
Abstract
De novo induction of Foxp3+ regulatory T cells (Tregs) is particularly efficient in gut-draining mesenteric and celiac lymph nodes (mLN and celLN). Here we used LN transplantations to dissect the contribution of stromal cells and environmental factors to the high Treg-inducing capacity of these LN. After transplantation into the popliteal fossa, mLN and celLN retained their high Treg-inducing capacity, whereas transplantation of skin-draining LN into the gut mesenteries did not enable efficient Treg induction. However, de novo Treg induction was abolished in the absence of dendritic cells (DC), indicating that this process depends on synergistic contributions of stromal and DC. Stromal cells themselves were influenced by environmental signals as mLN grafts taken from germ-free donors and celLN grafts taken from vitamin A–deficient donors did not show any superior Treg-inducing capacity. Collectively, our observations reveal a hitherto unrecognized role of LN stromal cells for the de novo induction of Foxp3+ Tregs.
Similar content being viewed by others


Introduction
The intestinal mucosa is constantly exposed to a plethora of bacteria colonizing the gastrointestinal tract. The density of microbiota increases along the gastrointestinal tract and in the colon reaches >1012 bacteria per gram feces. Intestinal microbiota exert important functions for our well being by breaking down otherwise indigestible material, synthesizing vitamins, and inhibiting colonization by pathogenic microbes. To maintain this symbiotic relationship with our microbiota, the intestinal immune system mounts regulated immune responses, including the generation of regulatory T cells (Tregs) expressing the lineage specification factor Foxp3. A wealth of data have demonstrated the central role of Foxp3+ Tregs in immune tolerance to self and innocuous foreign antigens.1 The majority of Foxp3+ Tregs is generated during thymic development, shows features of a stable T-cell lineage, and is believed to be selected for the recognition of self-antigens.2 Still, in the intestine the Treg repertoire needs to be complemented by specificities directed against commensal microbiota and food antigens,3 and this expansion of the Foxp3+ Treg pool is achieved by peripheral conversion of conventional naive Foxp3−CD4+ T cells into Foxp3+ Tregs.4, 5 Indeed, this de novo induction of Foxp3+ Tregs is particularly efficient in the intestinal immune system,6, 7, 8 and Tregs present in the colonic mucosa are induced in response to commensal microbiota.9 Yet, the inter-relation of immune regulatory processes and intestinal microbiota is even more complex: beside Treg specificities directed against microbiota, metabolic activities of commensal bacteria can also promote de novo Foxp3+ Treg induction.10
This intimate inter-relation of Treg induction, immune regulation, and intestinal microbiota requires a dedicated cellular and molecular set up in the intestinal immune system. A key factor during the peripheral generation of Foxp3+ Tregs is transforming growth factor-β (TGF-β) that leads to the direct induction of Foxp3 expression,4 a process that is further facilitated by addition of interleukin (IL)-2 and/or the vitamin A metabolite retinoic acid (RA).7, 11 TGF-β and RA are prominently produced in the intestine, and the high capacity of intestinal dendritic cells (DC) to produce these factors is thought to drive the efficient de novo generation of Foxp3+ Tregs within mesenteric lymph nodes (mLN).7, 11 In particular, CD103+ DC within mLN and small intestine possess high levels of the RA-synthesizing enzyme retinal aldehyde dehydrogenase (RALDH)-27, 12, 13, 14 and thus possess a high Treg-inducing capacity in vitro.7, 11, 15 Yet, RALDH activity is not restricted to CD103+ intestinal DC,13 and inflammatory responses have been shown to terminate RALDH activity and tolerogenic properties of CD103+ DC in the intestinal immune system.16
More recent studies suggest that in addition to migratory CD103+ DC, non-hematopoietic sessile mLN stromal cells also contribute to the higher RA levels present in mLN.17 They express substantially higher levels of RALDH compared with stromal cells in skin-draining peripheral LN (pLN), and RA derived from mLN stromal cells can directly induce the expression of gut-homing molecules on activated T cells.12, 18 In the present study, we report on LN transplantation experiments revealing an important contribution of stromal cells for the efficient induction of Foxp3+ Tregs within gut-draining LN. Additionally, we show that stromal cells themselves acquire their tolerogenic properties by unique micro-environmental factors, namely commensal microbiota for gut-draining mLN and vitamin A for liver-draining celiac LN (celLN).
Results
Efficient de novo induction of Foxp3+ Treg in gut-draining LN
Previous studies showed enhanced induction of Foxp3+ Tregs after oral antigen feeding in gut-draining mLN compared with pLN,6, 11 with a particularly high induction capacity in liver-draining celLN.8 To uncouple LN-intrinsic properties from preferential accumulation of antigen in gut-draining LN following its oral application, we studied induction of Foxp3+ Tregs in different LN after systemic antigen application via the intravenous (i.v.) route. Thereto, Foxp3−CD4+ T cells from ovalbumin (Ova)-specific, T cell receptor (TCR)-transgenic DO11.10 mice were adoptively transferred to wild-type (WT) BALB/c recipient mice followed by i.v. injection of Ova peptide. Three days after antigen application, proliferation of adoptively transferred Ova-specific CD4+ T cells was comparable in pLN, mLN, and celLN (Figure 1a), suggesting that i.v. injection led to an equivalent distribution of antigenic peptide at these sites. When we determined the frequency of Foxp3+ cells among adoptively transferred CD4+ T cells, we observed that both gut-draining LN, celLN and mLN, showed a higher Treg-inducing capacity compared with pLN, with celLN being the most efficient site (Figure 1a). Thus, the differential capacity of celLN and mLN to de novo induce Foxp3+ Tregs, which could be observed after oral antigen feeding (Figure 1c), was still preserved after systemic antigen application, albeit at a lower level. This indicates that unique intrinsic properties of gut-draining LN contribute to their high Treg-inducing capacity.

