
Abstract
Follicular helper T cells (TFH cells) are specialized effector CD4+ T cells that help B cells develop germinal centers (GCs) and memory. However, the transcription factors that regulate the differentiation of TFH cells remain incompletely understood. Here we report that selective loss of Lef1 or Tcf7 (which encode the transcription factor LEF-1 or TCF-1, respectively) resulted in TFH cell defects, while deletion of both Lef1 and Tcf7 severely impaired the differentiation of TFH cells and the formation of GCs. Forced expression of LEF-1 enhanced TFH differentiation. LEF-1 and TCF-1 coordinated such differentiation by two general mechanisms. First, they established the responsiveness of naive CD4+ T cells to TFH cell signals. Second, they promoted early TFH differentiation via the multipronged approach of sustaining expression of the cytokine receptors IL-6Rα and gp130, enhancing expression of the costimulatory receptor ICOS and promoting expression of the transcriptional repressor Bcl6.
Similar content being viewed by others

Main
The provision of help from T cells to B cells is a critical component of adaptive humoral immunity1,2. During viral infection, the formation of germinal centers (GCs) by antigen-specific B cells requires key signals provided by follicular helper T cells (TFH cells)3, which results in the development of high-affinity long-lived plasma cells and memory B cells4,5. The differentiation of TFH cells begins outside of B cell follicles in a stepwise fashion. Early induction of molecules key to TFH differentiation, such as the transcriptional repressor Bcl6, the chemokine receptor CXCR5, the costimulatory receptor ICOS and the T cell–inhibitory receptor PD-1, occurs in the T cell zone when CD4+ T cells interact with antigen-presenting dendritic cells or other antigen-presenting cells, which then enable migration of the activated CD4+ T cells toward the border of B cell follicles. Upon recognizing cognate antigen-presenting B cells, the differentiating TFH cells migrate deep inside B cell follicles and further differentiate into GC TFH cells as they direct the generation of GC B cells.
The requirement for repeated interactions with antigen-presenting cells is an important feature of the differentiation of TFH cells3, which is presumably connected to maintenance of the activity of critical transcription factors such as Bcl6 (refs. 6,7,8), Batf9, STAT3 (refs. 10,11,12), STAT1 (ref. 10) and Ascl2 (ref. 13) that support such differentiation. Among those, Bcl6 function is absolutely critical. TFH differentiation is completely abrogated in Bcl6−/− CD4+ T cells6,7,8, and ectopic Bcl6 expression in CD4+ T cells leads to augmented TFH differentiation6,9. Various signaling molecules have been identified that can regulate Bcl6 expression in CD4+ T cells14. However, attempts to polarize CD4+ T cells to TFH cells in vitro through the use of interleukin 6 (IL-6) and IL-21 have failed to reproducibly induce the expression of Bcl6 and CXCR5. Therefore, there are clear gaps in the understanding of the molecular requirements for Bcl6 induction and the factors that support TFH differentiation3.
LEF-1 (encoded by Lef1) and TCF-1 (encoded by Tcf7) are transcription factors that contain a conserved high-mobility-group DNA-binding domain. TCF-1 and LEF-1 are known for their essential roles in early T cell development, including specification to the T cell lineage and β-selection during the CD4−CD8− double-negative stage15,16. TCF-1 and LEF-1 critically regulate commitment to the CD4+ T cell lineage versus commitment to the CD8+ T cell lineage upon completion of positive selection of CD4+CD8+ double-positive thymocytes17,18. In mature CD8+ T cells, TCF-1 and LEF-1 regulate the generation, maturation and longevity of memory CD8+ T cells in response to viral or bacterial infection19,20,21. In mature CD4+ T cells, TCF-1 promotes differentiation into the TH2 subset of helper T cells in vitro via positive regulation of the transcription factor GATA-3 (ref. 22). TCF-1 restrains the expression of IL-17A in developing thymocytes and activated CD4+ T cells23. In addition, TCF-1 can interact with the transcription factor Foxp3 and seems to oppose Foxp3-mediated repression of genes in CD4+ regulatory T cells24.
Here we looked for undiscovered regulators of early TFH differentiation and found that LEF-1 and TCF-1 were critical transcriptional regulators of such differentiation. Through the use of a knock-in reporter system and high-throughput sequencing for cDNA (RNA-seq), we found that these transcription factors had high expression in TFH cells after viral or bacterial infection. Deletion of Lef1 or Tcf7 or both in CD4+ T cells led to defects in TFH differentiation in a dose-dependent manner. As a consequence, the magnitude of B cell responses and GC reactions was substantially diminished in mice deficient in LEF-1 and/or TCF-1, after infection. Mechanistically, LEF-1 and TCF-1 regulated multiple interacting mechanisms upstream of Bcl6 to 'preferentially' instruct activated CD4+ T cells to undertake TFH differentiation.
Results
Transcriptional profiles of early TFH cells versus TH1 cells
The initial contact of CD4+ T cells with antigen-presenting cells in the T cell zone can promote the expression of key TFH cell molecules, including Bcl6 and CXCR5. By 72 h into an acute viral infection, the early TFH cells and TH1 cells have become committed to their fate25,26. Early TFH cells are IL-2RαloBcl6hiBlimp1−CXCR5hi, while early TH1 cells are IL-2Rα+ and T-bethiBcl6−Blimp1hi in the context of acute viral or bacterial infection25,26,27,28. To identify additional factors important in the programming of TFH cells, we performed gene-expression analysis of early TFH cells and TH1 cells by RNA-seq. For this we used cells from SMARTA mice (which have transgenic expression of a T cell antigen receptor specific for the lymphocytic choriomeningitis virus (LCMV) gp61 epitope) with the additional modification of replacement of coding sequence in one allele of the endogenous gene Prdm1 (which encodes the transcription factor Blimp1) with sequence encoding yellow fluorescent protein (Blimp1-YFP). We transferred congenically marked (CD45.1+) CD4+ T cells from those mice into C57BL/6 (B6) (CD45.2+) host mice and then acutely infected the host mice with the Armstrong strain of LCMV. We isolated early TFH cells and TH1 cells 3 d after infection and purified the cells to homogeneity by sorting IL-2Rα−Blimp1-YFP− cells and IL-2Rα+Blimp1-YFP+ cells, respectively. We performed RNA-seq on RNA isolated from the cells and obtained transcriptome profiles of early TFH cells and TH1 cells (Fig. 1a,b). Our analysis revealed that approximately 1,200 genes were upregulated more than 1.5-fold in early TFH cells relative to their expression in TH1 cells, and 1,600 genes were downregulated more than 1.5-fold (Fig. 1b). Early TFH cells expressed many genes that are also 'preferentially' expressed by fully differentiated TFH cells and GC TFH cells (Bcl6, Cxcr5, Pdcd1, Pou2af1 and Tnfsf8, among others) and had low expression of many genes repressed in fully differentiated TFH cells and GC TFH cells (Prdm1, Tbx21, IL2ra, Gzmb and Prf1, among others) (Fig. 1a,b). Thus, major attributes of TFH and TH1 cells were transcriptionally well defined by day 3 of an acute viral infection.

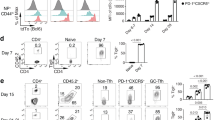
(a) RNA-seq analysis of selected genes of interest in early TFH versus TH1 CD45.1+ Blimp1-YFP SMARTA cells isolated from B6 mice 3 d after transfer of SMARTA cells and infection with LCMV (left half), and of TH1 cells (CXCR5−), TFH cells (PD-1loCXCR5+) and GC TFH cells (PD-1hiCXCR5+) sorted 8 d after LCMV from CD45.2+ B6 mice (right half), presented as high (red) to low (blue) expression. (b) Scatter plot of genes upregulated (red) or downregulated (blue) 1.5-fold or more in early TFH cells relative to their expression in TH1 cells; select genes of interest are labeled. (c) Immunoblot analysis of LEF-1 (two isoforms) and β-actin (loading control) in shCtrl+ and shLef1+ SMARTA cells. (d–f) Frequency of shCtrl+ or shLef1+ CD45.1+ SMARTA cells (Ametrine+CD45.1+CD4+CD19−) among total CD4+ T cells (d) and phenotyping of shCtrl+ and shLef1+ SMARTA cells (e,f) obtained from B6 host mice 3 d after transfer of SMARTA cells infected with shRNAmir-expressing retrovirus, and infection of the hosts with LCMV. Numbers adjacent to outlined areas (e,f) indicate percent Bcl6+CXCR5+ TFH cells (e) or IL-2Rα−CXCR5+ TFH cells (f) among SMARTA cells. Each symbol (d–f) represents an individual mouse (n = 7 per group). *P < 0.05 and **P < 0.001 (Student's t-test). Data are from one experiment with 20 mice and two biological replicates (a,b), are representative of two experiments (c) or are pooled from two independent experiments (d–f; mean ± s.e.m.).
LEF-1 is a transcriptional regulator of TFH differentiation
To further filter the 2,800 gene-expression differences between early TFH cells and TH1 cells, we focused on transcription factors. We performed an additional set of RNA-seq experiments with CD4+ T cells activated in vitro under TH1-polarizing conditions (IL-12 plus antibody to IL-4 (anti-IL-4) plus antibody to transforming growth factor-β) or with IL-6 (IL-6 plus antibody to interferon-γ plus anti-IL-12). We used these screening conditions because in vitro stimulation of CD4+ T cells in the presence of IL-6 resulted in some gene-expression changes associated with TFH differentiation (Supplementary Fig. 1a–c). Most notably, Il21 was robustly induced by IL-6; however, we did not detect major aspects of TFH cell biology in IL-6-stimulated CD4+ T cells, such as expression of CXCR5 protein or sustained expression of Bcl6 (refs. 3,13,29,30) (Supplementary Fig. 1f). This outcome suggested that key transcriptional regulators required for TFH differentiation were not induced under IL-6 conditions in vitro. We next performed a comparative analysis of gene-expression differences between the early TFH cells generated in vivo and the CD4+ T cells stimulated in vitro with IL-6. To find critical previously unidentified early upstream transcriptional regulators of TFH differentiation, we focused on genes that met two conditions: 'preferential' expression by early TFH cells in vivo and lack of a difference in expression after in vitro stimulation with IL-6, relative to expression after stimulation without IL-6. Lef1 satisfied these two conditions (Fig. 1b and Supplementary Fig. 1d,g), and we selected it for further analysis in part because LEF-1 is required for the formation of memory CD8+ T cells20 and there are similarities between the differentiation of TFH cells and that of memory CD8+ T cells25,31.
