
Abstract
Background:
Cdc20 is an essential component of cell division and responsible for anaphase initiation regulated by securin degradation. Cdc20 function is strongly regulated by the spindle assembly checkpoint to ensure the timely separation of sister chromatids and integrity of the genome. We present the first results on Cdc20 in a large clinical breast cancer material.
Methods:
The study was based on 445 breast cancer patients with up to 20 years of follow-up (mean 10.0 years). DNA content was determined by image cytometry on cell imprints, and Cdc20 and securin immunohistochemistry on tissue microarrays of breast cancer tissue.
Results:
In our results, high Cdc20 and securin expression was associated with aneuploid DNA content. In prognostic analyses, high Cdc20 immunoexpression alone and in combination with high securin immunoexpression indicated aggressive course of disease and up to 6.8-fold (P<0.001) risk of breast cancer death. Particularly, high Cdc20 and securin immunoexpression identified a patient subgroup with extremely short, on average 2.4 years, breast cancer survival and triple-negative breast cancer (TNBC) subtype.
Conclusions:
We report for the first time the association of high Cdc20 and securin immunoexpression with extremely poor outcome of breast cancer patients. Our experience indicates that Cdc20 and securin are promising candidates for clinical applications in breast cancer prognostication, especially in the challenging prognostic decisions of TNBC.
Similar content being viewed by others

Main
According to present knowledge, the most essential steps in cell division occur in metaphase–anaphase transition during chromosome segregation (Yamanaka et al, 2012). Accuracy of chromosome segregation is dependent on mitotic spindle assembly checkpoint (SAC), which delays anaphase initiation until all sister chromatids are correctly attached to the mitotic spindle by their kinetochores (Musacchio and Salmon, 2007; Kim and Yu, 2011). Inappropriately functioning SAC has been linked to chromosomal instability (CIN) and aneuploidy (Jallepalli and Lengauer, 2001; Suijkerbuijk and Kops, 2008). The key event of SAC is inhibition of anaphase-promoting complex/cyclosome (APC/C), a ubiquitin ligase that targets degradation of securin and cyclin B1 when activated by Cdc20. Activation of the APC–Cdc20 complex and degradation of securin and cyclin B1 then mark anaphase onset in normally regulated cell division. The checkpoint proteins, such as Mad1, Mad2, BubR1, Bub1, Bub3, Mps1 and AuroraB form a complex signalling network that inhibits APC/C-Cdc20 (Musacchio and Salmon, 2007; Kim and Yu, 2011). Mad2 and BubR1 function in APC/C inhibition both independently and as a part of a more potent inhibitory complex with Bub3 and Cdc20, the so called mitotic checkpoint complex (MCC) (Sudakin et al, 2001; Tang et al, 2001; Fang, 2002).
Human Cdc20/p55cdc, a homologue of Saccharomyces cerevisiae cell division cycle 20 protein, is one of the most important components of the mammalian cell-cycle mechanism (Weinstein et al, 1994; Weinstein, 1997). As an integral part of the SAC, Cdc20 ensures that anaphase proceeds only when the centromeres of all sister chromatids are lined up in the metaphase plate and properly attached to microtubules. Recently Cdc20 overexpression has been associated with inappropriately functioning SAC and aneuploidization in oral cancer (Mondal et al, 2007). High Cdc20 expression has been reported in several human cancer cell lines (Kim et al, 2005b; Iacomino et al, 2006; Thirthagiri et al, 2007; Yuan et al, 2006; Jiang et al, 2011; Chang et al, 2012) and several carcinoma tissues (Kim et al, 2005b; Ouellet et al, 2006; Kidokoro et al, 2008; Jiang et al, 2011). High Cdc20 expression has also been linked to poor prognosis in lung (Kato et al, 2012), oral squamous cell (Moura et al, 2013), bladder (Choi et al, 2013), colon (Wu et al, 2013) and pancreatic (Chang et al, 2012) carcinomas.
Overexpression of securin has been associated with unfavourable prognosis in multiple cancer types (Vlotides et al, 2007; Salehi et al, 2008). In our previous work, we have shown that high securin expression predicts aneuploidy and unfavourable prognosis in human breast cancer (Karra et al, 2012). Active securin binds to and inhibits separase activity, whereas degradation of securin releases separase to cleave centromeric cohesion at anaphase onset – a process dependent on APC-Cdc20 activation (Nasmyth, 1999; Nasmyth et al, 2000). Thus, securin seems to have an anaphase promoting quality, as the prior interaction with securin is needed for separase to function properly (Kumada et al, 1998; Jensen et al, 2001). Both overexpression and lack of securin have been suggested to compromise chromosomal stability (Jallepalli et al, 2001; Bernal et al, 2002; Yu et al, 2003; Kim et al, 2005a, 2007). Recently a single mutation in securin was shown to induce CIN (Mora-Santos et al, 2013). The complete role of securin in diseased cells is still not settled but multifactorial mechanisms in aneuploidy, apoptosis, angiogenesis, tumour cell transformation and microenvironment regulation and role in DNA repair checkpoint have been suggested (Tfelt-Hansen et al, 2006; Vlotides et al, 2007; Salehi et al, 2008). Increasing evidence suggests clinically significant applications of securin in association with disease outcome and as a biomarker for subsequent therapeutic interventions (Solbach et al, 2005; Cho-Rok et al, 2006; Kakar and Malik, 2006; Panguluri et al, 2008).
