apoptotic toxin from the insect pathogen Xenorhabdus nematophila are present in plant and human pathogens. J BiolChem. 282, 9571–9580 (2007).
Lang, A. E. et al. Photorhabdus luminescens toxins ADP-ribosylate actin and RhoA to force actin clustering. Science 327, 1139–1142 (2010).
Duchaud, E. et al.The genome sequence of the entomopathogenic bacterium Photorhabdus luminescens.Nat Biotechnol. 21, 1307–1313 (2003).
Ahantarig, A., Chantawat, N., Waterfield, N. R., ffrench-Constant, R. & Kittayapong, P. PirAB toxin from Photorhabdus asymbiotica as a larvicide against dengue vectors. Appl Environ Microbiol. 75, 4627–4629 (2009).
Wu, G., Yi, Y., Sun, J., Li, M. & Qiu, L. No evidence for priming response in Galleria mellonella larvae exposed to toxin protein PirA2B2 from Photorhabdus luminescens TT01: An association with the inhibition of the host cellular immunity. Vaccine 33, 6307–6313 (2015).
Wu, G. Q. & Yi, Y. H. Effects of dietary heavy metals on the immune and antioxidant systems of Galleria mellonella larvae. Comp. Biochem. Phys. C 167 (2015).
Sun, J., Liu, C. & Qiu, L. [Cloning, expression and insecticidal activity of the pirA2B2 gene from Photorhabdus luminescens TT01]. Wei Sheng Wu Xue Bao. 52, 532–537 (2012).
Eleftherianos, I. et al. A single locus from the entomopathogenic bacterium Photorhabdus luminescens inhibits activated Manduca sexta phenoloxidase. FemsMicrobiol Lett. 293, 170–176 (2009).
Au, C., Dean, P., Reynolds, S. E. & ffrench-Constant, R. H. Effect of the insect pathogenic bacterium Photorhabdus on insect phagocytes. Cell Microbiol. 6, 89–95 (2004).
Li, Y. et al. Photorhabdus luminescens PirAB-fusion protein exhibits both cytotoxicity and insecticidal activity. FemsMicrobiol Lett. 356, 23–31 (2014).
Pech, L. L. & Strand, M. R. Plasmatocytes from the moth Pseudoplusia includens induce apoptosis of granular cells. J Insect Physiol. 46, 1565–1573 (2000).
Bergin, D., Brennan, M. & Kavanagh, K. Fluctuations in haemocyte density and microbial load may be used as indicators of fungal pathogenicity in larvae of Galleria mellonella. Microbes Infect. 5, 1389–1395 (2003).
Strand, M. R. The insect cellular immune response. Insect Sci. 15, 1–14 (2008).
Goldenberg, P. Z., Huebner, E. & Greenberg, A. H. Activation of lobster hemocytes for phagocytosis. J InvertebrPathol. 43, 77–88 (1984).
Acknowledgements
The National Natural Science Foundation of Guangdong Province, China (2016A030310301), and the Medical Scientific Research Foundation of Guangdong Province, China (A2016607), supported this research.
Author information
Authors and Affiliations
Contributions
G.W. performed the experiments and wrote the manuscript, and Y.Y. analysed the data and edited the manuscript. All authors have read and approved the manuscript for publication.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Wu, G., Yi, Y. Haemocoel injection of PirA1B1 to Galleria mellonella larvae leads to disruption of the haemocyte immune functions. Sci Rep 6, 34996 (2016). https://doi.org/10.1038/srep34996
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep34996
This article is cited by
-
Dodecanol, metabolite of entomopathogenic fungus Conidiobolus coronatus, affects fatty acid composition and cellular immunity of Galleria mellonella and Calliphora vicina
Scientific Reports (2021)
-
Identification of Arylphorin interacting with the insecticidal protein PirAB from Xenorhabdus nematophila by yeast two-hybrid system
World Journal of Microbiology and Biotechnology (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