When expressed in SMARTA CD4+ T cells, a retroviral vector expressing microRNA-adapted short hairpin RNA (shRNAmir) targeting Lef1 (shLef1) inhibited expression of both isoforms of LEF-1 protein (Fig. 1c). To determine whether the early differentiation of TFH cells in vivo was dependent on LEF-1, we transferred SMARTA CD45.1+ CD4+ T cells expressing control shRNAmir (shCtrl) targeting Cd19 (a gene not expressed in CD4+ T cells) or shLef1+ SMARTA CD4+ T cells into B6 mice. At 3 d after infection of recipient mice with LCMV, shLef1+ SMARTA CD4+ T cells produced approximately half as many early TFH cells as did shCtrl+ SMARTA CD4+ T cells, as assessed by flow cytometry with phenotyping of either Bcl6+CXCR5+ cells (Fig. 1e) or IL-2Rα−CXCR5+ cells (Fig. 1f). The effect of the knockdown of LEF-1 was selective to TFH differentiation, as the activation of SMARTA CD4+ T cells (as assessed by upregulation of expression of the activation marker CD44; data not shown) and proliferation (Fig. 1d) were similar for shCtrl+ CD4+ T cells and shLef1+ CD4+ T cells. The reduced TFH differentiation of shLef1+ CD4+ T cells indicated that LEF-1 might be an important and dose-limiting contributor to this process.
LEF-1 controls TFH differentiation and GC formation
We next investigated whether LEF-1 function in CD4+ T cells was important for GC TFH differentiation and GC reactions. We transferred shLef1+ or shCtrl+ SMARTA CD4+ T cells into B6 mice and analyzed the recipient mice 8 d after acute infection with LCMV. The activation and proliferation of CD4+ T cells were not affected by reduced Lef1 expression, compared with that of shCtrl+ CD4+ T cells (Fig. 2a), but the TFH differentiation of shLef1+ cells was impaired (Fig. 2b,c). The TFH-differentiation defect of shLef1+ cells was less severe at day 8 than that observed on day 3 (Fig. 2b), potentially due to the fact that sustained gene knockdown in CD4+ T cells in vivo is difficult to accomplish under conditions of rapid proliferation. We observed milder TFH-differentiation defects for most retrovirus-expressed shRNAmirs, including shRNAmir directed against Bcl6, at peak proliferation time points than at early time points after infection (data not shown). Nevertheless, shLef1+ SMARTA CD4+ T cells showed defective differentiation into GC TFH cells, identified here as PSGL-1loCXCR5+ T cells (Fig. 2d) or Bcl6+CXCR5+ T cells (Fig. 2e), compared with such differentiation of shCtrl+ SMARTA CD4+ T cells. As a result, the development of GC B cells (Bcl6+CD19+) was moderately impaired in the presence of shLef1+ SMARTA CD4+ T cells relative to their development in the presence of shCtrl+ cells (Fig. 2f). Thus, a reduction in LEF-1 expression in CD4+ T cells resulted in a loss of TFH cells and GC TFH cells and a proportional loss of GC B cells during an immune response to LCMV.

(a,b) Abundance of shCtrl+ or shLef1+ CD45.1+ SMARTA cells (Ametrine+CD45.1+CD4+CD19−) among total CD4+ T cells (a) and phenotyping of shCtrl+ and shLef1+ SMARTA cells (b), assessed by flow cytometry at 8 d after transfer of shLef1+ or shCtrl+ SMARTA cells into B6 host mice and infection of the hosts with LCMV. Numbers adjacent to outlined areas (b, left) indicate percent SLAMloCXCR5+ TFH cells among SMARTA cells. (c) Expression of CXCR5 on shCtrl+ or shLef1+ SMARTA cells from mice as in a,b. MFI, mean fluorescence intensity. (d,e) Phenotyping of shCtrl+ and shLef1+ SMARTA GC TFH cells from mice as in a,b. Numbers adjacent to outlined areas (left) indicate percent PSGL-1loCXCR5+ GC TFH cells (d) or Bcl6hiCXCR5+ GC TFH cells (e) among SMARTA cells. (f) Phenotyping of B cells from mice as in a,b. Numbers adjacent to outlined areas (left) indicate percent Bcl6+ CD19+ GC B cells among total B cells. Each symbol represents an individual mouse (n = 4–5 per group). *P < 0.05 (Student's t-test). Data are representative of two independent experiments (mean ± s.e.m.).
Ablation of Lef1 diminishes GC TFH differentiation
We next investigated the role of LEF-1 in TFH differentiation through the use of mice with conditional deletion of Lef1. Lineage-specific deletion of loxP-flanked Lef1 alleles (Lef1fl/fl) in thymocytes through the use of Cre recombinase expressed from the T cell–specific Cd4 promoter (Cd4-Cre) impairs CD4+ T cell lineage 'choice' and diminishes the output of CD4+ T cells18. To avoid this, we used mice with transgenic expression of Cre driven by the promoter of the human gene encoding the activation-costimulation molecule CD2 (hCD2-Cre), which results in gene ablation in mature T cells32. We also crossed mice to mice expressing an allele for the expression of green fluorescent protein (GFP) from the ubiquitously expressed 'Rosa26' locus (Rosa26-STOP-GFP; called 'Rosa26GFP' here). As marked by GFP expression due to excision of the loxP-flanked transcription-translation 'stop' sequence from the Rosa26GFP allele, over 70% of splenic CD4+ T cells in Rosa26GFPhCD2-Cre+ mice were GFP+, whereas less than 15% of CD4+ thymocytes were GFP+ (Supplementary Fig. 2a). We crossed Rosa26GFPhCD2-Cre+ mice to the Lef1fl/fl strain to generate Rosa26GFPLef1fl/flhCD2-Cre+ mice (called 'Lef1−/− mice' here). Both isoforms of LEF-1 were completely ablated in GFP+ CD4+ T cells from Lef1−/− mice (Supplementary Fig. 2b). Late deletion of LEF-1 did not detectably affect thymocyte development or cause aberrant activation of mature T cells (Supplementary Fig. 2d,f,h,i) but reduced total thymic cellularity by approximately 15% and mature CD4+ T cells by approximately 25% (Supplementary Fig. 2e,g). To determine the effect of LEF-1 deficiency in CD4+ T cells on TFH differentiation, we infected Lef1−/− mice and their control littermates (Lef1+/flhCD2-Cre− or Lef1+/+hCD2-Cre+) with vaccinia virus and assessed the presence of CD44hiCD62L− activated GFP+ CD4+ splenic T cells on day 8 after infection. The frequency of TH1 cells (SLAMhiCXCR5−) was similar in Lef1−/− mice and their control littermates, although the absolute number of SLAMhiCXCR5− TH1 cells was modestly lower in Lef1−/− mice than in their control littermates (P = 0.51; Supplementary Fig. 3), consistent with the slightly smaller CD4+ T cell compartment in uninfected Lef1−/− mice than in their uninfected control littermates (Supplementary Fig. 2g). In contrast, the number of SLAM−CXCR5+ TFH cells was more markedly diminished in vaccinia virus–infected Lef1−/− mice compared with the number of these cells in their infected control littermates (Fig. 3a). In particular, the number of GC TFH cells was considerably lower in Lef1−/− mice than in their control littermates (by Bcl6+CXCR5+ and PD-1hiCXCR5+ phenotyping; Fig. 3b,c). These data further supported the proposal of role for LEF-1 in directing the differentiation of TFH cells.

Flow cytometry of cells from the spleen of Lef1−/− mice and their control littermates (Ctrl) 8 d after infection with vaccinia virus. Numbers adjacent to outlined areas (left) indicate percent SLAMloCXCR5+ TFH cells (a), Bcl6+CXCR5+ GC TFH cells (b) or PD-1hiCXCR5+ GC TFH cells (c) among cells gated as CD44hiCD62LloGFP+CD4+ T cells. Each symbol represents an individual mouse. *P < 0.01 and **P < 0.001 (Student's t-test). Data are pooled from four independent experiments (mean ± s.d.).
TCF-1 expression is retained in TFH cells but not in TH1 cells
RNA-seq analysis of early TFH cells and TH1 cells isolated from B6 mice revealed that Tcf7 also had high expression in early TFH cells, but Tcf7 was not induced by in vitro stimulation of CD4+ T cells with IL-6 (Fig. 1b and Supplementary Fig. 1e,g). Given that LEF-1 and TCF-1 are related transcription factors, we investigated whether TCF-1 was also an early regulator of TFH differentiation. For this purpose, we generated mice with sequence encoding GFP inserted into the Tcf7 locus (Tcf7GFP; Supplementary Fig. 4a). The Tcf7-GFP reporter had abundant expression in CD4+ T cells, CD8+ T cells and CD4+CD25+ regulatory T cells but was absent in B220+ cells (Supplementary Fig. 4b–d), which demonstrated the reporter fidelity. The expression of Tcf7-GFP was highest in CD44loCD62L+ naive T cells but was moderately diminished in antigen-experienced T cell subsets such as CD44hiCD62L+ memory-phenotype T cells, and particularly CD44hiCD62L− effector-phenotype T cells (Supplementary Fig. 4b,c). To analyze TCF-1 expression kinetics in antigen-specific CD4+ T cells, we generated Tcf7GFP/+ SMARTA mice and adoptively transferred naive CD44loCD62L+ CD45.2+ CD4+ T cells from those mice into CD45.1+ congenic recipients. Following infection with LCMV, Tcf7-GFP expression was greatly diminished in SLAMhi CXCR5− TH1 cells relative to its expression in naive CD4 T cells by day 8 after infection, while Tcf7-GFP expression was maintained at a high level by most SLAMloCXCR5+ TFH cells (Fig. 4a).

(a) Flow cytometry analyzing expression of the Tcf7-GFP reporter (right) in TH1 (CXCR5−SLAMhi) and TFH (CXCR5+SLAMlo) donor (CD45.2+) SMARTA CD4+ T cells (gated as outlined at left) 8 d after transfer of Tcf7GFP/+ SMARTA cells into CD45.1+ recipients and infection of the hosts with LCMV. Numbers above bracketed lines (right) indicate percent Tcf7-GFP+ cells. (b–f) Flow cytometry of splenic T cells (b–d), GC B cells (e) and plasma cells (f) from Tcf7−/− and Lef1−/−Tcf7−/− mice and their control littermates (Ctrl) 8 d after intravenous infection with vaccinia virus. Numbers adjacent to outlined areas (left) indicate percent SLAMloCXCR5+ TFH cells (b), Bcl6+CXCR5+ GC TFH cells (c) or PD-1hiCXCR5+ GC TFH cells (d), gated on CD44hiCD62L−GFP+CD4+ T cells in spleen, or percent GL7+Fas+ GC B cells (e) or IgDloCD138+ plasma cells (f). Each symbol (right) represents an individual mouse. *P < 0.05, **P < 0.01 and ***P < 0.001 (Student's t-test). Data are representative of two or more experiments (a) or are from three or more experiments (b–f; mean ± s.d.).
We next investigated whether the retention of TCF-1 expression was associated with the TFH-differentiation program in response to other in vivo stimuli. Following adoptive transfer of Tcf7GFP SMARTA CD4+ T cells, we infected recipient mice with Listeria monocytogenes expressing the gp61 epitope of LCMV. In other experiments, we directly infected Tcf7GFP/+ mice with vaccinia virus, as a second viral infection model. Whereas SLAMhiCXCR5− TH1 cells that developed in both systems downregulated Tcf7-GFP expression, SLAMloCXCR5+ TFH cells generated in response to both the bacterial and viral infections retained high expression of Tcf7-GFP (Supplementary Fig. 4e,f). Given that TCF-1 is known to be markedly downregulated in effector CD8+ T cells33, these observations indicated that retention of TCF-1 expression at the effector phase of a T cell response was unique to TFH cells and further suggested a possible requirement for TCF-1 in TFH differentiation.