Mesenteric and celiac lymph nodes (mLN and celLN) display a higher regulatory T cell (Treg)-inducing potential compared with peripheral LN (pLN) even after systemic antigen application. (a) Foxp3−CD62LhighCD4+ T cells from DO11.10xFoxp3gfp reporter mice were labeled with CPDViolet and adoptively transferred into BALB/c mice. Subsequently, recipients were immunized via repetitive intravenous (i.v.) injections of ovalbumin (Ova)323-339 peptide starting the next day and analyzed 3 days after the first immunization. Representative dot plots show purities of naive cells before transfer (ungated) and Foxp3gfp expression vs. proliferation of reanalyzed Ova-TCR+CD4+ cells from the indicated organs. Numbers indicate frequencies of gated Foxp3+ cells. (b) The graph depicts frequencies of Foxp3+ Tregs among transferred Ova-TCR+CD4+ cells for the indicated organs. Data from one representative out of three independent experiments are shown. Symbols represent individual mice (n=18), and lines indicate median. (c) BALB/c mice received Foxp3−CD62LhighCD4+ T cells from DO11.10xFoxp3gfp reporter mice and recipients were orally immunized via continuous delivery of Ova ad libitum via the drinking water (p.o.). Graph shows frequencies of Foxp3+ Tregs among transferred Ova-TCR+CD4+ cells for the indicated organs 6 days after the start of Ova feeding. Pooled data from two independent experiments are shown. Symbols represent values from individual mice (n=12), and lines indicate median. TCR, T-cell receptor.
Stromal cells determine tolerogenic properties of transplanted LN
To study the contribution of stromal cells to the Treg-inducing capacities of LN in vivo, we performed LN transplantations. First, we grafted either pLN or mLN into the gut mesenteries after excision of endogenous mLN. We have previously shown that as early as 8 weeks after transplantation hematopoietic cells of transplanted LN are completely replaced by hematopoietic cells of recipient origin, whereas graft-derived stromal cells are largely retained in the transplant.12, 19 Eight weeks after transplantation, Ova-specific CD4+ T cells were transferred into transplant-recipients and antigen was applied via the natural route by oral feeding. Four days after the start of antigen feeding, we observed a significantly higher frequency of Foxp3+ Tregs among adoptively transferred Ova-specific CD4+ T cells in transplanted mLN compared with transplanted pLN (Figure 2a). No differences in T-cell proliferation of transferred cells (Figure 2a) or in frequency of endogenous Foxp3+ Tregs were observed (data not shown). Of note, transplanted pLN still showed a substantial population of de novo induced Foxp3+ Tregs, suggesting that their Treg-inducing capacity is positively influenced by drainage from mucosal tissue and stromal cells are only one contributing factor supporting Treg induction in mLN.

Tolerogenic property of lymph nodes (LN) is imprinted. (a) Mesenteric LN (mLN) or peripheral LN (pLN) fragments were grafted into the gut mesenteries after excision of endogenous mLN. After at least 8 weeks of reconstitution, carboxyfluorescein succinimidyl ester (CFSE)-labelled DO11.10 cells were adoptively transferred, followed by oral administration of ovalbumin (Ova) on 2 consecutive days. Foxp3 expression in mLN was analyzed 4 days after the first antigen feeding. Representative dot plots gated on Ova-TCR+CD4+ cells from the indicated LN are shown, and numbers indicate frequencies of gated Foxp3+ Tregs. Graph shows pooled data from four independent experiments (n=10–13). Circles represent individual mice, and lines indicate median. (b) LN were removed from the popliteal fossa and replaced by popliteal LN (popLN), mLN, or celiac LN (celLN) from BALB/c donor mice. After a minimum of 10 weeks of reconstitution, transplanted mice received Foxp3− cells isolated from DO11.10 donors. One day later, recipients were immunized via repetitive i.v. injection of Ova323-339 peptide and analyzed on day 3 after the first immunization. Graph summarizes frequencies of Foxp3+ Tregs among transferred Ova-TCR+CD4+ cells recovered from transplanted LN. Pooled data from seven independent experiments are shown. Symbols represent individual mice (n=18–20), and lines indicate median. FACS, fluorescence-activated cell sorter; NS, not significant; TCR, T-cell receptor.
To further pinpoint the contribution of LN stromal cells to the high Treg-inducing capacities of gut-draining LN, we transplanted celLN and mLN into the popliteal fossa after excision of the endogenous popliteal LN (popLN). Transplanted popLN served as controls. Similar to the aforementioned transplantation model, as early as 5 weeks after transplantation into this non-tolerogenic, skin-draining site transplanted LN were shown to consist of donor-derived stromal cells, whereas virtually all hematopoietic cells were recipient-derived.18 In the present study, after a minimum of 10 weeks of reconstitution, Ova-specific Foxp3−CD4+ T cells were transferred into transplant-recipients and Ova peptide was applied systemically via the i.v. route. Three days later, frequencies of Foxp3+ Tregs among adoptively transferred T cells were determined. When compared with transplanted popLN, gut-draining celLN and mLN retained their high Treg-inducing properties after transplantation (Figure 2b). In summary, both transplantation settings revealed that stromal cells from gut-draining LN support Foxp3+ Treg induction irrespective of the LN localization. Such activity was present in transplanted LN several weeks after surgery, suggesting that stromal cells are stably imprinted with unique ‘site-specific’ properties.
Differential expression of IL-6 and RALDH-encoding genes does not explain the differences in Treg induction in mLN and pLN
To identify factors that might explain the site-specific differences in Treg induction, we compared the global cytokine milieu in transplanted LN. Although we noted some subtle differences, no overt bias towards a more pro-inflammatory cytokine milieu in transplanted pLN compared with transplanted mLN was observed (Supplementary Figure S1). However, RALDH-encoding genes were previously reported to show higher expression in mLN stromal cells and might positively influence Treg induction.12, 18 In addition, we noted here a higher expression of β8-integrin in stromal cells isolated from mLN compared with pLN (Figure 3a), which might affect the biological activity of TGF-β1.14 Indeed, latent TGF-β was slightly more potent to induce Foxp3+ Tregs in vitro in the presence of mLN stromal cells compared with pLN stromal cells (Supplementary Figure S2). Furthermore, the recently described higher levels of IL-6 expression in pLN compared with mLN stromal cells20, 21 might negatively influence Treg induction. To specifically address the function of stromal cell–derived IL-6, we compared the Treg-inducing capacity of WT and IL-6-deficient pLN transplanted into the gut mesenteries. However, we did not note any differences in the frequency of Foxp3+ Tregs among adoptively transferred TCR-transgenic cells (Figure 3b).