The purpose of this study is to evaluate the significance of Cdc20 independently and in combination with securin as a prognostic marker for breast cancer patients. The study is based on a material of 445 breast cancer patients with up to 22 years of follow-up (mean follow-up 10.0 years). Our findings suggest that the combination of high Cdc20 and securin expression identify a specific group of patients with aneuploid cell type and extremely short breast cancer survival, in average 2.4 years after diagnosis (median 2.7 years). In our material, Cdc20 and securin overexpression was associated with aggressive cell morphology and triple-negative breast carcinoma subtype.
Materials and methods
Patients
The patient material includes a total of 445 women with unilateral invasive breast cancer diagnosed and treated in Jyväskylä Central Hospital, Jyväskylä, Finland in 1987–1997 during the era of the national mammography screening programme of Central Finland. All patients were treated with surgical resection or mastectomy with axillary evacuation, radiation and/or adjuvant treatment with anti-estrogenic or cytostatic drugs depending on the patients’ age, hormone receptor and lymph node status according to the international guidelines for breast cancer treatment at the time of diagnosis (Goldhirsh et al, 2009). No pre-operative adjuvant treatments were administered. Complete clinical follow-up information was available from pathology reports and patient files. The clinico-pathological data of the material (Table 1) was collected according to the criteria presented by WHO (Lakhani and International Agency for Research on Cancer, World Health Organization, 2012). In addition, intrinsic breast cancer classification for the genetically identified breast cancer subtypes (Perou et al, 2000; Sorlie et al, 2001; Sotiriou et al, 2003) was performed as approximations recommended by the 12th St Gallen International Breast Cancer Conference Expert Panel (Hammond et al, 2010; Goldhirsch et al, 2011). The maximum follow-up period of the patient material was 22 years and 6 months (mean 10.0 years). Causes of death were obtained from autopsy reports, death certificates and from the Finnish Cancer Registry. The Ethical Committees of Jyväskylä Central Hospital and Turku University Hospital have approved the patient material and study setting.
Image cytometry analysis
For image cytometric analysis, material from tumours of 229 patients was collected as previously described by Karra et al, 2012. Briefly, cell imprints were prepared by applying a freshly cut surface of the tumour on glass slide. The histological diagnosis of all cell imprints was morphologically verified from routine formalin-fixed (pH 7.0), paraffin-embedded and H&E-stained tissue specimen from the same cut surface of the tumour. The imprints were stained according to Feulgen (CAS DNA Staining Kit, Becton Dickinson Cellular Imaging Systems, Elmhurst, IL, USA) and analysed by determining the DNA content of each tumour quantitatively as the intensity of nuclear staining in both automatic and visual evaluations in light microscopy (CAS 200 Image Analysis System, Cell Analysis Systems, Elmhurst, IL, USA). Calibration and biological reference of the image analysis system was based on internal control of diploid cells and external control of rat tetraploid hepatocytes.
DNA histograms were classified according to visual interpretation. Cases were allocated into diploid (⩾90% of cells forming a single peak between 1,6c and 2,4c), tetraploid (a minimum of 10% of cells forming a peak between 3,6c and 4,4c) or aneuploid (>10% of cells outside diploid and tetraploid ranges). Moreover, particularly strongly aneuploid cases were further described on the basis of observed fraction of cancer cells with DNA content exceeding 5c (5c exceeding rate, 5cER) and 16c (16c exceeding rate, 16cER) (Karra et al, 2012).
Immunohistochemistry and in situ hybridisation
Tissue material for Cdc20 and securin staining was obtained from tumours of all 445 non-consequtive breast cancer cases. For immunohistochemistry, tissue microarrays (TMA) were prepared by selecting a representative block of each patient and punching the tumour area for two tissue cores 0.6 mm in diameter. After arranging the cores precisely to TMAs, each of the six blocks contained 128–312 tissue cores.
Cdc20, oestrogen receptor (ER), progesterone receptor (PR), Ki-67 and Her2 immunostainings were performed with the automated immunostaining machine BenchMark XT (Roche Diagnostics/Ventana Medical Systems, Tucson, AZ, USA) using ultraView Universal DAB Detection Kit (Roche/Ventana). Securin immunostaining was performed with LabVision Autostainer 480 (Thermo-Fisher Scientific, Fremont, CA, USA) and detected with Power Vision+polymer kit, according to standard protocol (DPVB+110HRP; Immunovision Technologies, Vision Biosystems, Norwell, MA, USA), with diaminobenzidine as chromogen. Details for antibodies, antigen retrieval and staining methods are summarised in Table 2.
Interpretation of Cdc20, securin and Ki-67 positivity was performed on sets of 100 cancer cells (minimum of one set and maximum three sets of 100 cells evaluated in each tissue core). Tissue cores showing <100 invasive cells were excluded from the study. Cdc20 immunopositivity was observed in the cytoplasm of cancer cells. The cases (4.3%) with strong diffuse staining were clearly identifiable among all tissue cores. In addition, a fraction of cases showed weak diffuse staining (23% of cases) or staining in single cells (12% of cases). The majority of cases (61%) were classified negative on the basis of <5% of positive cancer cells observed. Securin immunopositivity was both nuclear and cytoplasmic and registered as the fraction (%) of positive cancer cells (median 7.5%, range 0–84.5%, 50% of cases between 4.0%–15.3%) as described by Karra et al, 2012. Interpretation of ER, PR and Ki-67 immunohistochemistry was performed according to generally accepted international guidelines (Hammond et al, 2010).