Both LEF-1 and TCF-1 are essential for TFH cell responses
To address the role of TCF-1 in TFH cells, we generated Rosa26GFPTcf7fl/flhCD2-Cre+ mice (called 'Tcf7−/− mice' here), in which all isoforms of TCF-1 were ablated in GFP+ CD4+ T cells (Supplementary Fig. 2c). To investigate the functional redundancy between LEF-1 and TCF-1, we also crossed Tcf7−/− with Lef1−/− mice to generate Lef1−/−Tcf7−/− mice (Rosa26GFPLef1fl/flTcf7fl/flhCD2-Cre+). Similar to Lef1−/− mice, Tcf7−/− mice and Lef1−/−Tcf7−/− mice did not have T cell–development defects or aberrant activation of mature T cells in (Supplementary Fig. 2). Although we observed slightly less thymic and splenic cellularity in Tcf7−/− mice than in their control littermates (Lef1+/flTcf7+/flhCD2-Cre− or Lef1+/+Tcf7+/+hCD2-Cre+), this difference was not evident in Tcf7−/− or Lef1−/−Tcf7−/− mice (Supplementary Fig. 2d,f,h,i). We assessed the CD4+ T cell responses of Lef1−/−Tcf7−/− mice in response to infection with vaccinia virus. On day 8 after infection, analysis of CD44hiCD62L− activated GFP+ CD4+ T cells revealed that the frequency and number of SLAMloCXCR5+ TFH cells were diminished in Tcf7−/− mice compared with that of control mice (Fig. 4b), with a comparable reduction in GC TFH cells (Bcl6+CXCR5+ and PD-1hiCXCR5+ phenotyping; Fig. 4c,d). We found greater defects in Lef1−/−Tcf7−/− mice than in Tcf7−/− mice (Fig. 4b–d), which indicated that both LEF-1 and TCF-1 contributed to regulating the differentiation of TFH cells .
Consistent with the observations reported above, Tcf7−/− and Lef1−/−Tcf7−/− mice exhibited a significantly lower frequency and number of GL7+Fas+ GC B cells than that of control mice (Fig. 4e), with the most severe GC B cell defect in Lef1−/−Tcf7−/− mice (Fig. 4e). The number of IgDloCD138+ plasma cells was moderately reduced in Tcf7−/− mice but was severely compromised in Lef1−/−Tcf7−/− mice, relative to that in their control littermates (Fig. 4f). As a result, the production of vaccinia virus–specific antibodies was significantly impaired in Lef1−/−Tcf7−/− mice compared with that of their control littermates (P = 0.017; Supplementary Fig. 5). In summary, our data indicated critical roles for LEF-1 and TCF-1 in TFH differentiation and, consequently, B cell–helping functions, in a CD4+ T cell–intrinsic manner.
Ectopic Lef1 expression augments TFH differentiation
We next investigated whether enhanced expression of one of these transcription factors (LEF-1 and TCF-1) could augment the TFH differentiation of antigen-specific CD4+ T cells. Given that LEF-1 and TCF-1 exhibited overlapping activities in instructing the differentiation of TFH cells, we assessed the TFH differentiation of CD4+ T cells after ectopic expression of LEF-1. LEF-1 can be expressed as two isoforms in CD4+ T cells due to differential promoter use (Fig. 1c), with the full-length isoform containing an amino-terminal β-catenin-binding domain. We constructed a retrovirus expressing full-length Lef1 (Lef1-RV) and confirmed increased expression of LEF-1 in Lef1-RV+ SMARTA CD45.1+ CD4+ T cells by flow cytometry (Fig. 5a) and immunoblot analysis (data not shown). We infected CD45.1+ SMARTA CD4+ T cells with control retrovirus expressing GFP alone (GFP-RV) or Lef1-RV and transferred the cells into B6 mice, which we then infected with LCMV. The overall activation and proliferation of Lef1-RV+ CD4+ T cells was normal compared with that of GFP-RV+ CD4+ T cells (Fig. 5b and data not shown). Ectopic LEF-1 expression resulted in enhanced TFH development of Lef1-RV+ cells relative to that of GFP-RV+ cells at 8 d after infection (Fig. 5c). Moreover, we found that Lef1-RV+ TH1 cells (SLAMhiCXCR5−) unexpectedly exhibited higher expression of the canonical TFH molecules CXCR5 (Fig. 5d) and PD-1 (Fig. 5e) than that of their GFP-RV+ counterparts. Most notably, GC TFH cells (with a phenotype of either PSGL-1loCXCR5+ or PD-1hiCXCR5+) developed at a significantly higher frequency among Lef1-RV+ SMARTA CD4+ T cells than among their GFP-RV+ counterparts (Fig. 5f,g).

(a) Flow cytometry analyzing the expression of LEF-1 in GFP-RV+ and Lef1-RV+ SMARTA cells. Ctrl, isotype-matched control antibody. (b) Frequency of GFP-RV+ or Lef1-RV+ (RV+) SMARTA cells (GFP+CD45.1+CD4+CD19−) among total CD4+ T cells, assessed by flow cytometry at 8 d after transfer of SMARTA cells into B6 mice (CD45.2+) and infection of the hosts with LCMV. (c) Phenotype of GFP-RV+ or Lef1-RV+ SMARTA cells as in b. Numbers adjacent to outlined areas (left) indicate percent SLAMloCXCR5+ TFH cells among GFP-RV+ or Lef1-RV+ SMARTA cells. (d,e) Expression of the canonical TFH cell markers CXCR5 (d) and PD-1 (e) on CXCR5− TH1 cells and CXCR5+ TFH cells among GFP-RV+ and Lef1-RV+ cells as in b, normalized to the mean fluorescence intensity of GFP-RV+ cells in each group. (f,g) Phenotype of GC TFH cells from mice as in b. Numbers adjacent to outlined areas (left) indicate percent PSGL-1loCXCR5+ GC TFH cells (f) or PD-1hiCXCR5+ GC TFH cells (g) among GFP-RV+ or Lef1-RV+ SMARTA cells. Each symbol represents an individual mouse (n = 9 per group). *P < 0.01 and **P < 0.001 (Student's t-test). Data are pooled from two independent experiments (mean ± s.e.m.).
LEF-1 enhances expression of IL-6 receptors and ICOS
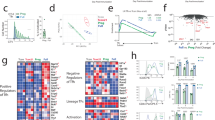
To gain insight into how LEF-1 regulates TFH differentiation, we performed RNA-seq analysis of GFP-RV+ or Lef1-RV+ CXCR5lo TH1 and CXCR5hi TFH SMARTA CD4+ T cells. We next used the transcriptional signatures of TFH and GC TFH cells and gene-set–enrichment analysis (GSEA) to investigate whether Lef1-RV+ TH1 cells showed enrichment for expression of these gene signatures compared with their expression in control (GFP-RV+) TH1 cells. We found substantial enrichment for expression of the TFH cell and GC TFH cell gene signatures (Supplementary Table 1) in TH1 cells constitutively expressing Lef1 (normalized enrichment score, 1.21 (TFH cells) or 1.29 (GC TFH cells); Fig. 6a) compared with their expression in control TH1 cells. Detailed examination revealed that the expression of Il6ra, Il6st, Bcl6, Cxcr5, Slamf6 and Pou2af1 was particularly different in Lef1-RV+ TH1 cells than in GFP-RV+ TH1 cells (Fig. 6b).

(a) GSEA of the TFH cell gene signature (left) and GC TFH cell gene signature (right) in Lef1-RV+ TH1 cells relative to the expression in GFP-RV+ TH1 cells, by RNA-seq analysis of GFP-RV+ and Lef1-RV+ SMARTA cells (CD45.1+CD4+CD19−) isolated from B6 mice 4 d after transfer of SMARTA cells and infection of hosts with LCMV, then sorted into populations of CXCR5+ TFH cells or CXCR5− TH1 cells. (b) Heat map of selected genes upregulated in Lef1-RV+ TH1 cells relative to their expression in GFP-RV+ TH1 cells (high (red) to low (blue)), from mice as in a. (c,d) Expression of IL-6Rα (c) and gp130 (d) on GFP-RV+ or Lef1-RV+ SMARTA cells (CD45.1+CD4+CD19−), assessed by flow cytometry 3 d after transfer of SMARTA cells into B6 host mice (CD45.2+) and infection of the hosts with LCMV. (e,f) Expression of IL-6Rα (e) and gp130 (f) on GFP-RV+ or Lef1-RV+ TH1 and TFH cells from mice as in c,d. (g,h) Expression of ICOS on total RV+ SMARTA cells (g) or on the CXCR5+ TFH and CXCR5− TH1 cell subpopulations (h) from mice as in c,d. Each symbol (c–h) represents an individual mouse (n = 10–14 per group). *P < 0.05, **P < 0.01 and ***P < 0.001 (Student's t-test). Data are from one experiment with eight mice in each and two biological replicates (a,b) or are pooled from three experiments (c–h; mean ± s.e.m.).
Given the induction of both Il6ra and Il6st (which encode the IL-6Rα and gp130 receptors for IL-6, respectively) in Lef1-RV+ TH1 cells and the fact that signaling via IL-6 receptors is one of the earliest signals that instruct TFH differentiation3, we investigated whether LEF-1-augmented TFH differentiation might be mediated through enhanced surface expression of IL-6Rα and gp130. We analyzed the expression of IL-6Rα and gp130 on the surface of Lef1-RV+ or GFP-RV+ SMARTA CD4+ T cells at day 3 after infection with LCMV, a time when signaling via IL-6 receptors is known to be critical for TFH differentiation10. The ectopic expression of LEF-1 in Lef1-RV+ SMARTA CD4+ T cells resulted in higher expression of IL-6Rα than that on GFP-RV+ SMARTA CD4+ T cells (Fig. 6c). In a comparison of IL-6Rα expression on naive CD4+ T cells and that on activated Lef1-RV+ or GFP-RV+ SMARTA CD4+ T cells, we found that overexpression LEF-1 reduced the downregulation of IL-6Rα expression observed on activated GFP-RV+ CD4+ T cells (Fig. 6c). Overexpression of LEF-1 had a similar effect on gp130, reducing the downregulation of gp130 expression observed on activated GFP-RV+ CD4+ T cells (Fig. 6d). We then assessed the expression of IL-6Rα and gp130 on TFH and TH1 subpopulations. We observed modestly higher IL-6Rα expression on TFH cells, whereas Lef1-RV+ TH1 cells expressed >150% more IL-6Rα than did GFP-RV+ TH1 cells (Fig. 6e). While the expression of gp130 was only moderately higher on total Lef1-RV+ SMARTA CD4+ T cells than on GFP-RV+ SMARTA CD4+ T cells (Fig. 6d), gp130 expression was 'preferentially' upregulated on Lef1-RV+ TH1 cells compared with its expression on GFP-RV+ TH1 cells (Fig. 6f).
