
Abstract
RNA encoding the human serotonin 5-HT2C receptor (5-HT2CR) undergoes adenosine-to-inosine RNA editing events at five positions, resulting in an alteration of amino acids in the second intracellular loop. Several edited 5-HT2CRs possess a reduced G-protein coupling efficiency compared to the completely non-edited isoform. The current studies show that the efficacy of the hallucinogenic drug lysergic acid diethylamide and of antipsychotic drugs is regulated by RNA editing, suggesting that alterations in editing efficiencies or patterns might result in the generation of a 5-HT2CR population differentially responsive to serotonergic drugs. An examination of the efficiencies of RNA editing of the 5-HT2CR in prefrontal cortex of control individuals vs. subjects diagnosed with schizophrenia or major depressive disorder revealed no significant differences in RNA editing among the three populations. However, subjects who had committed suicide (regardless of diagnosis) exhibited a statistically significant elevation of editing at the A-site, which is predicted to change the amino acid sequence in the second intracellular loop of the 5-HT2CR. These findings suggest that alterations in RNA editing may contribute to or complicate therapy in certain psychiatric disorders.
Similar content being viewed by others
Main
The involvement of serotonin (5-HT) in psychiatric disorders has been hypothesized based largely on pharmacological studies. Atypical antipsychotic drugs, which have superior therapeutic and side effect profiles, bind with high affinity to various subtypes of serotonin receptors, including 5-HT2A and 5-HT2C receptors (Roth and Meltzer 1995). In addition, hallucinogenic drugs such as lysergic acid diethylamide (LSD) and psilocybin, which produce behavioral and cerebral metabolic disturbances that resemble symptoms of acute schizophrenia (Vollenweider et al. 1997, 1998), are agonists at 5-HT2A and 5-HT2C receptors (Burris et al. 1991; Egan et al. 1998). The pharmacological properties of the 5-HT2C receptor (5-HT2CR) have also suggested a role in depression (Moreau et al. 1996; Martin et al. 1998) as well as anxiety (Kennett et al. 1997).
The 5-HT2CR is a G-protein coupled receptor, linked to activation of phospholipase C with the production of inositol phosphates and diacylglycerol (Hoyer et al. 1994; Sanders-Bush and Canton 1995). A recently identified genetic polymorphism of the coding region of the 5-HT2CR, (Cys23Ser, Lappalainen et al. 1995), has been associated with psychotic symptoms in Alzheimer's disease (Holmes et al. 1998) and with an increased risk for hospitalization in schizophrenic patients (Segman et al. 1997). No other 5-HT2CR polymorphisms have been identified within the coding region; however, we have recently discovered that the 5-HT2CR is modified by RNA editing, a post-transcriptional event that generates 5-HT2CR isoforms with distinct functional properties (Burns et al. 1997). Five positions, termed A, B, C, D, and E, within the human 5-HT2CR undergo adenosine-to-inosine RNA editing events in a region that encodes the predicted second intracellular loop of the receptor. Concurrent editing at all five sites leads to a fully edited transcript, encoding a novel receptor isoform with VGV at positions 156, 158 and 160 instead of INI in the non-edited isoform (Niswender et al. 1999; Fitzgerald et al. 1999).
The VGV receptor isoform (5-HT2C-VGVR) and another extensively edited isoform (5-HT2C-VSVR) couple less efficiently to the phospholipase C signaling cascade, resulting in a decreased potency for serotonin (Burns et al. 1997; Niswender et al. 1999; Fitzgerald et al. 1999). RNA editing also generates 5-HT2CR isoforms that differ in their ability to interact with G-proteins in the absence of agonist, a phenomenon termed constitutive activity. The h5-HT2C-INI and h5-HT2C-VGV receptors were shown to behave at the extremes of the constitutive activity continuum with h5-HT2C-INI R exhibiting the highest degree of constitutive G-protein coupling and h5-HT2C-VGVR, the lowest (Herrick-Davis et al. 1999). These differential coupling abilities result in distinct affinities of the receptor isoforms for ligands that prefer to bind to either the coupled or uncoupled form of the receptor (Niswender et al. 1999).