Cases for Her2-amplification testing were selected on the basis of immunohistochemistry, and gene amplification was thereafter confirmed with in situ hybridisation. Her2/Chr17 double in situ hybridisation was performed with the BenchMark XT (Roche/Ventana) using Ventana Her2 DNA probe (Roche/Ventana), Inform Chromosome 17 probe (Roche/Ventana), ultraView SISH detection kit to detect Her2 (Roche/Ventana) and ultraView Alkaline Prosphatase Red ISH Detection Kit to detect Chr17 (Roche/Ventana). Briefly, ISH Protease 3 (Roche/Ventana) for 8 min was used as a pretreatment step, and Her2 hybridisation was performed at 52°C for 6 h and Chr17 hybridisation at 44°C for 2 h. Interpretation of both immunohistochemistry and in situ hybridisation was performed according to generally accepted international guidelines (Wolff et al, 2007).
Statistical analysis
For statistical analyses, patients were allocated into low-level and high-level expression groups based on Cdc20 and securin immunopositivity and cutoff points based on reported prognostic associations and mean values in the patient material (Karra et al, 2012). Survival analyses for Cdc20 and securin were performed as Kaplan–Meier estimates to demonstrate cumulative percentages of breast cancer-specific mortality, and Cox’s proportional hazards models to assess differences between the groups of patients with different prognoses. Cox regression analysis was performed to characterise prognostic associations adjusted for the established prognostic features: tumour size, axillary lymph node status, histological and intrinsic cancer type and histological grade. Relation between the proteins under study and the established clinical prognosticators were quantified as hazard ratios (HRs) with 95% confidence intervals (CI). Associations between Cdc20 and securin immunopositivity, features of DNA content and the established prognosticators of breast cancer were analysed by Fisher’s exact and Wilcoxon rank sum tests, and the results were quantified as odds ratios with 95% CIs. P-values <0.05 were considered statistically significant. The validity of proportional hazards assumption was assessed both visually and numerically, and no marked deviation for assumptions were observed. Kaplan–Meier survival plots were generated using R 2.15.0. All computations were performed with SAS for Windows, Version 9.3 (SAS Institute, Cary, NC, USA).
Results
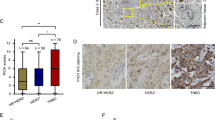
Photomicrographs in Figure 1 show the immunostaining patterns of Cdc20 and securin in invasive breast cancer. Cdc20 immunopositivity was allocated into four expression groups: negative in >95% of cancer cells (score 0), positive in >5% of single cancer cells (score 1+), weak diffuse staining (score 2+) and strong diffuse staining in >95% of cancer cells (score 3+). The cases classified as score 3+ (Figure 1D) were a small (n=19, 4.3% of the whole material) but distinct patient group clearly distinguished from the other cases based on the strong Cdc20 immunoexpression and extremely poor prognosis (P<0.001) (Figure 2). In further analyses, this high expression group was evaluated separately from the rest of the cases (low-expression group). In securin stainings the average fraction of immunopositive cancer cells was 10.2% (median 7.5%, range 0–84.5%, 50% of cases between 4.0–15.3%). For statistical analyses, the threshold at 10% immunopositivity was used for dividing the material in low and high expression groups (low 62.9% and high 37.1% of cases) (Karra et al, 2012).

Examples of high and low immunostaining patterns for Cdc20 and securin in human breast cancer including both positive and negative staining mitotic figures.

Kaplan–Meier curves showing breast cancer-specific patient survival in the different Cdc20 expression groups ( P <0.001). Cdc20 immunopositivity is expressed as score 0 (negative in >95% of cancer cells), score 1+ (positive in >5% of single cancer cells), score 2+ (weak diffuse staining in >95% of cancer cells) and score 3+ (strong diffuse staining in >95% of cancer cells).
In DNA cytometric analyses, cases with high and low Cdc20 expression showed aneuploid cell type in 62.5% and 35.6% of cases, respectively. Table 3 summarises the associations for Cdc20 expression and ploidy. In statistical analyses, the association between high Cdc20 and ploidy sparsely failed to show statistical significance (P=0.059), probably because of the low number of cases in this group. In our material, all cases showing the combination of high Cdc20 and securin expression had abnormal DNA content. High expression of both proteins predicted 19-fold odds (P=0.004, CI 1.1–344.8) for aneuploid DNA content as compared with cases with low expression in both stainings. In addition, high Cdc20 expression in combination with high securin expression predicted the occurrence of 5cER cells (OR 17.0, CI 2.1–135.1).
In prognostic analysis involving all patients and the entire follow-up period of maximum 22 years (mean 10.0 years), high Cdc20 immunoexpression predicted twofold risk of breast cancer death (P=0.047, CI 1.0–3.9). We have earlier reported a strong association between securin immunoexpression and poor prognosis of breast cancer (Karra et al, 2012). When the established prognosticators of breast cancer were added into the multivariate analysis with securin and Cdc20, high securin expression was shown as statistically significant prognosticator along with axillary lymph node status, tumour size and histological grade (grade 1 vs 2) (Table 4). Table 5 summarises the associations between Cdc20 and securin immunoexpression, DNA ploidy, and the established prognosticators of human breast cancer. Despite the observed unfavourable prognosis associated with high Cdc20 expression, the majority of the patients were axillary lymph node negative (63%). This association was, however, not statistically significant, probably owing to small number of patients in the subgroup.