Role of Itgb8, Il6, and RALDH (retinal aldehyde dehydrogenase) activity for lymph nodes (LN) stromal cells. (a) Expression of Itgb8 normalized to Hprt expression was assessed in primary FACS (fluorescence-activated cell sorter)-sorted CD45−CD24− stromal cells derived from peripheral LN (pLN) or mesenteric LN (mLN) via real-time PCR. Each bar represents mean±s.d. from three independent experiments. (b) Endogenous mLN were excised from mesenteries and replaced by pLN from either wild type or Il6-deficient donors. After reconstitution, CD4+ T cells from congenic, T-cell receptor (TCR)-transgenic OT-II mice were adoptively transferred followed by oral administration of ovalbumin on 2 consecutive days. Foxp3 expression in congenic CD45.1+CD4+ T cells was analyzed in the transplanted LN 4 days after the start of antigen feeding. Bars represent mean±s.d. of four (Il6+/+) and eight mice (Il6−/−). Please note that conversion efficacy is consistently lower in OT-II cells on the C57BL/6 background compared with DO11.10 cells on the BALB/c background. Still, in both genetic systems conversion efficacy is higher in transplanted mLN compared with pLN (data not shown). (c) Foxp3−CD4+ T cells were sorted from Foxp3-IRES-GFP mice and activated in vitro using CD3/CD28-coated beads in the presence or absence of pLN- or mLN-derived stromal cells. Cells were cultured for 5 days in serum-free medium in the absence (left) or presence (right) of transforming growth factor (TGF)-β1, and retinol was added as indicated. At day 5, Foxp3 expression among CD4+ T cells was determined by flow cytometry. Each symbol indicates mean measurement obtained from analysis of duplicate or triplicate wells analyzed in independently performed experiments. IL, interleukin.
To test whether the higher Treg-inducing capacity of mLN stromal cells relies on their differential expression of RALDH-encoding genes, we performed in vitro studies, because no viable mouse strains are available to test this pathway in the transplantation set-up. Thereto, naive Foxp3−CD4+ T cells were stimulated in vitro using CD3/CD28-coated beads in the presence of stromal cells derived from pLN or mLN. To exclude any influence of serum-associated retinoids, all assays were performed in serum- and retinoid-free medium containing IL-2. In the absence of any additional reagents and/or cells, the frequency of induced Foxp3+ Tregs was very low (Figure 3c). Addition of stromal cells derived from pLN did not reveal any increase of Foxp3+ Tregs, but co-cultures of naive T cells with mLN-derived stromal cells showed a modest but statistically significant increase in Treg frequencies. Importantly, addition of retinol did not lead to a further increase, and similar results were obtained in the presence of TGF-β, where generally higher levels of induced Tregs were observed (Figure 3c). Together, these results suggest that none of the so far tested candidates convincingly explained the differences in the Treg-inducing capacity observed in the LN transplantation experiments.
Stromal cell–DC cooperation shapes Foxp3+ Treg-inducing capacity of LN
As stromal cells did not exert strong direct effects on Treg induction in vitro, we speculated that in vivo stromal cells might also affect T-cell responses indirectly through modulation of DC functions. We first asked whether Treg could be induced in vivo in the absence of DC and used CD11cDTR mice22 to selectively deplete CD11c+ DC. Although DC depletion had little influence on peptide-induced proliferation of adoptively transferred antigen-specific T cells, we observed that induction of Foxp3+ Tregs was nearly completely abrogated after DC depletion (Figure 4). No side effects of diphtheria toxin on Foxp3+ Treg induction were seen in control mice. Together, these findings suggest that DC are absolutely required for in vivo Treg induction.

Dendritic cells have a key role for the de novo induction of regulatory T cells (Tregs). (a) Lethally irradiated BALB/c mice were reconstituted with bone marrow (BM) isolated from CD11cDTR or BALB/c mice to generate CD11cDTR→WT (wild type) or WT→WT chimeras, respectively. After at least 16 weeks of reconstitution, mice were treated with diphtheria toxin (DT) every second day, and controls received phosphate-buffered saline (PBS). One day after the first treatment, mice received Foxp3−CD62LhighCD4+ T cells from DO11.10xFoxp3gfp reporter mice, were immunized via repetitive i.v. injection of ovalbumin (Ova)323-339 peptide starting the next day, and were analyzed 3 days after the first immunization. Representative dot plots show Foxp3gfp expression vs. proliferation of Ova-TCR+CD4+ cells recovered from mesenteric lymph nodes corresponding to the indicated treatment. Numbers indicate frequencies of gated Foxp3+ Tregs. (b) Frequencies of Foxp3+ Tregs among Ova-TCR+CD4+ cells are shown for the indicated chimeras and treatments. Circles represent individual control WT→WT chimeras and triangles individual CD11cDTR→WT chimeras. Closed symbols indicate PBS-treated and open symbols DT-treated chimeras. Pooled data from three independent experiments are shown (n=8), and lines indicate median. FACS, fluorescence-activated cell sorter.
One might thus speculate that the differential Treg-inducing capacity in transplanted LN might be simply due to different frequencies and/or subtypes of DC. However, we did not observe any drastic differences in the frequency of CD11chigh DC between popLN, celLN, and mLN transplanted into popliteal fossa, and also the frequency of CD103+ DC was comparable (Figure 5a). Furthermore, the frequency of TCR-transgenic (KJ+) T cells did not differ between the various transplants. Together, these findings indicate that the ratios of DC to antigen-specific T cell are unchanged and cannot explain the higher Treg-inducing capacity in transplanted gut-draining LN.

Unchanged ratios of dendritic cells (DC) to transgenic T cells in transplanted lymph nodes (LN). Mice were treated as described in Figure 2b. About 16 weeks after LN transplantation, transplanted LNs were analyzed for frequencies of (a) CD19−B220−CD11chigh cells (cDCs), (b) CD103 expression on cDCs, and (c) Ova (ovalbumin)-TCR+ (KJ+) among CD4+ T cells via flow cytometry. Pooled data (n=4–6) from two independent experiments are shown. Dots represent data obtained from individual mice, and lines indicate median. Statistical testing was performed via one-way analysis of variance with Tukey‘s post test. celLN, celiac lymph nodes; mLN, mesenteric lymph nodes; NS, not significant; popLN, popliteal lymph nodes.