RNA-seq analysis also revealed that Icos expression was upregulated in Lef1-RV+ TH1 cells compared with its expression in GFP-RV+ TH1 cells (Fig. 6b). Because ICOS has essential roles during both early stages and late stages of TFH differentiation26, we further assessed ICOS expression. Expression of ICOS protein was higher on Lef1-RV+ T cells than on GFP-RV+ cells (Fig. 6g), and its upregulation occurred predominantly on Lef1-RV+ TH1 cells (Fig. 6h), to levels comparable to those on GFP-RV+ TFH cells. These observations indicated that LEF-1 functioned to help CD4+ T cells retain surface expression of IL-6 receptors and upregulate ICOS expression to enhance the responsiveness of activated CD4+ T cells to signaling via IL-6 and the ligand for ICOS, two essential signals for early TFH differentiation.
We then investigated whether overexpression of LEF-1 could restore TFH differentiation in the absence of Bcl6. Bcl6fl/flCd4-Cre CD4+ T cells fail to differentiate into TFH cells during acute viral infection or immunization with protein34. Lef1-RV+ or GFP-RV+ Bcl6fl/flCd4-Cre SMARTA CD4+ T cells transferred into B6 mice failed to differentiate into TFH cells in vivo at day 8 after infection of the recipient mice with LCMV (Supplementary Fig. 6). These results indicated that LEF-1-mediated regulation of the IL-6 receptor complex and ICOS expression acted upstream of Bcl6 expression early in TFH differentiation.
Extensive gene-regulation defects in Lef1−/−Tcf7−/− GC TFH cells
We further assessed the requirements for LEF-1 and TCF-1 in the expression of key TFH cell molecules by transcriptomic analysis of Lef1−/−Tcf7−/− GC TFH cells. We performed RNA-seq analysis of total RNA extracted from GC TFH cells (sorted as PD-1hiCXCR5+ cells among CD44hiCD62LloGFP+CD4+ T cells) isolated from Lef1−/−Tcf7−/− and control mice (Lef1+/flTcf7+/flhCD2-Cre− or Lef1+/+Tcf7+/+hCD2-Cre+) on day 8 after infection with vaccinia virus. We found that 306 genes were downregulated and 668 genes were upregulated in Lef1−/−Tcf7−/− GC TFH cells relative to their expression in control GC TFH cells (false-discovery rate, <0.01; change in expression, ≥1.5-fold; Fig. 7a). In line with the enhanced expression of Il6st and Icos induced by overexpression of LEF-1, Lef1−/−Tcf7−/− GC TFH cells had a much lower abundance of Il6st and Icos transcripts than did control cells (Fig. 7b). Flow cytometry showed lower expression of gp130 and ICOS on Lef1−/−Tcf7−/− CXCR5+ TFH cells than on control TFH cells (Fig. 7c,d). Although the decrease in Il6ra mRNA in Lef1−/−Tcf7−/− GC TFH cells did not reach statistical significance in the transcriptomic analysis, expression of IL-6Rα protein was consistently lower on Lef1−/−Tcf7−/− CXCR5+ TFH cells than on control TFH cells (Fig. 7e). These observations indicated essential and overlapping roles for both LEF-1 and TCF-1 in supporting the expression of IL-6 receptors and ICOS during TFH differentiation.

(a) RNA-seq analysis of genes upregulated (red) or downregulated (blue) 1.5-fold or more (green diagonal lines indicate expression boundaries) in PD-1hiCXCR5+ GC TFH cells sorted from the spleen of Lef1−/−Tcf7−/− mice relative to their expression in such cells from their control littermates (Ctrl) 8 d after infection with vaccinia virus; select genes of interest are labeled. (b) Selected genes (right margins) regulated differentially in GC TFH cells from Lef1−/−Tcf7−/− mice relative to their regulation in cells from their control littermates. (c–e) Expression of gp130 (c), ICOS (d) and IL-6Rα (e) by CXCR5+ TFH and CXCR5− TH1 cells from Lef1−/−Tcf7−/− mice and their control littermates 8 d after intravenous infection with vaccinia virus, analyzed by flow cytometry; results (right) were normalized to the mean fluorescence intensity for control TFH cells. Numbers in plots (left) indicate mean fluorescence intensity. Each symbol represents an individual mouse (n = 5–9 per group). (f) Quantitative RT-PCR analysis of Ascl2 and Prdm1 in CXCR5− TH1 cells, PD-1loCXCR5+ TFH cells and PD-1hiCXCR5+ GC TFH cells sorted from Lef1−/−Tcf7−/− mice and their control littermates 8 d after intravenous infection with vaccinia virus; results were normalized to those of control TFH cells. ND, not reliably detected. *P < 0.05, **P < 0.01 and ***P < 0.001 (Student's t-test). Data are from one experiment (a,b), four independent experiments (c–e; mean ± s.d.) or two experiments with each sample measured in duplicate (f; mean ± s.d.).
The abundance of Bcl6 transcripts was lower in PD-1hiCXCR5+ GC TFH cells from Lef1−/−Tcf7−/− mice than in those from control mice, while the expression of Prdm1 was substantially elevated in Lef1−/−Tcf7−/− GC TFH cells (Fig. 7b). Bcl6 and Blimp1 are known to have mutually antagonistic roles during TFH differentiation6. Blimp1 directly inhibits Bcl6 expression and is a potent inhibitor of TFH differentiation6,28,30. We confirmed the enhanced expression of Prdm1 in Lef1−/−Tcf7−/− PD-1hiCXCR5+ GC TFH by quantitative PCR (Fig. 7f). This increase was specific to GC TFH cells (PD-1hiCXCR5+) and TFH cells (PD-1loCXCR5+), because TH1 cells (CXCR5−) from Lef1−/−Tcf7−/− mice and control mice had similar expression of Prdm1 (Fig. 7f). The transcription factor Ascl2 is important in TFH differentiation13. Ascl2 expression was lower in Lef1−/−Tcf7−/− GC TFH cells than in control cells, but this reduction was less pronounced in PD-1loCXCR5+ TFH cells (Fig. 7f). Expression of Rorc (which encodes the transcription factor RORγt) and Il17a was almost completely absent in control GC TFH cells, but these genes were expressed in Lef1−/−Tcf7−/− GC TFH cells (Fig. 7b). Although the expression of genes characteristic of TH17 cells is not normally observed after infection with vaccinia virus, our observations were in line with the known role of TCF-1 in restraining TH17 differentiation23 and indicated that LEF-1 and TCF-1 might suppress alternative helper T cell fates during TFH differentiation, perhaps in conjunction with Bcl6, which is also known to suppress alternative cell fates3. Other transcriptional changes observed in Lef1−/−Tcf7−/− GC TFH cells compared with the transcription in control GC TFH cells included differential expression of genes encoding transcription factors of the POU family (decreased expression of Pou2af1 and Pou6f1, and increased expression of Pou3f1 and Pou5f1) and key molecules of the Notch signaling pathway (decreased expression of Hes5 and Psen2, and increased expression of Rbpj) (Fig. 7b). The role of these factors in TFH cells remains to be investigated. Overall, these observations suggested that LEF-1 and TCF-1 contributed to the regulation of many genes in activated, antigen-specific CD4+ T cells in vivo, including the positive regulation of Bcl6 and repression of Blimp1 to induce TFH differentiation.
Direct binding of TCF-1 to key TFH cell–associated genes
We used chromatin immunoprecipitation (ChIP) followed by deep sequencing (ChIP-seq) to determine whether LEF-1 and TCF-1 directly regulated the differentially expressed genes identified above. Both TCF-1 and LEF-1 have a highly homologous high-mobility-group DNA-binding domain that recognizes the same DNA consensus motif. Because reagents used for ChIP analysis of TCF-1 are of substantially higher quality than those available for such analysis of LEF-1, we focused on identifying TCF-1-bound genes in TFH cells. Because most TFH cells retained TCF-1 expression similar to that of naive CD4+ T cells (Fig. 4a), we used our ChIP-seq data for TCF-1 that we obtained with naive wild-type CD4+ T cells (data not shown) as a reference for the identification of potential DNA-binding sites for TCF-1. We observed enrichment for binding of TCF-1 at the transcription start site (TSS) of IL6st, the TSS of Bcl6, a region 2.8 kilobases upstream of the Bcl6 TSS (–2.8 kb) and intron 3 of Prdm1 in naive CD4+ T cells, relative to its binding in the majority of the genome, but it was not associated with Il6ra or Ascl2 (Supplementary Fig. 7a). We then performed ChIP analysis of TCF-1 in wild-type and Tcf7−/− naive CD4+ T cells to ensure binding specificity. As a positive control, TCF-1 bound to the TSS of Axin2, a well-characterized TCF-1-responsive gene15, in wild-type naive CD4+ T cells, and this binding was completely abrogated in Tcf7−/− naive CD4+ T cells (Fig. 8a). In addition, TFH cells (CXCR5+) from B6 mice infected with vaccinia virus showed enrichment for the binding of TCF-1 to Axin2 relative to its binding in TH1 cells (CXCR5−) from such mice (Fig. 8a), consistent with higher expression of TCF-1 protein in TFH cells than in TH1 cells. TCF-1 bound to Il6st in wild-type naive CD4+ T cells (Fig. 8b, right), and TFH cells also showed enrichment for such binding relative to binding in the Tcf7−/− negative control cells (Fig. 8b). Although TCF-1 did not bind to Il6ra in wild-type naive CD4+ T cells, it was recruited to the Il6ra TSS in wild-type TFH cells (Fig. 8b, left), which suggested that recruitment of TCF-1 to this site is part of the TFH differentiation program. Wild-type TH1 cells did not exhibit enrichment for the binding of TCF-1 at Il6st or Il6ra compared with its binding in naive CD4+ T cells (Fig. 8b), in line with the diminished expression of both IL-6Rα and gp130 on TH1 cells (Fig. 7c,e). We did not detect binding of TCF-1 to the TSS of Icos (Supplementary Fig. 7b). These data suggested that TCF-1 directly regulated induction of the expression of IL-6 receptor chains to sustain expression of the IL-6 receptor complex by activated CD4+ T cells in vivo, which allowed TFH differentiation (Supplementary Fig. 8).

ChIP analysis of the binding of TCF-1 to the positive control gene Axin2 (a), the TSS of Il6ra and Il6st (b), the TSS and a regulatory region 2.8 kb upstream of Bcl6 (c), and the TSS of Ascl2 and intron 3 of Prdm1 (d) in naive Tcf7+/+ cells (CD44loCD62L+), naive Tcf7−/− CD4+ T cells (GFP+CD44loCD62L+CD4+), Tcf7+/+ TFH cells (CXCR5+CD44hiCD62L−CD4+) and Tcf7+/+ TH1 cells (CXCR5−CD44hiCD62L−CD4+), with the last two populations sorted from B6 mice 8 d after infection with vaccinia virus; results were normalized to those obtained by ChIP with immunoglobulin G and are presented relative to those obtained for the promoter region of the control gene Hprt. *P < 0.05, **P < 0.01 and ***P < 0.001 (Student's t-test). Data are from three independent experiments (mean and s.d.).