Figure 3A shows the Kaplan–Meier curves of the patient material divided according to Cdc20 immunoexpression (P=0.047). The prognostic implications were still intensified when analysing Cdc20 and securin immunoexpression in combination (Figure 3B). In our material, this analysis indicated 4.3-fold risk of breast cancer death (P<0.001, CI 2.0–8.9) for patients with high Cdc20 and high securin expression as related to low Cdc20 and low securin expression. Detailed quartile estimations of Kaplan–Meier analysis for individual patients suggested that the majority (75%) of patients with low Cdc20 expression were alive 7.9 years after diagnosis, whereas the majority (75%) of patients with high Cdc20 expression were alive only 1.9 years after diagnosis. In the same vein, the majority (75%) of patients with the most favourable combination of Cdc20 and securin (low expression for both) looked forward to 13.3 years of survival. Instead, the majority (75%) of patients with the most unfavourable combination of Cdc20 and securin (high expression for both) could expect only 1.3-year breast cancer survival.

Disease-specific survival determined according to Cdc20 and securin immunoexpression in 445 breast cancer patients after the whole follow-up period (maximum 22 years, mean 10.0) and after the first 5 years after primary diagnosis. (A) shows Kaplan–Meier estimates of patient groups with low (scores 0, 1+ and 2+) and high (score 3+) Cdc20 immunoexpression. (B) presents Kaplan–Meier survival curves of patient material divided according to the combination of cdc20 and securin immunopositivity (curve A: low Cdc20 and low securin, curve B: high Cdc20 and low securin expression, and curve C: high Cdc20 and high securin expression). (C) shows in detail the survival after the first 5 years after diagnosis. The subgroup with high Cdc20 and low securin expression is not presented owing to the small amount of cases (n=4).
The prognostic value of the combination of Cdc20 and securin immunoexpression was particularly strong in Kaplan–Meier analysis modelling short-term survival (up to 5 years of diagnosis) (Figure 3C). In this setting, patients showing high expression for both Cdc20 and securin were associated with 6.8-fold risk of breast cancer death (P<0.001, CI 3.2–14.9) as compared with patients with low Cdc20 and securin expression. When the established prognosticators of breast cancer were included in the multivariate analysis, only securin, nodal status and tumour size showed statistically significant prognostic associations at 5 years of follow-up. In addition to the extremely aggressive course of disease, the small subgroup (n=19) of patients with high expression for both Cdc20 and securin stood out histologically on the basis of their particularly atypical morphology with strong pleomorphism, several multinucleated cancer cells and high mitotic activity with frequent pathological mitoses (Figure 4). Analysis of this subgroup with the help of the established prognosticators showed that all but one of the cases represented the triple-negative ‘basal-like’ cell type in the surrogate genetic classification of St Gallen International Expert consensus (Goldhirsch et al, 2011). The remaining one case showed hormone receptor positivity but was Her2-oncogen negative. Based on this very small patient group, the combined high expression of Cdc20 and securin predicted 88-fold odds for triple-negative breast cancer (TNBC) subtype (P<0.001, CI 11.3–688.3).

Two examples showing the morphology of human breast cancer with high Cdc20 and high securin. Cancer cells exhibit strong cell atypia with pleomorphic and multinucleated cancer cells and high mitotic activity with frequent pathological mitoses.
Discussion
We report for the first time on the prognostic associations of the essential cell-cycle regulator, Cdc20, in human breast cancer. As an activating subunit of APC/C, Cdc20 is known to drive mitosis from metaphase to anaphase, where the APC/C-Cdc20-mediated degradation of securin is of critical importance (Peters, 2006). The role of Cdc20 in sister chromatid separation has previously been demonstrated in Cdc20 depleted mice whose embryos are arrested in metaphase at the two-cell stage with high levels of cyclin B1 and securin (Li et al, 2007). In a further experiment with Cdc20 and securin double mutant embryo, metaphase was not arrested, but the loss of securin could not rescue the embryos form Cdc20 deficiency-induced lethality (Li et al, 2007).
Experiments with mice have suggested that mutant, SAC-inhibition resistant Cdc20 promotes tumorigenesis (Li et al, 2009). Vice versa, knockdown of Cdc20 expression has resulted in growth suppression of tumour cells (Kidokoro et al, 2008; Taniguchi et al, 2008). Increase in Cdc20 expression has been reported in many human cancers, often with associated less favourable prognosis (Chang et al, 2012; Choi et al, 2013; Kato et al, 2012; Moura et al, 2013; Wu et al, 2013). In cDNA microarray analyses based on human cancer tissues, including breast cancer, Kidokoro et al, (2008) found Cdc20 to be overexpressed in a large set of human malignancies. The Cdc20 expression was increased more than threefolds in 44% of all cancer tissues examined and in 60% of the breast cancer tissues (Kidokoro et al, 2008).
Previously, Nilsson (2011) has suggested that the level of Cdc20 might be one of the key features determining response to anti-mitotic cancer therapeutics. Downregulation of Cdc20 in breast cancer cells has been associated with inhibition of cell proliferation in vitro (Jiang et al, 2011, 2012). Furthermore, treatment with siRNA against Cdc20 has been shown to induce G2/M arrest and suppress cell growth (Kidokoro et al, 2008; Taniguchi et al, 2008).