Commensal microbiota and vitamin A instruct the tolerogenic capacity of mLN and celLN stromal cells
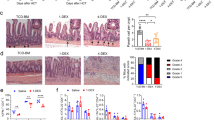
After establishing that stromal cells from gut-draining LN support efficient Treg induction, we wanted to identify factors that imprint such tolerogenic properties within the stromal cells. Potential candidate factors that profoundly affect intestinal immune functions include commensal microbiota and vitamin A–derived metabolites. We first analyzed Treg induction in mice, in which the commensal microbiota had been reduced by treatment of adult specific pathogen-free (SPF)-housed mice with a cocktail of antibiotics. Ova-specific Foxp3−CD4+ T cells were adoptively transferred, followed by antigen feeding via the drinking water. Six days later, comparable frequencies of Foxp3+ Tregs among adoptively transferred cells were present in mLN and celLN of antibiotics-treated mice and untreated controls (Figure 6a), suggesting that during the period of Treg induction the commensal microbiota had only little influence. However, when we transplanted LN from adult germ-free mice into the popliteal fossa of SPF-housed recipients and tested their Treg-inducing capacity 20 weeks later by adoptive T-cell transfer and systemic antigen application, we observed a significantly reduced frequency of induced Foxp3+ Tregs within transplanted mLN taken from germ-free mice compared with SPF controls (Figure 6b). Interestingly, comparable Treg frequencies were found within transplanted celLN taken from germ-free and SPF control mice, suggesting that the high Treg-inducing capacity of celLN is not influenced by the commensal microbiota. Notably, we observed no overt change in the total number of stromal cells comparing germ-free and SPF mice (Supplementary Figure S3), indicating that differences in stromal cell numbers are unlikely to bias our results.

Commensal microbiota shape tolerogenic property of mesenteric lymph nodes (mLN) stromal cells only during ontogeny. (a) Adult specific pathogen-free (SPF)-housed BALB/c mice were treated with antibiotics for a period of approximately 8 weeks. Subsequently, Foxp3− cells from DO11.10 mice were adoptively transferred into antibiotics-treated or untreated BALB/c mice. One day later, ovalbumin (Ova) was supplied ad libitum via the drinking water, and the mice were analyzed 6 days after onset of Ova feeding. Graph shows frequencies of Foxp3+ cells among transferred Ova-TCR+CD4+ cells for the indicated organs of untreated controls (filled circles, n=12) and antibiotics-treated mice (open circles, n=16). Circles represent individual mice from two independent experiments, and lines indicate median. Statistical analysis was performed via an unpaired t-test. (b) Popliteal lymph nodes (popLN) were replaced by celiac lymph nodes (celLN), mLN, or popLN from SPF-housed or germ-free donor mice. After a reconstitution time of about 20 weeks, transplanted mice received Foxp3−cells from DO11.10 mice. One day later, recipients were immunized via repetitive i.v. injection of Ova323-339 peptide and analyzed 3 days after the first immunization. No differences in the degree of T-cell proliferation were observed between the groups (data not depicted). Graph shows frequencies of Foxp3+ cells among transferred Ova-TCR+CD4+ cells for the indicated groups. Pooled data from five independent experiments (n=9–14) are shown, and lines indicate median. Statistical analysis was performed via an unpaired t-test. FACS, fluorescence-activated cell sorter; NS, not significant; TCR, T-cell receptor.
To address the role of vitamin A on the imprinting of stromal cells, we generated vitamin A–deficient mice and transplanted their LN into the popliteal fossa of standard diet–fed recipients. Eight-to-ten weeks after LN transplantation, their Treg-inducing capacity was analyzed as described above. Whereas the Treg-inducing capacity of transplanted mLN was not influenced by vitamin A deficiency, transplanted celLN taken from vitamin A–deficient donors displayed a significantly reduced frequency of induced Foxp3+ Tregs when compared with control celLN (Figure 7). Together, these findings suggest that commensal microbiota and vitamin A have reciprocal roles in the shaping of tolerogenic properties of stromal cells in mLN and celLN, respectively.

Vitamin A instructs tolerogenic capacity of celiac lymph nodes (celLN) but not mesenteric LN (mLN) stromal cells. Popliteal LN (popLN) were replaced by celLN, mLN, or popLN taken from vitamin A–deficient donor mice and LN taken from standard diet-fed mice served as controls. After a reconstitution time of about 8–10 weeks, transplanted mice received Foxp3– cells from DO11.10 mice. One day later, recipients were immunized via repetitive i.v. injection of ovalbumin (Ova)323-339 peptide and analyzed 3 days after the first immunization. No differences in the degree of T-cell proliferation were observed between the groups (data not depicted). Graph shows frequencies of Foxp3+ cells among transferred Ova-TCR+CD4+ cells for the indicated groups. Pooled data from four independent experiments (n=6–11) are shown, and lines indicate median. Statistical analysis was performed via an unpaired t-test. FACS, fluorescence-activated cell sorter; NS, not significant; TCR, T-cell receptor; VAD, vitamin A-deficient.
Discussion
Foxp3+ Tregs are key to the intestine’s ability to balance tolerance towards microbiota and dietary antigens on the one hand and vigorous immunity to pathogens on the other hand. Here, we show that in the intestinal immune system commensal microbiota and vitamin A prewire LN stromal cells to support tolerance induction in the gut.
Previous work emphasized distinct properties of DC subsets to support mucosal tolerance. Indeed, we show here, for the first time, that in vivo Foxp3+ Treg induction essentially requires the presence of CD11c+ DC. We found that DC depletion did not impair T-cell proliferation, indicating that in response to i.v. peptide injection other cells types, potentially including B cells, are sufficient to induce T-cell proliferation but insufficient to generate Foxp3+ Tregs. Foxp3 induction may require unique cell contact–triggered signals, such as low costimulatory signals provided by antigen-presenting DC. Yet, additional DC-produced factors may also support Treg induction in a paracrine manner. In fact, RA injected subcutaneously resulted in induction of gut-homing molecules CCR9 (C-C motif chemokine receptor 9) and α4β7-integrin in popLN,23 indicating that at least RA can act in a paracrine manner and may affect T-cell responses throughout a given compartment.