We next investigated by ChIP the association of TCF-1 with genes encoding transcription factors key to TFH differentiation. TCF-1 bound to intron 3 of Prdm1, the major regulatory site of Prdm1 expression35, in both naive CD4+ T cells and CXCR5+ TFH cells (Fig. 8d), which suggested direct involvement of TCF-1 and its homolog LEF-1 in the suppression of Blimp1 in TFH cells. Given that Prdm1 is not expressed by naive CD4+ T cells, binding of TCF-1 at this site suggested that TCF-1 might antagonize Prdm1 expression upon T cell activation. In addition, we observed specific binding of TCF-1 to the TSS of Bcl6 and an upstream regulatory region of Bcl6 in naive CD4+ T cells (Fig. 8c), and this binding pattern was maintained in TFH cells (Fig. 8c). We observed robust enrichment for TCF-1 at Prdm1, Bcl6, Il6ra and Il6st in wild-type TFH cells relative to its abundance at those genes in Tcf7−/− TFH cells (Supplementary Fig. 7b). We did not observe enrichment for TCF-1 binding in the Ascl2 TSS (Fig. 8c,d), although we could not exclude the possibility that Ascl2 is regulated by LEF-1 and TCF-1 through more distal regulatory regions. Binding of TCF-1 to the upstream region of Bcl6 and the Prdm1 intron was abrogated in TH1 cells relative to its binding in TFH cells (Fig. 8d), in line with the substantially reduced expression of TCF-1 in TH1 cells. These observations suggested that downregulation of TCF-1 in TH1 cells was important for upregulation of Blimp1 and Blimp1-mediated repression of Bcl6 in TH1 cells, while retention of TCF-1 in early TFH cells ensured proper upregulation of Bcl6 and subsequent suppression of Blimp1 during TFH differentiation (Supplementary Fig. 8).
Discussion
TFH differentiation can be initiated at an early time point during T cell activation, but the regulators of this important 'decision' process are still being defined. Here we initiated an investigation to identify previously unknown pathways in TFH differentiation by characterizing genes differentially expressed in early TFH cells in vivo relative to their expression in TH1 cells but not modulated by supplementation with IL-6 in vitro. We found that a pair of transcription factors, LEF-1 and TCF-1, influenced TFH differentiation by regulating circuits upstream of Bcl6. We found that LEF-1 and TCF-1 coordinated TFH differentiation by two general mechanisms. First, they established the responsiveness of naive CD4+ T cells to TFH cell signals by promoting the expression of IL-6 receptor chains and binding to Prdm1 and Bcl6. Second, they promoted early TFH differentiation of activated CD4+ T cells via multipronged activities that sustained expression of IL-6Rα and gp130, enhanced ICOS expression and promoted Bcl6 expression while inhibiting Blimp1 expression.
IL-6 is a critical early regulator of TFH differentiation, as Il6−/− mice fail to undergo any differentiation of TFH cells during the dendritic cell–priming phase of an acute antiviral immune response10. In mice whose dendritic cells constitutively overexpress IL-6, the main alteration in phenotype observed is a substantial increase in TFH cells and GCs36. Therefore, regulation of the expression of IL-6 receptors on naive CD4+ T cells and early activated CD4+ T cells is a mechanism by which LEF-1 and TCF-1 influence TFH differentiation.
Bcl6 is essential for TFH differentiation, while Blimp1 is a powerful antagonist of such differentiation. Our observations that expression of LEF-1 resulted in aberrant expression of Bcl6 in TH1 cells, Blimp1 expression was aberrantly upregulated in Lef1−/−Tcf7−/− GC TFH cells, and the genes encoding Bcl6 and Blimp1 were both targets directly bound by TCF-1 indicated that LEF-1 and TCF-1 probably dually regulate both of these critical transcription factors. While we cannot rule out the possibility that the de-repression of Prdm1 resulted from reduced Bcl6 expression in Lef1−/−Tcf7−/− TFH and GC TFH cells, we speculate that LEF-1 and TCF-1 directly repress Prdm1 expression. LEF-1 and TCF-1 are known to positively and negatively regulate gene expression, depending on the interacting factors. For examples, both proteins can interact with the coactivator β-catenin and with transcriptional corepressors of the TLE family, and LEF-1 and TCF-1 repress Cd4 in CD8+ T cells18. Future analysis of molecular mechanisms by which LEF-1 and TCF-1 regulate Prdm1 and Bcl6 will be important, as will analysis of how LEF-1 and TCF-1 interact with other regulators of Bcl6 and Prdm1, such as STAT1, STAT3, STAT5, Foxo1 and Klf2 (refs. 3,10,11,28,37,38). Nevertheless, our data have provided proof that LEF-1 and TCF-1 regulate the balance between Bcl6 expression and Blimp1 expression.
ICOS expression was selectively impaired on Lef1−/−Tcf7−/− TFH cells, and ICOS expression was enhanced on Lef1-RV+ cells. In multiple models, moderate changes in ICOS have been observed to enhance the differentiation of TFH cells38,39,40,41 or their function42. ICOS seems to be not a direct target of LEF-1 and TCF-1, although distal cis elements have not been explored. Alternatively, ICOS might be indirectly regulated by LEF-1 and TCF-1. Future studies should further elucidate the LEF-1 and TCF-1 signaling axes that modulate ICOS expression. Overall, the combined influence of LEF-1 and TCF-1 on IL-6Rα, gp130, Bcl6, Blimp1 and ICOS produces a dense network of interactions that create a strong pro-TFH cell signaling environment in a cell that sustains the expression of LEF-1 and/or TCF-1.
The functions of LEF-1 and TCF-1 probably continue to be important in fully differentiated TFH cells and GC TFH cells. LEF-1 and TCF-1 both continue to be expressed in GC TFH cells. Bcl6 expression is essential in GC TFH cells3, and continued regulation of both Bcl6 and Prdm1 are central aspects of GC TFH cell biology. ICOS is also a major regulator of GC TFH cell biology26,40. Signaling via the IL-6 receptor is not usually essential in GC TFH cells due to compensatory abilities of IL-21 or IL-27 at later time points29,43,44. Nevertheless, the IL-6 receptor probably has a major role in sustaining GC TFH cells under normal physiological conditions. IL-6 is required for sustaining TFH cell and GC responses during chronic infection with LCMV in mice45, and IL-6 is positively associated with TFH cells and GCs in macaques positive for simian immunodeficiency virus46.
The activities of LEF-1 and TCF-1 seem to pre-program the responsiveness of a given naive CD4+ T cell to TFH cell signals, prior to any exposure of the cell to antigen. Therefore, we speculate that transient or sustained inflammatory or pathogenic conditions that alter the expression of LEF-1 or TCF-1 in naive T cells might have a global effect that alters the capacity of naive CD4+ T cells to respond to TFH cell–induction signals in the presence of pathogens or autoimmunity triggers. Ultimately, it will be useful to determine how homeostatic signals act in concert with LEF-1 and TCF-1 to modulate the expression or poised status of TFH cell–associated genes in naive CD4+ T cells to properly orchestrate the development progression from naive cell to the TFH cell or non-TFH cell fate.
LEF-1 and TCF-1 have high expression in resting naive CD4+ and CD8+ T cells, but the expression of LEF-1 and TCF-1 is downregulated in effector CD8+ T cells and TH1 cells, which suggests Lef1 and Tcf7 are regulated by T cell activation. Dwell time at the T cell antigen receptor influences TFH differentiation versus non-TFH differentiation in a manner intrinsic to the signal strength of the receptor47. We speculate these processes may be interrelated.
In conclusion, our study has identified previously unknown roles for LEF-1 and TCF-1 in TFH differentiation. Better understanding of the downstream targets of LEF-1 and TCF-1 in activated CD4+ T cells will improve the understanding of TFH cell biology. Finally, better understanding of the signals that regulate LEF-1 and TCF-1 will have implications for understanding how to enhance or inhibit the differentiation of TFH cells.
Methods
Mice and viral infection.
C57BL/6J (B6), B6.SJL, Cd4-Cre, and Rosa26GFP mice were from the Jackson Laboratory. Mouse strains described below were from in-house breeders of either the La Jolla Institute or the University of Iowa animal facility. SMARTA mice (specific for LCMV glycoprotein amino acids 66–77 presented by I-Ab)48 and Tcf7fl/fl and Lef1fl/fl mice16,18 have been described. Bcl6fl/fl mice were from T. Takemori49 and hCD2-Cre mice were from P.E.L.32. Blimp1-YFP mice (expressing a bacterial artificial chromosome transgene) were crossed to the SMARTA strain to generate Blimp1-YFP SMARTA mice26. Tcf7-GFP reporter mice were generated in-house (unpublished data). All mice analyzed were 6–12 weeks of age, and both sexes were included without randomization or 'blinding' of researchers to mouse or sample identity. All mouse experiments were performed under protocols approved by the Institutional Animal Use and Care Committees of the La Jolla Institute and the University of Iowa. For acute viral infection, 2.5 × 105 to 5.0 × 105 plaque-forming units of LCMV (Armstrong strain) and 2.5 × 105 plaque-forming units of vaccinia virus were used. Virus was prepared in plain DMEM and was injected intraperitoneally or intravenously.
Flow cytometry.
Single-cell suspensions were prepared from the spleen of mice infected with LCMV or vaccinia virus, and surfaces were stained as described16,26. The fluorochrome-conjugated antibodies were as follows: anti-CD4 (RM4-5), anti-CD44 (IM7), anti-CD62L (MEL-14), anti-PD-1 (J43), anti-IL-6Rα (D7715A7), anti-gp130 (KGP130), anti-ICOS (C398.4A), anti-Fas (15A7), anti-GL7 (GL7), anti-IgD (11-26), anti-CD138 (281-2) and anti-Bcl6 (K112-91) (all from eBiosciences); anti-SLAM (TC15-12F12.2; BioLegend); and anti-PSGL-1 (2PH1; BD Biosciences). For detection of CXCR5, a two-step26 or three-step6 staining protocol was used with biotinylated anti-CXCR5 or unconjugated anti-CXCR5, respectively (2G8; BD Biosciences). For intracellular detection of Bcl6, surface-stained cells were fixed and permeabilized with the Foxp3/Transcription Factor Staining Buffer Set (eBiosciences), followed by incubation with fluorochrome-conjugated anti-Bcl6. Data were collected on an LSRII and a FACSVerse (BD Biosciences) and were analyzed with FlowJo software (TreeStar).
Immunoblot analysis.
For analysis of the knockdown of LEF-1 or targeted deletion of TCF-1 and LEF-1, shCtrl+ and shLef1+ SMARTA cells or CD4+ and CD8+ T cells (5 × 105 each) were sorted, followed by denaturation for 5 min at 100 °C in SDS loading buffer. Cell lysates were probed with anti-TCF-1 (C46C7; Cell Signaling Technology), anti-LEF-1 (C18A7 and C12A5; Cell Signaling Technology) or anti-β-actin (loading control; I-19; Santa Cruz Biotechnology).
Retroviral transduction.