Consistent with their role in regulation of cell proliferation, we found that both Cdc20 and securin were associated with Ki-67 labelling index and histological grade, which includes the feature of mitotic activity. In the present statistical analyses, securin and histological grade were independent prognosticators of invasive breast cancer. Previously, securin has been reported to intensify the prognostic power of Ki-67 alone by identifying a specific subgroup of patients with more favourable outcome than indicated by Ki-67 alone (Talvinen et al, 2008, 2009). In our observations from immunohistochemistry, only part of the mitotic figures in breast cancer tissue stain positive for Cdc20 or securin (Figures 1E and F). In light of these morphological and statistical findings, Cdc20 and securin seem to be potential prognosticators for clinical pathology, although the present data does not yet allow testing for their prognostic value in relation to the gold standard of breast cancer prognostication, the Nottingham Prognostic Index (Elston and Ellis, 1991).
The prognostic analyses based on a long-term follow-up indicated substantially increased risk of breast cancer death for patients with high Cdc20 immunoexpression alone (HR 2.0, P=0.047) and in combination with high securin immunoexpression (HR 4.3, P<0.001). These findings are emphasised by the long-term follow-up of the patient material. Still, the most impressive prognostic associations were obtained when focusing on the first 5 years after diagnosis. In our material, patients with high Cdc20 and securin immunoexpression had 6.8-fold risk of breast cancer death within 5 years from diagnosis. The results suggest that the combination of high Cdc20 and securin expression indicates biologically aggressive disease and particularly sinister outcome and could, therefore, be applicable in identifying patients who would benefit from the most effective oncological treatments. In our material, the combination of Cdc20 and securin expression resulted in an impressive survival difference between the groups of most favourable and most unfavourable prognosis. After the whole follow-up period, the fraction of patients dead of breast cancer was 62% for patients with high expression for Cdc20 and securin, and 24% for patients with low expression for Cdc20 and securin.
Currently, the main clinical problem is the lack of targeted therapies for TNBC (O’Toole et al, 2013). Of special interest in our results is, therefore, the observation that TNBCs were strongly overrepresented among cases with high Cdc20 and securin expression (94% of cases). Unfortunately, the patient subgroup is too small (n=19) and the statistical associations are too weak (CI 11.3–688.3) for any definitive conclusions. Still, the findings seem to indicate, for subgroups divided according to Cdc20 and securin expression, a considerable survival difference, which is worth further investigations. TNBCs have been used as a surrogate for the particularly aggressive subtype known as ‘basal-like’ cancers, which are most commonly diagnosed in younger age, in association with BRCA1 mutation and with a distinct pattern of progression and metastasis (Dent et al, 2007; Anders and Carey, 2008; Billar et al, 2010; Foulkes et al, 2010). We observed immunopositivity for basal cytokeratins in all but one of the cases with high Cdc20 and securin expression. According to literature, TNBC is not a single subtype of breast cancer but comprises a heterogeneous group of malignancies with distinct molecular signature and therapeutic responses (Peddi et al, 2012; Chiorean et al, 2013; Stagg and Allard, 2013). Lehmann et al (2011) have recently reported that part of TNBCs identified on the basis of pathways related to cell-cycle control (Basal-like 1 cancer type) may identify a patient subgroup with extremely poor prognosis.
We report for the first time on the prognostic association of Cdc20 and securin expression in breast cancer patients. In our material, high expression of both proteins predicted strong probability (19-fold odds, P=0.004) for aneuploid DNA content as compared with low expression cases. In prognostic analysis, high Cdc20 immunoexpression alone predicted twofold risk of breast cancer death (P=0.047), whereas the combination of high Cdc20 and high securin indicated 4.3-fold risk of breast cancer death (P<0.001). In multivariate analyses, high securin expression was shown as a statistically significant prognosticator along with axillary lymph node status, tumour size and histological grade. Our findings suggest that Cdc20 and securin are potential candidates for clinical applications in breast cancer prognostication.
Change history
10 June 2014
This paper was modified 12 months after initial publication to switch to Creative Commons licence terms, as noted at publication
References
Anders C, Carey LA (2008) Understanding and treating triple-negative breast cancer. Oncology (Williston Park) 22: 1233–1239.
Bernal JA, Luna R, Espina A, Lazaro I, Ramos-Morales F, Romero F, Arias C, Silva A, Tortolero M, Pintor-Toro JA (2002) Human securin interacts with p53 and modulates p53-mediated transcriptional activity and apoptosis. Nat Genet 32: 306–311.
Billar JA, Dueck AC, Stucky CC, Gray RJ, Wasif N, Northfelt DW, McCullough AE, Pockaj BA (2010) Triple-negative breast cancers: unique clinical presentations and outcomes. Ann Surg Oncol 17 (Suppl 3): 384–390.
Chang DZ, Ma Y, Ji B, Liu Y, Hwu P, Abbruzzese JL, Logsdon C, Wang H (2012) Increased CDC20 expression is associated with pancreatic ductal adenocarcinoma differentiation and progression. J Hematol Oncol 5: 15–8722-5-15.
Chiorean R, Braicu C, Berindan-Neagoe I (2013) Another review on triple negative breast cancer. Are we on the right way towards the exit from the labyrinth? Breast 22: 1026–1033.
Cho-Rok J, Yoo J, Jang YJ, Kim S, Chu IS, Yeom YI, Choi JY, Im DS (2006) Adenovirus-mediated transfer of siRNA against PTTG1 inhibits liver cancer cell growth in vitro and in vivo. Hepatology 43: 1042–1052.
Choi JW, Kim Y, Lee JH, Kim YS (2013) High expression of spindle assembly checkpoint proteins CDC20 and MAD2 is associated with poor prognosis in urothelial bladder cancer. Virchows Arch 463: 681–687.