In this article, we show that the fully edited human 5-HT2C receptor isoform (h5-HT2C-VGV) exhibits a marked reduction in sensitivity to LSD and atypical antipsychotic drugs, suggesting a possible role for 5-HT2CR editing in the etiology and pharmacotherapy of schizophrenia. To test this hypothesis, we compared the efficiency of editing of the 5-HT2CR mRNA in prefrontal cortex of subjects diagnosed with schizophrenia vs. matched controls. A second group of psychiatric patients diagnosed with major depression was analyzed to test the specificity of any observed changes. Although there were no significant differences in RNA editing as a function of diagnosis, a subset of psychiatric patients who had committed suicide exhibited a statistically significant elevation of editing at the A-site, resulting in a predicted increase in 5-HT2CRs containing a valine at amino acid 156. These studies suggest that editing alterations of the 5-HT2CR may play a role in suicide.
MATERIALS AND METHODS
Phosphoinositide Hydrolysis Assay and Radioligand Binding
Transiently transfected COS-7 cells were generated and used for inositol monophosphate analysis as previously described (Herrick-Davis et al. 1997). For isolation of inositol monophosphates from stable NIH-3T3 cells lines (Niswender et al. 1999), cells were pretreated with antagonists for 30 minutes in the presence of 10 mM lithium chloride and 10 μM pargyline. Agonists were then added and incubations continued for 15 minutes unless otherwise noted. [3H]-Inositol monophosphates were isolated by column chromatography as previously described (Barker et al. 1994). Radioligand binding competition assays in transiently transfected COS-7 cells were performed as previously described using 1 nM 3H-mesulergine (Herrick-Davis et al. 1999). For stable cell lines, radioligand binding was assayed in crude membrane homogenates prepared in 50 mM Tris-HCl, 10 mM MgCl2, pH 7.5, as previously described (Barker et al. 1994) with the exception that centrifugation was omitted. Data were analyzed using the GraphPad Prism® software.
Subjects and Tissue Collection
Brain tissue was obtained at autopsy from the Cuyahoga County Coroner's Office, Cleveland, Ohio, USA. Tissue samples of left prefrontal cortex (areas 8, 9) located on the superior frontal gyrus were dissected, frozen in isopentane cooled in dry ice, and stored at −80°C.
The study was performed in compliance with policies of an institutional review board and informed written consent was obtained from the next of kin for all subjects. Tissue samples were collected from 13 subjects with schizophrenia (7 males and six females; 44.5 ± 10.5 years of age; 16.5 ± 6.1 hours post mortem interval; mean ± SD), 13 subjects with major depression (9 males and four females; 45.5 ± 15.2 years of age; 18.9 ± 4.1 hours post mortem interval), and 13 psychiatrically-normal comparison subjects (10 males and three females; 45.9 ± 12.2 years of age; 18.0 ± 7.4 hours post mortem interval). The cause of death was ruled by the County Coroner. Six subjects with schizophrenia died from suicide, and twelve subjects with major depression died from suicide. Eleven normal control subjects died from natural or accidental causes and two died from homicide.
Blood and urine samples from all subjects were examined by the toxicology laboratory of the County Coroner's Office. As previously described, qualitative and quantitative assays were used to detect various classes of psychoactive compounds (Stockmeier et al. 1997). All subjects with schizophrenia had a prescription for an antipsychotic medication at some time in their lives, and seven had such a prescription filled within the last month of life. Of those subjects with major depression, five had a prescription for an antidepressant medication at some time in their lives, and four had such a prescription within the last month of life. Additionally, two subjects with major depression had a prescription for an antipsychotic medication during their lifetime, with one receiving such a prescription during the last month of life.
Retrospective Psychiatric Assessments
Retrospective psychiatric assessments were used as previously described to evaluate psychiatric diagnoses among the subjects (Stockmeier et al. 1997; Jope et al. 1998). Based on diagnostic interviews, the following three groups were identified: (1) subjects with a current diagnosis of schizophrenia, (2) subjects with a current diagnosis of major depression, and (3) control subjects without evidence of an Axis I psychiatric disorder. At three to six months after the death, a trained interviewer met with a knowledgeable informant who either lived with or had frequent weekly contact with the deceased, and administered a structured diagnostic interview. Data on lifetime and current mental illness in the deceased were gathered with a modified Schedule for Affective Disorders and Schizophrenia; lifetime version (SADS-L; Endicott and Spitzer 1978). For three subjects with schizophrenia, only hospitalization records were available for making the Axis I diagnosis.