Naive SMARTA CD4+ T cells were purified by negative selection with either magnetic beads (Miltenyi Biotec) or an EasySep kit (StemCell), and were resuspended in D-10 medium (DMEM containing 10% FCS, 2 mM GlutaMax (Life Technologies), 100 U/ml penicillin and streptomycin (Life Technologies) and 50 μM β-mercaptoethanol) with 2 ng/ml human IL-7 or 10 ng/ml human IL-2 (Peprotech). 2 × 106 SMARTA cells were seeded in 24-well plates coated with 8 μg/ml anti-CD3 (17A2; BioXcell) and anti-CD28 (37.51; BioXcell). Retroviral supernatants were added at 24 and 36 h after stimulation. After 72 h of in vitro stimulation, SMARTA cells were transferred into six-well plates in D-10 medium with 10 ng/ml human IL-2, followed by incubation for 2 d. One day before reporter-expressing cells were sorted (with a FACSAria from BD Biosciences) for transfer, the culture medium was replaced with D-10 medium with 2 ng/ml human IL-7. Detailed information has been published50.
Cell sorting.
All cell sorting was done on a FACSAria (BD Biosciences). For RNA-seq analysis, early TFH cells (IL-2Rα−Blimp1-YFP−) or early TH1 cells (IL-2Rα+Blimp1-YFP+) among SMARTA cells, or the CXCR5− subset (TH1), PD-1loCXCR5+ subset (TFH), and PD-1hiCXCR5+ subset (GC TFH) of activated GFP+CD4+ splenic T cells of Lef1−/−Tcf7−/− mice or their control littermates were sorted on day 3 after infection with LCMV or on day 8 after infection with vaccinia virus, respectively. GFP-RV+ or Lef1-RV+ SMARTA cells were sorted as SLAMhiCXCR5lo (TH1) or SLAMloCXCR5hi (TFH) cells on day 4 after LCMV infection. For ChIP analysis, CXCR5− (TH1) and CXCR5+ (TFH) cells were sorted from activated CD4+ splenic T cells on day 8 after infection with vaccinia virus. Also, CD44loCD62Lhi naive CD4+ T cells were sorted from wild-type or Tcf7−/− (Tcf7fl/flCd4-Cre) mice.
Retrovirus production and cell transfer.
Mouse Lef1 cDNA (6401514; Open Biosystems) was cloned into a retroviral expression vector (pMIG-GFP). The Lef1-specific shRNA sequence (Transomic) was cloned into pLMPd-Ametrine vector, as reported26,31. The vector pLMPd-Ametrine with shRNA sequence (5′-TGCTGTTGACAGTGAGCGAATGGATAAGTCTGACGACCTATAGTGAAGCCACAGATGTAT AGGTCGTCAGACTTATCCATGTGCCTACTGCCTCGGA-3′) directed against mouse Cd19 served as a negative control (shCtrl) in knockdown experiments. Virions were obtained from Plat-E cells as described50. Culture supernatants were collected 24 and 48 h after transfection, then were filtered through a 0.45-μm syringe filter and saved at 4 °C until used for transduction.
Naive or retrovirus-transduced SMARTA cells were transferred intravenously into mice via the retro-orbital sinus. For transduced SMARTA cells, 100% of the transferred cells were transduced (Ametrine+CD45.1+). The number of cells transferredwas 4 × 105 to 5 × 105, 2 × 105, or 5 × 103 SMARTA cells on day 3, 4 or 8, respectively.
In vitro activation of CD4+ T cells.
Naive SMARTA cells were negatively isolated through the use of a CD4+ T cell isolation kit (Miltenyi or StemCell). 2 × 106 SMARTA cells were seeded on 24-well plates coated with 8 μg/ml anti-CD3 (17A2; BioXcell) and anti-CD28 (37.51; BioXcell). For TH1 polarization, SMARTA cells were treated with 20 μg/ml of anti-IL-4 (11B11; BioXcell) and antibody to transforming growth factor-β (1D11; BioXcell) and 20 ng/ml of recombinant mouse IL-12 (Peprotech). For IL-6 condition, 10 μg/ml of antibody to interferon IFN-γ (XMG1.2; BioXcell) and anti-IL-12 (R1-5D9; BioXcell) and 20 ng/ml of recombinant mouse IL-6 (Peprotech) were added to the culture medium.
Quantitative RT-PCR.
Total RNA from the sorted cells was extracted and reverse-transcribed, and quantitative PCR was performed as described16.
RNA-seq and transcriptome analysis (protocol used by the Xue laboratory).
Total RNA was extracted from PD-1+CXCR5+ cells sorted from Tcf7−/−Lef1−/− mice or their control littermates, and two samples were obtained for each genotype. cDNA synthesis and amplification were performed with a SMARTer Ultra Low Input RNA Kit, starting with 10 ng of total RNA per sample, according to the manufacturer's instructions (Clontech). cDNA was fragmented with a Q800R sonicator (Qsonica) and was used as input for a NEBNext Ultra DNA Library Preparation Kit (NEB). Libraries were sequenced on a HiSeq2000 (Illumina) in single-read mode, with a read length of 50 nucleotides producing 60 × 106 to 70 × 106 reads per sample. Sequence data in 'fastq' format were generated with the CASAVA 1.8.2 processing pipeline from Illumina.
The sequencing quality of RNA-seq libraries was assessed by the FastQC quality control tool for high-throughput sequence data (version 0.10.1; Bioinformatics Group of the Babraham Institute). Because of biased GC content in the 5′ end, the first 12 bases of each read in all four samples were 'trimmed off'. The reproducibility of RNA-seq data was evaluated by computation of Pearson's correlation of FPKM (fragments per kilobase of exon per million fragments mapped) values for all genes in biological replicates. The Pearson's correlation coefficient between the two biological replicates was 0.937 for the control samples and 0.986 for the Tcf7−/− Lef1−/− samples, indicative of good reproducibility.
The RNA-seq libraries were then processed by the RSEM package ('RNA-seq by Expectation-Maximization'; version 1.2.19) for estimation of the expression level of each gene. The expression level of a gene is reported as a 'gene-level' FPKM value. EBSeq (version 1.5.4), an integral component of the RSEM package, was used for the identification of differentially expressed genes. Genes of the mm9 (UCSC) assembly of the mouse genome from the iGenome collection of reference sequences were used for gene annotation.
RNA-seq and transcriptome analysis (protocol used by the Crotty laboratory).
Cells were stored in Trizol, and total RNA was extracted from the cells with an miRNeasy Mini Kit (Qiagen 217004). For RNA-seq analysis of early TFH cells and TH1 cells: poly(A) RNA was isolated from 200 ng total RNA of each sample through the use of a Poly(A) Purist MAG kit (AM1922; Ambion). The resulting poly(A) RNA was then fragmented and prepared according to the manufacturer's instructions (ABI 4452437 Rev B), into 'bar-coded', strand-specific libraries with The SOLiD Total RNA-seq Kit (ABI 4445374). Following library preparation, 15 ng of each library was converted into SOLiD Wildfire compatible fragments with a 5500 W Conversion Primer Kit (Life Technologies) and five rounds of PCR. Libraries were then pooled at equimolar concentrations with a Quant-iT PicoGreen dsDNA Assay Kit (Life Technologies) and were sequenced on a 5500XL W Genetic Analyzer (Life Technologies). SOLiD 5500-2 sequencing outcomes were converted from 'color space' to 'nucleotide space' through the use of solid2fastq script (Galaxy). For RNA-seq analysis of GFP-RV+ or Lef1-RV+ SMARTA cells obtained 4 d after infection with LCMV, 500 ng of each sample's total RNA was prepared into mRNA libraries according to manufacturer's instructions (RS-122-2103; Illumina). The resulting libraries were deep sequenced on an Illumina 2500 in Rapid Run Mode, through the use of single-end reads with a length of 50 nucleotides (>24 × 106 reads per condition). The single-end reads that passed Illumina filters were filtered for reads aligning to tRNA, rRNA, adaptor sequences, and 'spike-in' controls.
The reads were then aligned to the UCSC mm9 reference genome through the use of TopHat software (version 1.4.1). 'DUST scores' (for filtering low-complexity regions) were calculated with PRINSEQ Lite data preprocessing software (version 0.20.3), and low-complexity reads (with a 'DUST score' of >4) were removed from the BAM files (binary alignment map). The alignment results were parsed via SAMtools to generate SAM files (sequence alignment map). Read counts to each genomic feature were obtained with the htseq-count program (version 0.6.0) with the 'union' option. After removal of absent features (zero counts in all samples), the raw counts were then imported to software of the R project for statistical computing (R/Bioconductor package DESeq2) for the identification of genes differentially expressed among samples. DESeq2 normalizes counts by dividing each column of the 'count table' (samples) by the size factor of the column. The size factor is calculated by division of the samples by geometric means of the sequence reads of the genes. This brings the count values to a common scale suitable for comparison. P values for differential expression were calculated with the binomial test for differences between the base means of two conditions. These P values were then adjusted for multiple-test correction with the Benjamini-Hochberg algorithm to control the false-discovery rate. We considered genes as being expressed differentially between two groups of samples when the DESeq2 analysis resulted in an adjusted P value of <0.05 and the difference in gene expression was 1.5-fold. Cluster analyses, including principal-component analysis and hierarchical clustering, were performed with standard algorithms and metrics. Hierarchical clustering was performed with complete linkage with Euclidean metric.
Heat maps.
Heat maps were generated with normalized data of RNA-seq analyses for early TFH cells and TH1 cells and for GFP-RV+ and Lef1-RV+ TFH cells and TH1 cells. Microarray analysis used published TH1 cell sets, TFH cell sets and GC TFH cell sets (GEO accession code GSE21380)51 and the GenePattern software suite (Broad Institute).
GSEA.
GSEA was performed with GSEA software from the Broad Institute. Gene sets were generated in-house with genes that had a difference in expression of more than twofold in TFH cells (PD-1loCXCR5+) and GC TFH cells (PD-1hiCXCR5+) relative to their expression in TH1 cells (PD-1−CXCR5−) (GEO accession code GSE21380). Enrichment for genes that were upregulated more than 1.2-fold in Lef1-RV+ TH1 cells relative to their expression in GFP-RV+ TH1 cells was then ranked by the 'Diff_of_Classes' metric of GSEA software.
ChIP.
Sorted CD4+ T cells were cross-linked for 5 min with 1% formaldehyde in medium, were processed with a truChIP Chromatin Shearing Reagent Kit (Covaris) and were sonicated for 5 min on Covaris S2 ultrasonicator. The sheared chromatin was immunoprecipitated with anti-TCF-1 (C46C7; Cell Signaling Technologies) or control rabbit immunoglobulin G (2729; Cell Signaling Technologies) and was washed as described18. The immunoprecipitated DNA segments were used for quantification by PCR. For calculation of enrichment in the binding of TCF-1 in a given cell type, each ChIP sample analyzed with TCF-1 was first normalized to corresponding ChIP sample analyzed with immunoglobulin G, and the signal at a target region was then normalized to that at the Hprt promoter region.
Statistical analysis.
Data sets were analyzed with the Student's t-test with a two-tailed distribution assuming equal sample variance.
Accession codes.
References
Crotty, S. A brief history of T cell help to B cells. Nat. Rev. Immunol. 15, 185–189 (2015).
Ueno, H., Banchereau, J. & Vinuesa, C.G. Pathophysiology of T follicular helper cells in humans and mice. Nat. Immunol. 16, 142–152 (2015).
Crotty, S. T follicular helper cell differentiation, function, and roles in disease. Immunity 41, 529–542 (2014).