Dent R, Trudeau M, Pritchard KI, Hanna WM, Kahn HK, Sawka CA, Lickley LA, Rawlinson E, Sun P, Narod SA (2007) Triple-negative breast cancer: clinical features and patterns of recurrence. Clin Cancer Res 13: 4429–4434.
Elston CW, Ellis IO (1991) Pathological prognostic factors in breast cancer. I. The value of histological grade in breast cancer: experience from a large study with long-term follow-up. Histopathology 19: 403–410.
Fang G (2002) Checkpoint protein BubR1 acts synergistically with Mad2 to inhibit anaphase-promoting complex. Mol Biol Cell 13: 755–766.
Foulkes WD, Smith IE, Reis-Filho JS (2010) Triple-negative breast cancer. N Engl J Med 363: 1938–1948.
Goldhirsch A, Ingle JN, Gelber RD, Coates AS, Thürlimann B, Senn HJ Panel members (2009) Thresholds for therapies: highlights of the St Gallen International Expert Consensus on the primary therapy of early breast cancer (2009) Ann Oncol 20: 1319–1329.
Goldhirsch A, Wood WC, Coates AS, Gelber RD, Thurlimann B, Senn HJ Panel members (2011) Strategies for subtypes—dealing with the diversity of breast cancer: highlights of the St. Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2011. Ann Oncol 22: 1736–1747.
Hammond ME, Hayes DF, Dowsett M, Allred DC, Hagerty KL, Badve S, Fitzgibbons PL, Francis G, Goldstein NS, Hayes M, Hicks DG, Lester S, Love R, Mangu PB, McShane L, Miller K, Osborne CK, Paik S, Perlmutter J, Rhodes A, Sasano H, Schwartz JN, Sweep FC, Taube S, Torlakovic EE, Valenstein P, Viale G, Visscher D, Wheeler T, Williams RB, Wittliff JL, Wolff AC (2010) American Society of Clinical Oncology/College Of American Pathologists guideline recommendations for immunohistochemical testing of estrogen and progesterone receptors in breast cancer. J Clin Oncol 28: 2784–2795.
Iacomino G, Medici MC, Napoli D, Russo GL (2006) Effects of histone deacetylase inhibitors on p55CDC/Cdc20 expression in HT29 cell line. J Cell Biochem 99: 1122–1131.
Jallepalli PV, Lengauer C (2001) Chromosome segregation and cancer: cutting through the mystery. Nat Rev Cancer 1: 109–117.
Jallepalli PV, Waizenegger IC, Bunz F, Langer S, Speicher MR, Peters JM, Kinzler KW, Vogelstein B, Lengauer C (2001) Securin is required for chromosomal stability in human cells. Cell 105: 445–457.
Jensen S, Segal M, Clarke DJ, Reed SI (2001) A novel role of the budding yeast separin Esp1 in anaphase spindle elongation: evidence that proper spindle association of Esp1 is regulated by Pds1. J Cell Biol 152: 27–40.
Jiang J, Jedinak A, Sliva D (2011) Ganodermanontriol (GDNT) exerts its effect on growth and invasiveness of breast cancer cells through the down-regulation of CDC20 and uPA. Biochem Biophys Res Commun 415: 325–329.
Jiang J, Thyagarajan-Sahu A, Krchnak V, Jedinak A, Sandusky GE, Sliva D (2012) NAHA, a novel hydroxamic acid-derivative, inhibits growth and angiogenesis of breast cancer in vitro and in vivo. PLoS One 7: e34283.
Kakar SS, Malik MT (2006) Suppression of lung cancer with siRNA targeting PTTG. Int J Oncol 29: 387–395.
Karra H, Pitkanen R, Nykanen M, Talvinen K, Kuopio T, Soderstrom M, Kronqvist P (2012) Securin predicts aneuploidy and survival in breast cancer. Histopathology 60: 586–596.
Kato T, Daigo Y, Aragaki M, Ishikawa K, Sato M, Kaji M (2012) Overexpression of CDC20 predicts poor prognosis in primary non-small cell lung cancer patients. J Surg Oncol 106: 423–430.
Kidokoro T, Tanikawa C, Furukawa Y, Katagiri T, Nakamura Y, Matsuda K (2008) CDC20, a potential cancer therapeutic target, is negatively regulated by p53. Oncogene 27: 1562–1571.
Kim D, Pemberton H, Stratford AL, Buelaert K, Watkinson JC, Lopes V, Franklyn JA, McCabe CJ (2005a) Pituitary tumour transforming gene (PTTG) induces genetic instability in thyroid cells. Oncogene 24: 4861–4866.
Kim DS, Franklyn JA, Smith VE, Stratford AL, Pemberton HN, Warfield A, Watkinson JC, Ishmail T, Wakelam MJ, McCabe CJ (2007) Securin induces genetic instability in colorectal cancer by inhibiting double-stranded DNA repair activity. Carcinogenesis 28: 749–759.
Kim JM, Sohn HY, Yoon SY, Oh JH, Yang JO, Kim JH, Song KS, Rho SM, Yoo HS, Kim YS, Kim JG, Kim NS (2005b) Identification of gastric cancer-related genes using a cDNA microarray containing novel expressed sequence tags expressed in gastric cancer cells. Clin Cancer Res 11: 473–482.
Kim S, Yu H (2011) Mutual regulation between the spindle checkpoint and APC/C. Semin Cell Dev Biol 22: 551–558.