The tolerogenic function of DC does not seem to be a DC-intrinsic property but appears to be dynamically regulated by the local tissue micro-environment. In the intestine, gut epithelial cells produce TGF-β and RA that condition DC to support tolerance induction.24, 25 Here, we follow the hypothesis that within gut-draining LN tolerogenic properties of DC are further solidified by the action of stromal cells. Again, RA might be one of the decisive factors in this process. RALDH expression, in particular RALDH1 and RALDH3, is higher in mLN compared with pLN stromal cells,12 and RALDH activity can be induced in DC by RA itself.26 Indeed, bone marrow (BM) DC acquired higher RALDH expression when injected into transplanted LN (data not shown). However, no difference was found in this respect between transplanted pLN and mLN, and upregulation of RALDH in DC entering the mLN could not be explained solely by the activity of stromal cells. Thus, there is a picture emerging that RA levels in mLN are the product of stromal cells plus DC, which synergistically interact to maintain high RA levels throughout the mLN compartment and preset the mLN environment to a tolerogenic state. Besides RALDH, we tested two additional prime candidates potentially modulating Foxp3+ Treg generation, TGF-β and β8-integrin. Whereas Tgfb1 expression in stromal cells was generally low and did not differ between pLN and mLN (data not shown), Itgb8 expression was markedly higher in mLN compared with pLN stromal cells. TGF-β is produced in a latent form that needs to be activated to exert its biological function,27 and recent data suggest that the ability of intestinal CD103+ DC to drive Foxp3+ Treg induction in the absence of exogenous TGF-β is linked to their β8-integrin-dependent, TGF-β-activating function.14, 28 Thus, besides their RA-producing function we here identified another factor that is differentially expressed in pLN and mLN stromal cells and that might contribute to enhance Foxp3+ Treg induction in the intestinal immune system.
Stromal cells represent a heterogeneous array of different cell types.17 Based on their anatomical location in the T-cell zone, fibroblastic reticular cells, which are defined as gp38+ (podoplanin) CD31− cells, might be the main population acting on incoming DC. Upon stimulation, fibroblastic reticular cells secrete various cytokines and chemokines, which critically contribute to T-cell homeostasis.29, 30 A recent array analysis of stromal cells from mLN and pLN reported the differential expression of IL-6 and of the cell fate–determining homeodomain transcription factor NKX2.3 as well as several other more subtle differences.20, 21 Thus, it is tempting to speculate that LN-specific differences in fibroblastic reticular cells are an important factor contributing to the different Treg-inducing capacities of pLN and mLN by modulating the tolerogenic phenotype of incoming DC. Nevertheless, IL-6 is not a decisive factor negatively influencing the Treg-inducing capacity of pLN, as WT and IL-6-deficient pLN transplanted into the gut mesenteries displayed similar frequencies of de novo induced Foxp3+ Tregs.
The tolerogenic properties of LN stromal cells are stably maintained after transplantation. In fact, Treg-inducing capacities did not differ in transplanted LN analyzed up to 20 weeks after transplantation. This stable tolerogenic state may be achieved by developmental programming, but also epigenetic changes are an attractive explanation for the observed stability of LN stromal cell functions. Alternatively, exogenous signals may modulate tissue-specific properties of stromal cells. In favor of this idea, the tolerogenic function of LN stromal cells was acquired in response to micro-environmental factors, i.e., commensal microbiota and vitamin A. Unexpectedly, celLN and mLN differed in their mode of imprinting. Whereas mLN taken from germ-free mice did not support Foxp3+ Treg conversion, corresponding celLN showed unimpaired tolerogenic properties. A reciprocal scenario was observed with respect to the role of vitamin A: vitamin A deficiency selectively affected the tolerogenic properties of celLN but not mLN stromal cells. Intriguingly, microbial densities increase along the intestinal axis, and encounter of microbiota-derived stimuli might be more intimate in mLN compared with celLN. Conversely, vitamin A is largely stored in the liver,31 which might contribute to the predominant influence of vitamin A in celLN but not mLN.
In the steady state, stromal cells in adult mice show little proliferation and maintain their phenotype without ongoing instructive signals. Once mLN stromal cells had been exposed to microbiota-derived signals in early life, they acquire a tolerogenic state, which cannot be reverted by antibiotic eradication of the commensal microbiota in the adulthood. Thus, stromal cells seem to constitute a stabilizing element for mucosal tolerance induction. This is in contrast to DC; lamina propria DC show a rapid turnover, constitutively travel to mLN, and are re-placed by DC progenitors on a permanent basis. Thus, newly generated DC need to rapidly gain tolerogenic properties. Within the intestinal lamina propria of healthy individuals, several cell types, in particular epithelial cells, instruct incoming DC.24, 25 We speculate that this process might be ongoing in gut-draining LN where stromal cells further modulate the tolerogenic properties of DC. Thus, stromal cells might turn out as attractive therapeutic targets to modulate intestinal tolerance.
In conclusion, the present study identifies resident stromal cells as novel players in tissue-specific induction of tolerance. In addition, we show that the phenotype of stromal cells is not inherited but may be strongly influenced by micro-environmental factors that contribute to the tolerogenic capacity of gut-draining LN.
Methods
Mouse strains
OT-II, Foxp3-IRES-GFP,32 B6;129S2-Il6tm1Kopf/j, C57BL/6, and CD45.1 congenic mice were bred at the Hannover Medical School (MHH, Hannover, Germany). BALB/c and C57BL/6 mice were purchased from Harlan Laboratories (Venray, The Netherlands), Charles River (Sulzfeld, Germany) or Janvier (Le Genest-Saint-Isle, France). DO11.10 mice were obtained from the breeding facilities of the Bundesanstalt für Risikobewertung (BfR, Berlin, Germany), the Helmholtz Centre for Infection Research (HZI, Braunschweig, Germany), or the MHH. CD11cDTRxBALB/c mice22 were bred at the HZI. DO11.10xFoxp3gfp reporter mice33 were kindly provided by Alexander Rudensky (Memorial Sloan-Kettering Cancer Center, New York, NY, USA) and Ludger Klein (Institute for Immunology, LMU, Munich, Germany) and were bred at the HZI. Germ-free BALB/c mice were generated at the MHH by Cesarean section and were housed under axenic conditions in sterile incubators. All mice other than germ-free mice were maintained under SPF conditions in isolated ventilated cages. Water and Ova-free diet were supplied ad libitum. In all experiments, gender- and age-matched mice were used. The experiments were performed in accordance with the institutional, state, and federal guidelines.