Shlomchik, M.J. & Weisel, F. Germinal center selection and the development of memory B and plasma cells. Immunol. Rev. 247, 52–63 (2012).
Victora, G.D. & Nussenzweig, M.C. Germinal centers. Annu. Rev. Immunol. 30, 429–457 (2012).
Johnston, R.J. et al. Bcl6 and Blimp-1 are reciprocal and antagonistic regulators of T follicular helper cell differentiation. Science 325, 1006–1010 (2009).
Nurieva, R.I. et al. Bcl6 mediates the development of T follicular helper cells. Science 325, 1001–1005 (2009).
Yu, D. et al. The transcriptional repressor Bcl-6 directs T follicular helper cell lineage commitment. Immunity 31, 457–468 (2009).
Ise, W. et al. The transcription factor BATF controls the global regulators of class-switch recombination in both B cells and T cells. Nat. Immunol. 12, 536–543 (2011).
Choi, Y.S., Eto, D., Yang, J.A., Lao, C. & Crotty, S. Cutting edge: STAT1 is required for IL-6-mediated Bcl6 induction for early follicular helper cell differentiation. J. Immunol. 190, 3049–3053 (2013).
Nurieva, R.I. et al. Generation of T follicular helper cells is mediated by interleukin-21 but independent of T helper 1, 2, or 17 cell lineages. Immunity 29, 138–149 (2008).
Ray, J.P. et al. Transcription factor STAT3 and type I interferons are corepressive insulators for differentiation of follicular helper and T helper 1 cells. Immunity 40, 367–377 (2014).
Liu, X. et al. Transcription factor achaete-scute homologue 2 initiates follicular T-helper-cell development. Nature 507, 513–518 (2014).
Choi, Y.S., Yang, J.A. & Crotty, S. Dynamic regulation of Bcl6 in follicular helper CD4 T (Tfh) cells. Curr. Opin. Immunol. 25, 366–372 (2013).
Weber, B.N. et al. A critical role for TCF-1 in T-lineage specification and differentiation. Nature 476, 63–68 (2011).
Yu, S. et al. The TCF-1 and LEF-1 transcription factors have cooperative and opposing roles in T cell development and malignancy. Immunity 37, 813–826 (2012).
Steinke, F.C. & Xue, H.-H. From inception to output, Tcf1 and Lef1 safeguard development of T cells and innate immune cells. Immunol. Res. 59, 45–55 (2014).
Steinke, F.C. et al. TCF-1 and LEF-1 act upstream of Th-POK to promote the CD4+ T cell fate and interact with Runx3 to silence Cd4 in CD8+ T cells. Nat. Immunol. 15, 646–656 (2014).
Jeannet, G. et al. Essential role of the Wnt pathway effector Tcf-1 for the establishment of functional CD8 T cell memory. Proc. Natl. Acad. Sci. USA 107, 9777–9782 (2010).
Zhou, X. & Xue, H.-H. Cutting edge: generation of memory precursors and functional memory CD8+ T cells depends on T cell factor-1 and lymphoid enhancer-binding factor-1. J. Immunol. 189, 2722–2726 (2012).
Zhou, X. et al. Differentiation and persistence of memory CD8+ T cells depend on T cell factor 1. Immunity 33, 229–240 (2010).
Yu, Q. et al. T cell factor 1 initiates the T helper type 2 fate by inducing the transcription factor GATA-3 and repressing interferon-γ. Nat. Immunol. 10, 992–999 (2009).
Yu, Q., Sharma, A., Ghosh, A. & Sen, J.M. T cell factor-1 negatively regulates expression of IL-17 family of cytokines and protects mice from experimental autoimmune encephalomyelitis. J. Immunol. 186, 3946–3952 (2011).
van Loosdregt, J. et al. Canonical Wnt signaling negatively modulates regulatory T cell function. Immunity 39, 298–310 (2013).
Choi, Y.S. et al. Bcl6 expressing follicular helper CD4 T cells are fate committed early and have the capacity to form memory. J. Immunol. 190, 4014–4026 (2013).
Choi, Y.S. et al. ICOS receptor instructs T follicular helper cell versus effector cell differentiation via induction of the transcriptional repressor Bcl6. Immunity 34, 932–946 (2011).
Pepper, M., Pagán, A.J., Igyártó, B.Z., Taylor, J.J. & Jenkins, M.K. Opposing signals from the bcl6 transcription factor and the interleukin-2 receptor generate T helper 1 central and effector memory cells. Immunity 35, 583–595 (2011).
Johnston, R.J., Choi, Y.S., Diamond, J.A., Yang, J.A. & Crotty, S. STAT5 is a potent negative regulator of TFH cell differentiation. J. Exp. Med. 209, 243–250 (2012).
Eto, D. et al. IL-21 and IL-6 are critical for different aspects of B cell immunity and redundantly induce optimal follicular helper CD4 T cell (Tfh) differentiation. PLoS ONE 6, e17739 (2011).
Oestreich, K.J., Mohn, S.E. & Weinmann, A.S. Molecular mechanisms that control the expression and activity of Bcl-6 in TH1 cells to regulate flexibility with a TFH-like gene profile. Nat. Immunol. 13, 405–411 (2012).
Chen, R. et al. In vivo RNA interference screens identify regulators of antiviral CD4+ and CD8+ T cell differentiation. Immunity 41, 325–338 (2014).
Vacchio, M.S. et al. A ThPOK-LRF transcriptional node maintains the integrity and effector potential of post-thymic CD4+ T cells. Nat. Immunol. 15, 947–956 (2014).
Zhao, D.-M. et al. Constitutive activation of Wnt signaling favors generation of memory CD8 T cells. J. Immunol. 184, 1191–1199 (2010).
Nance, J.P., Belanger, S., Johnston, R.J., Takemori, T. & Crotty, S. Cutting edge: T follicular helper cell differentiation is defective in the absence of Bcl6 BTB repressor domain function. J. Immunol. 194, 5599–5603 (2015).
Tunyaplin, C. et al. Direct repression of prdm1 by Bcl-6 inhibits plasmacytic differentiation. J. Immunol. 173, 1158–1165 (2004).
Kim, S.J., Zou, Y.R., Goldstein, J., Reizis, B. & Diamond, B. Tolerogenic function of Blimp-1 in dendritic cells. J. Exp. Med. 208, 2193–2199 (2011).
Lee, J.-Y. et al. The transcription factor KLF2 restrains CD4+ T follicular helper cell differentiation. Immunity 42, 252–264 (2015).
Stone, E.L. et al. ICOS coreceptor signaling inactivates the transcription factor FOXO1 to promote Tfh cell differentiation. Immunity 42, 239–251 (2015).
Pratama, A. et al. MicroRNA-146a regulates ICOS-ICOSL signalling to limit accumulation of T follicular helper cells and germinal centres. Nat. Commun. 6, 6436 (2015).
Weber, J.P. et al. ICOS maintains the T follicular helper cell phenotype by down-regulating Krüppel-like factor 2. J. Exp. Med. 212, 217–233 (2015).
Vogel, K.U. et al. Roquin paralogs 1 and 2 redundantly repress the Icos and Ox40 costimulator mRNAs and control follicular helper T cell differentiation. Immunity 38, 655–668 (2013).
Xu, H. et al. Follicular T-helper cell recruitment governed by bystander B cells and ICOS-driven motility. Nature 496, 523–527 (2013).
Batten, M. et al. IL-27 supports germinal center function by enhancing IL-21 production and the function of T follicular helper cells. J. Exp. Med. 207, 2895–2906 (2010).
Poholek, A.C. et al. In vivo regulation of Bcl6 and T follicular helper cell development. J. Immunol. 185, 313–326 (2010).
Harker, J.A., Lewis, G.M., Mack, L. & Zuniga, E.I. Late interleukin-6 escalates T follicular helper cell responses and controls a chronic viral infection. Science 334, 825–829 (2011).
Petrovas, C. et al. CD4 T follicular helper cell dynamics during SIV infection. J. Clin. Invest. 122, 3281–3294 (2012).
Tubo, N.J. et al. Single naive CD4+ T cells from a diverse repertoire produce different effector cell types during infection. Cell 153, 785–796 (2013).
Oxenius, A., Bachmann, M.F., Zinkernagel, R.M. & Hengartner, H. Virus-specific MHC-class II-restricted TCR-transgenic mice: effects on humoral and cellular immune responses after viral infection. Eur. J. Immunol. 28, 390–400 (1998).
Kaji, T. et al. Distinct cellular pathways select germline-encoded and somatically mutated antibodies into immunological memory. J. Exp. Med. 209, 2079–2097 (2012).
Choi, Y.S. & Crotty, S. Retroviral vector expression in TCR transgenic CD4+ T cells. Methods Mol. Biol. 1291, 49–61 (2015).
Yusuf, I. et al. Germinal center T follicular helper cell IL-4 production is dependent on signaling lymphocytic activation molecule receptor (CD150). J. Immunol. 185, 190–202 (2010).
Acknowledgements
We thank J. Yang for help with ex vivo screening of target genes obtained from RNA-seq analysis of early TFH cells and TH1 cells; Z. Fu for help with GSEA and for the generation of heat maps; the La Jolla Institute Flow Cytometry Flow Cytometry Core Facility (C. Kim, K.V. Gunst and L. Nosworthy) and the University of Iowa Flow Cytometry Core facility (J. Fishbaugh, H. Vignes and G. Rasmussen) for cell sorting; T. Takemori (RIKEN, Research Center for Integrated Medical Sciences) for Bcl6fl/fl mice; I. Antoshechkin (Millard and Muriel Jacobs Genetics and Genomics Laboratory at the Caltech) for Tcf7−/−Lef1−/− RNA-seq; and J.T. Harty (University of Iowa) for vaccinia virus (for the Xue laboratory). Supported by the American Cancer Society (RSG-11-161-01-MPC to H.-H.X.), the US National Institutes of Health (AI105351, AI112579, AI115149 and AI119160 to H.-H.X.; AI113806 to W.P.; AI109976, AI063107 and AI072543 to S.C.; and AI007485 for support for to J.A.G.) and the University of Iowa Presidential Graduate Research Fellowship program (J.A.G.).
Author information
Authors and Affiliations
Contributions
Y.S.C., J.A.G., S.X., Q.S. and F.L. performed the experiments and analyzed the data; Z.Z. analyzed the RNA-seq data under the supervision of W.P.; P.E.L. provided reagents; Y.S.C., H.-H.X. and S.C. conceived of the project and wrote the paper; and H.-H.X. and S.C. supervised the overall study.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 1 Downregulation of the expression of Lef1 and Tcf7 in activated CD4+ T cells.
(a-e) RNA-sequencing analysis was performed with naïve SMARTA cells or SMARTA cells that were in vitro stimulated under TH1 polarizing condition or IL-6 condition for three days (described in Methods section). Shown are normalized reads for Ifng (a), Tbx21 (b), Il21 (c), Lef1 (d), and Tcf7 (e) transcripts. (f) SMARTA cells that were stimulated in vitro under IL-6 condition were analyzed for surface expression of CXCR5 by FACS. Shown are overlaid histograms of SA-PE signals obtained by staining CD4+ T cells with αIgD-Biotin Abs (filled gray histogram) and αCXCR5-Biotin (red histogram) Abs. (g) Fold changes in gene expression obtained from RNA-sequencing analyses for early TFH vs. early TH1 and for in vitro stimulated CD4+ T cells (as described in a-e) vs. CD44loCD62Lhi naïve CD4+ T cells.