Kumada K, Nakamura T, Nagao K, Funabiki H, Nakagawa T, Yanagida M (1998) Cut1 is loaded onto the spindle by binding to Cut2 and promotes anaphase spindle movement upon Cut2 proteolysis. Curr Biol 8: 633–641.
Lakhani SR International Agency for Research on Cancer, World Health Organization (2012) WHO Classification of Tumours of the Breast. International Agency for Research on Cancer: Lyon.
Lehmann BD, Bauer JA, Chen X, Sanders ME, Chakravarthy AB, Shyr Y, Pietenpol JA (2011) Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J Clin Invest 121: 2750–2767.
Li M, Fang X, Wei Z, York JP, Zhang P (2009) Loss of spindle assembly checkpoint-mediated inhibition of Cdc20 promotes tumorigenesis in mice. J Cell Biol 185: 983–994.
Li M, York JP, Zhang P (2007) Loss of Cdc20 causes a securin-dependent metaphase arrest in two-cell mouse embryos. Mol Cell Biol 27: 3481–3488.
Mondal G, Sengupta S, Panda CK, Gollin SM, Saunders WS, Roychoudhury S (2007) Overexpression of Cdc20 leads to impairment of the spindle assembly checkpoint and aneuploidization in oral cancer. Carcinogenesis 28: 81–92.
Mora-Santos M, Castilla C, Herrero-Ruiz J, Giraldez S, Limon-Mortes MC, Saez C, Japon MA, Tortolero M, Romero F (2013) A single mutation in Securin induces chromosomal instability and enhances cell invasion. Eur J Cancer 49: 500–510.
Moura IM, Delgado ML, Silva PM, Lopes CA, do Amaral JB, Monteiro LS, Bousbaa H (2013) High CDC20 expression is associated with poor prognosis in oral squamous cell carcinoma. J Oral Pathol Med 43: 225–231.
Musacchio A, Salmon ED (2007) The spindle-assembly checkpoint in space and time. Nat Rev Mol Cell Biol 8: 379–393.
Nasmyth K (1999) Separating sister chromatids. Trends Biochem Sci 24: 98–104.
Nasmyth K, Peters JM, Uhlmann F (2000) Splitting the chromosome: cutting the ties that bind sister chromatids. Science 288: 1379–1385.
Nilsson J (2011) Cdc20 control of cell fate during prolonged mitotic arrest: do Cdc20 protein levels affect cell fate in response to antimitotic compounds? Bioessays 33: 903–909.
O'Toole SA, Beith JM, Millar EK, West R, McLean A, Cazet A, Swarbrick A, Oakes SR (2013) Therapeutic targets in triple negative breast cancer. J Clin Pathol 66: 530–542.
Ouellet V, Guyot MC, Le Page C, Filali-Mouhim A, Lussier C, Tonin PN, Provencher DM, Mes-Masson AM (2006) Tissue array analysis of expression microarray candidates identifies markers associated with tumor grade and outcome in serous epithelial ovarian cancer. Int J Cancer 119: 599–607.
Panguluri SK, Yeakel C, Kakar SS (2008) PTTG: an important target gene for ovarian cancer therapy. J Ovarian Res 1: 6.
Peddi PF, Ellis MJ, Ma C (2012) Molecular basis of triple negative breast cancer and implications for therapy. Int J Breast Cancer 2012: 217185.
Perou CM, Sorlie T, Eisen MB, van de Rijn M, Jeffrey SS, Rees CA, Pollack JR, Ross DT, Johnsen H, Akslen LA, Fluge O, Pergamenschikov A, Williams C, Zhu SX, Lonning PE, Borresen-Dale AL, Brown PO, Botstein D (2000) Molecular portraits of human breast tumours. Nature 406: 747–752.
Peters JM (2006) The anaphase promoting complex/cyclosome: a machine designed to destroy. Nat Rev Mol Cell Biol 7: 644–656.
Salehi F, Kovacs K, Scheithauer BW, Lloyd RV, Cusimano M (2008) Pituitary tumor-transforming gene in endocrine and other neoplasms: a review and update. Endocr Relat Cancer 15: 721–743.
Solbach C, Roller M, Peters S, Nicoletti M, Kaufmann M, Knecht R (2005) Pituitary tumor-transforming gene (PTTG): a novel target for anti-tumor therapy. Anticancer Res 25: 121–125.
Sorlie T, Perou CM, Tibshirani R, Aas T, Geisler S, Johnsen H, Hastie T, Eisen MB, van de Rijn M, Jeffrey SS, Thorsen T, Quist H, Matese JC, Brown PO, Botstein D, Lonning PE, Borresen-Dale AL (2001) Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc Natl Acad Sci USA 98: 10869–10874.
Sotiriou C, Neo SY, McShane LM, Korn EL, Long PM, Jazaeri A, Martiat P, Fox SB, Harris AL, Liu ET (2003) Breast cancer classification and prognosis based on gene expression profiles from a population-based study. Proc Natl Acad Sci USA 100: 10393–10398.
Stagg J, Allard B (2013) Immunotherapeutic approaches in triple-negative breast cancer: latest research and clinical prospects. Ther Adv Med Oncol 5: 169–181.
Sudakin V, Chan GK, Yen TJ (2001) Checkpoint inhibition of the APC/C in HeLa cells is mediated by a complex of BUBR1, BUB3, CDC20, and MAD2. J Cell Biol 154: 925–936.
Suijkerbuijk SJ, Kops GJ (2008) Preventing aneuploidy: the contribution of mitotic checkpoint proteins. Biochim Biophys Acta 1786: 24–31.