Antibodies and flow cytometry
Fluorochrom-conjugated anti-CD4 (RM4-5), anti-CD4 (RmCD4.2), anti-CD11c (N418), anti-CD24 (M1/69), anti-CD25 (PC61.5), anti-CD45.1 (A20), anti-CD45.2 (104), anti-CD62L (MEL14), anti-CD86 (GL-1), anti-CD103 (M290), anti-Vβ5.1/5.2 (MR9-4), anti-MHCII IAb (AF6-120.1), anti-Ova-TCR (KJ1.26), anti-gp38 (8.1.1), Ter119 (TER-119), anti-Foxp3 (FJK-16s), and Streptavidin were purchased from eBioscience (San Diego, CA), BD (Heidelberg, Germany), Invitrogen (Darmstadt, Germany), and Biolegend (San Diego, CA) or produced and conjugated in our laboratory. Intracellular Foxp3 staining was performed according to the manufacturer’s instructions. Before analysis of unfixed cells, DAPI (4,6-diamidino-2-phenylindole; Sigma-Aldrich, St Louis, MO) was added at 50 ng ml−1 for exclusion of dead cells. In some experiments, dead cells were detected with LIVE/DEAD Fixable Dead Cell Stain (Invitrogen) or via scatter properties. Flow cytometry was performed using an LSRII (BD Biosciences, Heidelberg, Germany) flow cytometer with Diva software (BD), and data were analyzed with FlowJo software (TreeStar, Ashland, OR).
T-cell isolation and adoptive transfer
Transferred cells were purified from the spleens and LN of DO11.10 donors by magnetic enrichment of CD103−CD25−CD62Lhigh cells (Miltenyi Biotec, Bergisch-Gladbach, Germany) or from the spleens and LN of DO11.10xFoxp3gfp donors by a combination of MACS (magnetic-activated cell sorter) and FACS (fluorescence-activated cell sorter) sorting (FACSAria™ II, BD) to purify CD4+Foxp3gfp−CD62Lhigh cells. The resulting target population contained <0.5% Foxp3+ cells. Sorted cells were labeled with cell proliferation dyes (Invitrogen), and approximately 2.5 to 10 × 106 cells were injected i.v. per recipient mouse. In a few experiments, non-purified cell suspensions generated from LN of DO11.10 donors were transferred and transgenic cells subsequently identified by anti-TCR staining.
Immunization
For in vivo Foxp3+ Treg induction following the oral route, Ova protein was applied via the gastrointestinal tract either continuously in sterile drinking water (10 mg ml−1) or by gavage of two single doses of 50 mg Ova 1 and 2 days after adoptive T-cell transfer. For in vivo Foxp3+ Treg induction following the systemic route, 20 μg Ova323-339 peptide was injected i.v. on 2 consecutive days, starting 1 day after adoptive transfer of Foxp3− cells. At the indicated time points after immunization, cells were isolated from LN transplants and stained for surface markers and intracellular Foxp3 expression.
LN transplantations
For transplantations into the popliteal fossa, BALB/c-recipient mice were anesthetized with ketamine (WDT, Garbsen, Germany) and xylacine (CP Pharma, Burgdorf, Germany), the skin of the popliteal fossa of the right hind leg was opened, and the endogenous popLN and surrounding fat tissue were removed. popLN, mLN, or celLN dissected from conventionally housed SPF, germ-free, or vitamin A–deficient BALB/c donor mice were placed into the popliteal fossa, and the cut was sewn with absorbable suture (Catgut, Markneukirchen, Germany). LN transplantations into the mesenteries were performed as previously described.12, 19 Before subjecting to further experimental procedure, recipients were housed for a minimum of 8 weeks to ensure restoration of lymphatic connections to the LN.
Cytokine determination
Eight weeks post transplantation into the popliteal fossa, transplanted mLN and popLN were detected injecting 10 μl of Patent blue V sodium salt (25 mg ml−1, Sigma-Aldrich) in H2O, excised, and fat tissue removed. Protein lysates were generated with the Bio-Plex Cell Lysis Kit (Bio-Rad, Munich, Germany) according to the manufacturer’s instructions, and protein concentrations were determined by applying the Pierce BCA Protein Assay Kit (Thermo Scientific, Rockford, IL). Protein lysate concentrations were adjusted to 1 mg ml−1 with Bio-Plex Sample Diluent (Bio-Rad). Cytokine determination was performed with the Bio-Plex Pro Mouse Cytokine 23-plex Assay (Bio-Rad), measured by BioPlex 200 (Bio-Rad), and analyzed with the software Bio-Plex Manager (Bio-Rad).
Stromal cell–T cell co-cultures
Either pLN (pool of inguinal, brachial, axillary LN) or mLN pooled from three mice were cut into small pieces and digested for 1 h at 37 °C in RPMI 1640 (Invitrogen) supplemented with 0.1 mg ml−1 Liberase Blendzyme 2 and 40 U ml−1 DNase I (both Roche, Grenzach-Wyhlen, Germany) with slow shaking. Cells were released from the LN via repeated pipetting and filtering through a 100-μm Cell Strainer (BD Biosciences). Digested LN cell suspensions were cultured for 10 days, and medium was exchanged every second day to remove non-adherent cells. We routinely confirmed that our stromal cell preparations contained >99% CD45-negative cells. Polyclonal Foxp3−CD4+ T cells were purified from the spleens and LN of Foxp3-IRES-GFP mice by a combination of MACS and FACS sorting (see above). A total of 105 sorted T cells were added to 2 × 104 stromal cells and activated with CD3/CD28-coated beads (Invitrogen) and 300 U ml−1 rmIL-2 (Roche) in the absence or presence of 10 ng ml−1 rhTGF-β1 (PeproTech, Hamburg, Germany) or 50 nM retinol (Sigma-Aldrich). All cultures were performed in serum- and vitamin A–free medium (X-VIVO 15, BioWhittaker, Lonza, Cologne, Germany).