Supplementary Figure 2 T cell development in Lef1fl/flhCD2-Cre, Tcf7fl/flhCD2-Cre and Lef1fl/flTcf7fl/flhCD2-Cre mice.
(a-c) Ablation of TCF-1 and LEF-1 proteins in mature CD4+ T cells using the hCD2-Cre. a. Excision efficiency of floxed target sequence by hCD2-Cre as marked by GFP expression. hCD2-Cre transgene was crossed to the Rosa26-STOP-GFP mice, and thymic and splenic CD4+ T cells were analyzed for GFP expression. The percentage of GFP+ cells is shown. b and c. Effective ablation of LEF-1 (b) and TCF-1 (c) proteins in GFP+CD4+ T cells. GFP+CD4+ and GFP−CD4+ T cells were sorted from the splenocytes of hCD2-cre:Rosa26GFP:Tcf7fl/fl:Lef1fl/fl (Tcf7−/−Lef1−/−) mice. WT CD4+ splenic T cells were sorted as control. Cell lysates from the sorted cells were immunoblotted to detect LEF-1 or TCF-1 proteins. β-actin, equal loading control. Data are representative from ≥ 2 experiments. (d-i) hCD2-Cre-mediated ablation of LEF-1 and/or TCF-1 did not detectably disturb thymic development and naïve T cell compartments in the periphery. Thymocytes and splenocytes were collected from un-infected Lef1−/−, Tcf7−/−, Tcf7−/−Lef1−/− mice and littermate controls. The cells were surface-stained and analyzed for T cell subset distribution. d. Thymocyte development. The percentages of thymocytes in each developmental stage are shown as means ± s.d. from ≥ 3 experiments. e. Numbers of total, CD4+, and CD8+ thymocytes. (n = 7 for control, 3 for Lef1−/−, 6 for Tcf7−/−, and 6 for Tcf7−/−Lef1−/− mice.) f. T cell output in the periphery. The percentages of CD4+ and CD8+ T cells in the spleen are shown as means ± s.d. from ≥ 3 experiments. g. Numbers of total, CD4+, and CD8+ splenocytes. (n = 5 for control, 3 for Lef1−/−, 5 for Tcf7−/−, and 4 for Tcf7−/−Lef1−/− mice.) h and i Detection of naïve and antigen-experienced subsets in CD4+ and CD8+ T cells. CD44loCD62L+ naïve, CD44hiCD62L+ memory phenotype, and CD44hiCD62L− effector phenotype subsets were identified within CD4+ (h) and CD8+ (i) T cells. The percentages are shown as means ± s.d. from ≥ 3 experiments. (n = 4 for control, 3 for Lef1−/−, 4 for Tcf7−/−, and 3 for Tcf7−/−Lef1−/− mice.) In the bar graphs of (h) and (i), also marked are the fold changes in the cell numbers in gene-targeted animals over control mice. None of these changes reached statistical significance by student’s t-test.
Supplementary Figure 3 Lef1 deficiency in CD4+ T cells did not affect generation of TH1 cells after infection with vaccinia virus.
Lef1−/− mice and littermate controls were infected with vaccinia virus, and 8 days later, splenocytes were harvested and surface-stained. The activated GFP+CD4+ T cells were analyzed for SLAMhiCXCR5− TH1 T cells. The frequency of TH1 within activated CD4+ T cells calculated. Data are means ± s.d. (n = 4-5 from 4 independent experiments). The frequency of TH1 in Lef1−/− mice cells was comparable to that of littermates. While TH1 cell number was reduced about 40% in Lef1−/− mice in comparison to that of littermates, it did not reach statistical significance by Student’s t-test.
Supplementary Figure 4 Tcf7-GFP reporter mice.
(a-d) Generation and characterization of the Tcf7 -GFP reporter allele. a. Targeting strategy. TCF-1 is expressed in multiple isoforms in T cells due to differential promoter usage (from exon 1 or exon 3) and alternative splicing. The GFP expression cassette was inserted into intron 1 to track the expression of TCF-1 long isoforms. The GFP expression cassette contains a strong En2 gene splicing acceptor (En2-SA) to facilitate splicing of Tcf7 exon 1 to the reporter, an internal ribosome entry site (IRES) to facilitate independent translation of enhanced GFP, and the GFP cDNA followed by poly-adenylation sequence. b-c. The Tcf7-GFP reporter activity in T cell subsets. Splenocytes were harvested from Tcf7GFP/+ mice. GFP expression was detected in CD44loCD62L+ naïve, CD44hiCD62L+ memory phenotype, and CD44hiCD62L− effector phenotype subsets within CD4+ (b) and CD8+ (c) T cell compartments. d. The GFP reporter was also detected in CD4+CD25+ regulatory T cells but was absent in B220+CD19+ B cells. The percentage of GFP+ cells in each population is shown. (e-f) TCF-1/GFP expression is strongly retained in TFH differentiation program. e. Tcf7GFP/+ mice were infected with vaccinia virus, and eight days later splenocytes were harvested and surface-stained. TH1 and TFH cells were identified in CD44+CD62L− activated CD4+ T cells, and GFP expression was further analyzed. f. Tcf7GFP/+ SMARTA CD4+ T cells were adoptively transferred into CD45.1+ congenic mice followed by i.v. infection with Listeria monocytogenes expressing the GP61 epitope of LCMV. Four days later splenocytes were harvested and surface stained. TH1 and TFH cells were identified in CD45.2+ SMARTA T cells, and GFP expression was further analyzed. Values in histograms are percentages of GFP+ cells in each subset.
Supplementary Figure 5 Impaired vaccinia virus-specific immunoglobulin production in Lef1−/−Tcf7−/− mice.
Lef1−/−Tcf7−/− mice and littermate control mice were infected with vaccinia virus. Eight days later, serum was collected to measure vaccinia virus-specific Ig. Shown is amount of vaccinia virus-specific Ig produced by Lef1−/−Tcf7−/− or littermate control mice, calculated as relative units per ml (RU/mL) to a positive control mAb (LA5, mouse monoclonal anti-D8 mAb, as described in Matho et al., J. Virol. 86, 8050-8058, 2012)
Supplementary Figure 6 Ectopic Lef1 expression does not ‘rescue’ the TFH-differentiation defect of Bcl6-deficient SMARTA T cells.
GFP-RV+ or Lef1-RV+ Cd4-Cre Bcl6fl/fl SMARTA cells were transferred into CD45.2+ B6 mice, which were then infected with LCMV-Arm. Eight days later, RV+ SMARTA cells (a) and endogenous activated (CD44hiCD62Llo) CD4+ T cells (b) were analyzed for SLAMloCXCR5+ TFH differentiation. Gates indicate SLAMloCXCR5+ TFH cells. c. Frequency of SLAMloCXCR5+ TFH cells was calculated as % of endogenous CD44+CD62Llo CD4+ T cells or RV+ SMARTA cells.
Supplementary Figure 7 TCF-1 directly binds to key TFH cell genes.
a. Selection of TCF-1 binding sites at target gene loci. TCF-1 ChIP-seq was performed on naïve CD4+ T cells, and TCF-1 binding locations were summarized for select key TFH genes. TSS, transcription start site. Conservation (4th column) refers to conservation of the motif when mouse sequence was compared with 9 other species including rat, human, chimp, rhesus, marmoset, cow, horse, dog, and opossum. *, for the two motifs within the Bcl6 TSS, one is conserved among all but marmoset and opossum, and the other is conserved among all but dog and opossum. The primer sequences for amplification of each site are:
Il6ra TSS: 5’-AGGAGCACGCAGTTATAGGC and 5’-GTTCCGTTCGCAGAGTGAG;
Il6st TSS: 5’-GTGTCCCGTGAAATGCTACC and 5’-GTACGGACCGAAGGGAAAGT;
Bcl6 TSS: 5’-GGCTGATCTTCGGCATTTAT and 5’-TGGCTCCAAGTTTCCTATGTG;
Bcl6 upstream: 5’-CAGAGCAAAGGGAAATGTGC and 5’-AATAAAGCAGGGCTGGTGAG;
Prdm1 intron3: 5’-GTGTGGCTTCATCAAAGCAG and 5’-GAAGTTTGCGCGTCAGAGTA;
Ascl2 TSS: 5’-GCCTACTGACGTGCTTCCAT and 5’-TACATTCGGACCCTCTCTCG.
qPCR primers were:
Prdm1: 5’-TGGTATTGTCGGGACTTTGC and 5’- ATTGCTTTGGGTTGCTTTCC
Ascl2: 5’- GAGAGCTAAGCCCGATGGAG and 5’- CCCAGGTTTCTTGGGCTAGA.
b. Specific TCF-1 binding to key TFH gene loci in TFH cells. WT or Tcf7−/− mice were infected with vaccinia virus. Eight days later, CXCR5+ TFH cells were sorted from CD44+CD62L− activated CD4+ cells and used for ChIP with anti-TCF-1 antibody or rabbit IgG. Enriched TCF-1 binding was detected at the indicated gene loci in both WT and TCF-1-deficient TFH cells, and shown is ratio of enrichment in WT to Tcf7−/− TFH cells. Data are from 2 independent experiments with duplicate measurements. ns, not statistically significant; *, p<0.05; **, p<0.01; ***, p<0.001 compared with the Hprt locus.
Supplementary Figure 8 Proposed model of the functions of LEF-1 and TCF-1 in the TFH differentiation of CD4+ T cells.
Both in the steady-state and during early TFH differentiation, LEF-1/TCF-1 promote CD4+ T cells to develop into TFH cells by binding to Il6ra and Il6st to activate and/or to Prdm1 to repress. Together with signals downstream of ICOS and IL-6 receptors, LEF-1/TCF-1 also bind to and activate Bcl6 to support maintenance and polarization of TFH differentiation.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–8 and Supplementary Table 1 (PDF 1617 kb)
Rights and permissions
About this article

Cite this article
Choi, Y., Gullicksrud, J., Xing, S. et al. LEF-1 and TCF-1 orchestrate TFH differentiation by regulating differentiation circuits upstream of the transcriptional repressor Bcl6. Nat Immunol 16, 980–990 (2015). https://doi.org/10.1038/ni.3226
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ni.3226
This article is cited by
-
Aiolos represses CD4+ T cell cytotoxic programming via reciprocal regulation of TFH transcription factors and IL-2 sensitivity
Nature Communications (2023)
-
The TRIM37 variants in Mulibrey nanism patients paralyze follicular helper T cell differentiation
Cell Discovery (2023)
-
CTLA4 protects against maladaptive cytotoxicity during the differentiation of effector and follicular CD4+ T cells
Cellular & Molecular Immunology (2023)
-
TCF1 in T cell immunity: a broadened frontier
Nature Reviews Immunology (2022)
-
Identification of distinct functional thymic programming of fetal and pediatric human γδ thymocytes via single-cell analysis
Nature Communications (2022)