Talvinen K, Tuikkala J, Nevalainen O, Rantanen A, Hirsimäki P, Sundström J, Kronqvist P (2008) Proliferation marker securin identifies favourable outcome in invasive ductal breast cancer. Br J Cancer 99: 335–340.
Talvinen K, Karra H, Hurme S, Nykänen M, Nieminen A, Anttinen J, Kuopio T, Kronqvist P (2009) Securin promotes the identification of favourable outcome in invasive breast cancer. Br J Cancer 101: 1005–1010.
Tang Z, Bharadwaj R, Li B, Yu H (2001) Mad2-Independent inhibition of APCCdc20 by the mitotic checkpoint protein BubR1. Dev Cell 1: 227–237.
Taniguchi K, Momiyama N, Ueda M, Matsuyama R, Mori R, Fujii Y, Ichikawa Y, Endo I, Togo S, Shimada H (2008) Targeting of CDC20 via small interfering RNA causes enhancement of the cytotoxicity of chemoradiation. Anticancer Res 28: 1559–1563.
Tfelt-Hansen J, Kanuparthi D, Chattopadhyay N (2006) The emerging role of pituitary tumor transforming gene in tumorigenesis. Clin Med Res 4: 130–137.
Thirthagiri E, Robinson CM, Huntley S, Davies M, Yap LF, Prime SS, Paterson IC (2007) Spindle assembly checkpoint and centrosome abnormalities in oral cancer. Cancer Lett 258: 276–285.
Vlotides G, Eigler T, Melmed S (2007) Pituitary tumor-transforming gene: physiology and implications for tumorigenesis. Endocr Rev 28: 165–186.
Weinstein J (1997) Cell cycle-regulated expression, phosphorylation, and degradation of p55Cdc. A mammalian homolog of CDC20/Fizzy/slp1. J Biol Chem 272: 28501–28511.
Weinstein J, Jacobsen FW, Hsu-Chen J, Wu T, Baum LG (1994) A novel mammalian protein, p55CDC, present in dividing cells is associated with protein kinase activity and has homology to the Saccharomyces cerevisiae cell division cycle proteins Cdc20 and Cdc4. Mol Cell Biol 14: 3350–3363.
Wolff AC, Hammond ME, Schwartz JN, Hagerty KL, Allred DC, Cote RJ, Dowsett M, Fitzgibbons PL, Hanna WM, Langer A, McShane LM, Paik S, Pegram MD, Perez EA, Press MF, Rhodes A, Sturgeon C, Taube SE, Tubbs R, Vance GH, van de Vijver M, Wheeler TM, Hayes DF American Society of Clinical Oncology/College of American Pathologists (2007) American Society of Clinical Oncology/College of American Pathologists guideline recommendations for human epidermal growth factor receptor 2 testing in breast cancer. Arch Pathol Lab Med 131: 18–43.
Wu WJ, Hu KS, Wang DS, Zeng ZL, Zhang DS, Chen DL, Bai L, Xu RH (2013) CDC20 overexpression predicts a poor prognosis for patients with colorectal cancer. J Transl Med 11: 142.
Yamanaka S, Campbell NR, An F, Kuo SC, Potter JJ, Mezey E, Maitra A, Selaru FM (2012) Coordinated effects of microRNA-494 induce G(2)/M arrest in human cholangiocarcinoma. Cell Cycle 11: 2729–2738.
Yu R, Lu W, Chen J, McCabe CJ, Melmed S (2003) Overexpressed pituitary tumor-transforming gene causes aneuploidy in live human cells. Endocrinology 144: 4991–4998.
Yuan B, Xu Y, Woo JH, Wang Y, Bae YK, Yoon DS, Wersto RP, Tully E, Wilsbach K, Gabrielson E (2006) Increased expression of mitotic checkpoint genes in breast cancer cells with chromosomal instability. Clin Cancer Res 12: 405–410.
Acknowledgements
We wish to thank Mrs Sinikka Kollanus from Department of Pathology for technical assistance. The study is supported by Jyväskylä Central Hospital, Turku University Central Hospital and Cancer Society of South-Western Finland.
Author information
Authors and Affiliations
Corresponding author
Additional information
This work is published under the standard license to publish agreement. After 12 months the work will become freely available and the license terms will switch to a Creative Commons Attribution-NonCommercial-Share Alike 3.0 Unported License.
Rights and permissions
From twelve months after its original publication, this work is licensed under the Creative Commons Attribution-NonCommercial-Share Alike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article

Cite this article
Karra, H., Repo, H., Ahonen, I. et al. Cdc20 and securin overexpression predict short-term breast cancer survival. Br J Cancer 110, 2905–2913 (2014). https://doi.org/10.1038/bjc.2014.252
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/bjc.2014.252
Keywords
This article is cited by
-
p53 directly downregulates the expression of CDC20 to exert anti-tumor activity in mantle cell lymphoma
Experimental Hematology & Oncology (2023)
-
Differentially expressed discriminative genes and significant meta-hub genes based key genes identification for hepatocellular carcinoma using statistical machine learning
Scientific Reports (2023)
-
Epigenetic reprogramming of cell cycle genes by ACK1 promotes breast cancer resistance to CDK4/6 inhibitor
Oncogene (2023)
-
Structure-based virtual screening for identification of potential CDC20 inhibitors and their therapeutic evaluation in breast cancer
3 Biotech (2023)
-
Breast cancer prediction and categorization in the molecular era of histologic grade
Multimedia Tools and Applications (2023)