BM chimeras and in vivo DC depletion
BALB/c-recipient mice received an antibiotic regimen of 100 μg ml−1 enrofloxacine (Bayer, Leverkusen, Germany) in sterile drinking water ad libitum for 1 week. Recipients were then irradiated at a single dose of 10.5 Gy by a gamma ray irradiation device (Rad Source Technologies, Suwanee, GA). One day later, the BM was isolated from tibia and femur bones of WT BALB/c or CD11cDTR donor mice, and Thy1.2+ cells were removed by sensitive MACS sorting (Miltenyi Biotec). A total of 5 × 106 Thy1.2− BM cells from CD11cDTR or BALB/c mice were injected i.v. into irradiated BALB/c recipients to generate CD11cDTR→WT and WT→WT control BM chimeras. After continuous supply of enrofloxacine for an additional 4 weeks, mice were subjected to experimental procedure after at least 20 weeks of reconstitution. CD11cDTR→WT and WT→WT chimeras received 8 ng per gram body weight diphtheria toxin (Sigma) every second day via the intraperitoneal route. As controls, chimeras were injected with phosphate-buffered saline. One day after the first treatment, naive T cells were adoptively transferred, and recipient mice were immunized as described above.
Stromal cell isolation
For isolation of stromal cells, LN cell suspensions were generated as described above, and hematopoietic cells were removed from the digested cell suspensions using anti-CD45 MicroBeads (Miltenyi Biotec). The negative fraction was stained with antibodies and FACS-sorted as CD45−CD24−gp38+.
Quantitative RT-PCR analysis
RNA was isolated from sorted stromal cells using RNeasy Mini or Micro Kit (Qiagen, Hilden, Germany). cDNA was synthesized using random hexamer primers and SuperScript II or SuperScript III reverse transcriptase (both Invitrogen). Real-time PCR was performed using LightCycler 2.0 (Roche) with SYBR green (SYBR Premix Ex Taq, Takara Bio, Otsu, Japan). The following primers were used: Itgb8 (f: 5′-ACAGCATCGCATGGACCAA-3′, r: 5′-AAGCAACCCGATCAAGAATGTG-3′) and Hprt (f: 5′-TCAGTCAACGGGGGACATAAA-3′, r: 5′-GGGGCTGTACTGCTTAACCAG-3′).
Antibiotics treatment
BALB/c mice formerly housed under SPF conditions were treated with an antibiotic regimen by feeding of ampicillin (1 g l−1; Ratiopharm, Ulm, Germany), ciprofloxacin (200 mg l−1; Bayer Vital, Leverkusen, Germany), imipenem (250 mg l−1; MSD, White House Station, NJ), metronidazole (1 g l−1; Fresenius, Bad Homburg, Germany), and vancomycin (500 mg ml−1; Cell Pharm, Hannover, Germany) ad libitum via the drinking water. Mice were housed in sterile cages, which were changed daily for the first 2 weeks. Efficient bacterial reduction was monitored by weekly bacteriological evaluation of feces from individual mice.34 Lack of any cultivable bacteria (aerobic and anaerobic cultures of Gram-negative rods, Gram-positive rods, and Gram-positive cocci) for at least 2 consecutive weeks were taken as an indication of the removal of detectable commensal microbiota.
Generation of vitamin A–deficient mice
Seven days after mating of SPF-housed BALB/c mice, one half of pregnant mice were put on a vitamin A–deficient diet (Ssniff; <120 IU vitamin A kg−1), and the other half were kept on normal diet containing 15,000 IU vitamin A kg−1 (control group). Offsprings continuously received specific diets for a minimum of 8 weeks until LN transplantations were performed.
Statistical analysis
Prism software (GraphPad, La Jolla, CA) was utilized for statistical analysis and graphs. If not stated otherwise, statistical significance was assessed via one-way analysis of variance and Tukey–Kramer post test. All tests were performed with the assumption of a two-tailed probability, and differences were considered statistically significant when P<0.05 and highly significant when P<0.01 or P<0.001 (*P<0.05; **P<0.01; ***P<0.001).
References
Sakaguchi, S., Miyara, M., Costantino, C.M. & Hafler, D.A. FOXP3+ regulatory T cells in the human immune system. Nat. Rev. Immunol. 10, 490–500 (2010).
Huehn, J., Polansky, J.K. & Hamann, A. Epigenetic control of FOXP3 expression: the key to a stable regulatory T-cell lineage? Nat. Rev. Immunol. 9, 83–89 (2009).
Strauch, U.G. et al. Influence of intestinal bacteria on induction of regulatory T cells: lessons from a transfer model of colitis. Gut 54, 1546–1552 (2005).
Curotto de Lafaille, M.A. & Lafaille, J.J. Natural and adaptive foxp3+ regulatory T cells: more of the same or a division of labor? Immunity 30, 626–635 (2009).
Haribhai, D. et al. A requisite role for induced regulatory T cells in tolerance based on expanding antigen receptor diversity. Immunity 35, 109–122 (2011).
Mucida, D., Kutchukhidze, N., Erazo, A., Russo, M., Lafaille, J.J. & Curotto de Lafaille, M.A. Oral tolerance in the absence of naturally occurring Tregs. J. Clin. Invest. 115, 1923–1933 (2005).
Coombes, J.L. et al. A functionally specialized population of mucosal CD103+ DCs induces Foxp3+ regulatory T cells via a TGF-beta and retinoic acid-dependent mechanism. J. Exp. Med. 204, 1757–1764 (2007).
Siewert, C. et al. Experience-driven development: effector/memory-like alphaE+Foxp3+ regulatory T cells originate from both naive T cells and naturally occurring naive-like regulatory T cells. J. Immunol. 180, 146–155 (2008).
Lathrop, S.K. et al. Peripheral education of the immune system by colonic commensal microbiota. Nature 478, 250–254 (2011).
Atarashi, K. et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 331, 337–341 (2011).
Sun, C.M. et al. Small intestine lamina propria dendritic cells promote de novo generation of Foxp3 Treg cells via retinoic acid. J. Exp. Med. 204, 1775–1785 (2007).
Hammerschmidt, S.I. et al. Stromal mesenteric lymph node cells are essential for the generation of gut-homing T cells in vivo. J. Exp. Med. 205, 2483–2490 (2008).
Guilliams, M. et al. Skin-draining lymph nodes contain dermis-derived CD103- dendritic cells that constitutively produce retinoic acid and induce Foxp3+ regulatory T cells. Blood 115, 1958–1968 (2010).
Worthington, J.J., Czajkowska, B.I., Melton, A.C. & Travis, M.A. Intestinal dendritic cells activate transforming growth factor-beta and induce Foxp3+ T regulatory cells via integrin alphavbeta8. Gastroenterology 141, 1802–1812 (2011).
Matteoli, G. et al. Gut CD103+ dendritic cells express indoleamine 2,3-dioxygenase which influences T regulatory/T effector cell balance and oral tolerance induction. Gut 59, 595–604 (2010).
Laffont, S., Siddiqui, K.R. & Powrie, F. Intestinal inflammation abrogates the tolerogenic properties of MLN CD103+ dendritic cells. Eur. J. Immunol. 40, 1877–1883 (2010).
Roozendaal, R. & Mebius, R.E. Stromal cell-immune cell interactions. Annu. Rev. Immunol. 29, 23–43 (2011).
Molenaar, R. et al. Lymph node stromal cells support dendritic cell-induced gut-homing of T cells. J. Immunol. 183, 6395–6402 (2009).
Ahrendt, M., Hammerschmidt, S.I., Pabst, O., Pabst, R. & Bode, U. Stromal cells confer lymph node-specific properties by shaping a unique microenvironment influencing local immune responses. J. Immunol. 181, 1898–1907 (2008).
Fletcher, A.L. et al. Reproducible isolation of lymph node stromal cells reveals site-dependent differences in fibroblastic reticular cells. Front. Immun. 2, 35 (2011).
Malhotra, D. et al. Transcriptional profiling of stroma from inflamed and resting lymph nodes defines immunological hallmarks. Nat. Immunol. 13, 499–510 (2012).
Jung, S. et al. In vivo depletion of CD11c+ dendritic cells abrogates priming of CD8+ T cells by exogenous cell-associated antigens. Immunity 17, 211–220 (2002).
Hammerschmidt, S.I. et al. Retinoic acid induces homing of protective T and B cells to the gut after subcutaneous immunization in mice. J. Clin. Invest. 121, 3051–3061 (2011).
Iliev, I.D. et al. Human intestinal epithelial cells promote the differentiation of tolerogenic dendritic cells. Gut 58, 1481–1489 (2009).
Iliev, I.D., Mileti, E., Matteoli, G., Chieppa, M. & Rescigno, M. Intestinal epithelial cells promote colitis-protective regulatory T-cell differentiation through dendritic cell conditioning. Mucosal Immunol. 2, 340–350 (2009).
Molenaar, R. et al. Expression of retinaldehyde dehydrogenase enzymes in mucosal dendritic cells and gut-draining lymph node stromal cells is controlled by dietary vitamin A. J. Immunol. 186, 1934–1942 (2011).
Annes, J.P., Munger, J.S. & Rifkin, D.B. Making sense of latent TGFbeta activation. J. Cell Sci. 116, 217–224 (2003).
Paidassi, H. et al. Preferential expression of integrin alphavbeta8 promotes generation of regulatory T cells by mouse CD103+ dendritic cells. Gastroenterology 141, 1813–1820 (2011).
Siegert, S. et al. Fibroblastic reticular cells from lymph nodes attenuate T cell expansion by producing nitric oxide. PLoS One 6, e27618 (2011).
Lukacs-Kornek, V. et al. Regulated release of nitric oxide by nonhematopoietic stroma controls expansion of the activated T cell pool in lymph nodes. Nat. Immunol. 12, 1096–1104 (2011).
Winau, F., Quack, C., Darmoise, A. & Kaufmann, S.H. Starring stellate cells in liver immunology. Curr. Opin. Immunol. 20, 68–74 (2008).
Wang, Y. et al. Th2 lymphoproliferative disorder of LatY136F mutant mice unfolds independently of TCR-MHC engagement and is insensitive to the action of Foxp3+ regulatory T cells. J. Immunol. 180, 1565–1575 (2008).
Fontenot, J.D., Rasmussen, J.P., Williams, L.M., Dooley, J.L., Farr, A.G. & Rudensky, A.Y. Regulatory T cell lineage specification by the forkhead transcription factor foxp3. Immunity 22, 329–341 (2005).
Heimesaat, M.M. et al. Gram-negative bacteria aggravate murine small intestinal Th1-type immunopathology following oral infection with Toxoplasma gondii. J. Immunol. 177, 8785–8795 (2006).
Acknowledgements
We thank Maria Ebel and Uta Lauer for expert technical assistance and Lothar Gröbe for cell sorting. This work was supported by the Deutsche Forschungsgemeinschaft (SFB621, SFB633, and PA 921/1-1).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declared no conflict of interest.
Additional information
SUPPLEMENTARY MATERIAL is linked to the online version of the paper
Rights and permissions
About this article
Cite this article
Cording, S., Wahl, B., Kulkarni, D. et al. The intestinal micro-environment imprints stromal cells to promote efficient Treg induction in gut-draining lymph nodes. Mucosal Immunol 7, 359–368 (2014). https://doi.org/10.1038/mi.2013.54
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mi.2013.54
This article is cited by
-
Gut–liver axis: barriers and functional circuits
Nature Reviews Gastroenterology & Hepatology (2023)
-
Mining the microbiota to identify gut commensals modulating neuroinflammation in a mouse model of multiple sclerosis
Microbiome (2022)
-
TGF-β production by eosinophils drives the expansion of peripherally induced neuropilin− RORγt+ regulatory T-cells during bacterial and allergen challenge
Mucosal Immunology (2022)
-
Dendritic cell functions in the inductive and effector sites of intestinal immunity
Mucosal Immunology (2022)
-
The microbiota is dispensable for the early stages of peripheral regulatory T cell induction within mesenteric lymph nodes
Cellular & Molecular Immunology (2021)

{kind=link}
{kind=link}
{kind=link}