Information from the retrospective clinical interview, the coroner's office, previous hospitalizations, and medical records were independently evaluated by a clinical psychologist and a psychiatrist for assigning Axis I disorders, where appropriate. A consensus diagnosis was reached in conference using all sources of information, and the final diagnosis was compatible with the Diagnostic and Statistical Manual of Mental Disorders, fourth edition, DSM-IV; American Psychiatric Association 1994, classification. Among the subjects with schizophrenia, one subject had a current comorbid diagnosis of polysubstance abuse, and two other subjects had previous diagnoses of polysubstance abuse and alcohol dependence. Of those subjects meeting criteria for major depression, two had a comorbid diagnosis of dysthymia, and two had a previous (non-active) diagnosis of alcohol abuse. The control subjects did not meet criteria for an Axis I diagnosis, although one met criteria for a previous diagnosis of alcohol abuse. Subjects from each diagnostic group were matched in a yoked set by age and post-mortem interval, and coded to conceal the psychiatric diagnosis.
Preparation of RNA, RT-PCR, and Editing Efficiency Analyses
Frozen tissue samples were shipped overnight on dry ice to Vanderbilt University. Vanderbilt investigators were blinded to sample identity until all experimental analyses had been completed.
Total RNA was extracted using the Tri-Reagent® protocol (Molecular Research Centers, Cincinnati, OH). Reverse transcription-polymerase chain reaction (RT-PCR) of 5-HT2CR messenger RNA (mRNA) was performed as previously described (Burns et al. 1997) with the following modifications: oligo VU195 5′ GCAGTAACATCAAAGCTTGTCGGCG 3′ (coordinates 723–747 relative to the translation start site of the human 5-HT2CR cDNA [Saltzman et al. 1991]; bold sequence indicates an introduced Hind III restriction site) was employed for cDNA synthesis in the presence of 5 mM MgCl2, 1X AMV reverse transcriptase buffer (Promega Corporation, Madison, WI), 1 mM deoxynucleotide triphosphates (dNTPs), 20 units of RNasin, and six units of AMV reverse transcriptase in a total volume of 20 μl. Samples were incubated at 42°C for 45 minutes, 99°C for 5 minutes, and then place on ice. PCR reactions were performed in a total volume of 100 μl in the presence of 1X Taq polymerase buffer and 0.25 units of Taq polymerase (Promega), 1.5 mM MgCl2, 200 μM dNTPs, and 500 ng each of primers ESB-15 5′ CCAGGGAATTCAAACTTTGGTTGCTTAAGACTGAAGC 3′ (-30 to -5; Xie et al. 1996) and VU195 or, in some cases, VU186 5′ ATTAGAATTCTATTTGTGCCCCGTCTGG 3′ (372–389; Saltzman et al. 1991; bold sequence indicates introduced EcoRI restriction sites). Amplification proceeded for 1 min, 15 sec at 94°C, 1 min, 15 sec at 50°C, and 2 min, 30 sec at 72°C for 35 cycles. Products were purified on a 2% agarose gel and primer extension analyses at the A-, C- and D-sites were performed as described previously (Burns et al. 1997) and as illustrated in Figure 1. For sequencing analyses, PCR fragments were digested with EcoRI and Hind III (Promega) and unidirectionally subcloned into pBKSII- (Stratagene). Single isolates were sequenced using the Promega fmole sequencing system®. Samples that did not amplify after three to five attempts were excluded from the analyses.

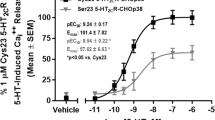
Differential responses of LSD in NIH-3T3 cells stably expressing human edited 5-HT2CRs. Typical dose-response curves for the generation of [3H]-inositol monophosphates after 5-HT (squares) or LSD (triangles) stimulation are shown for the h5-HT2C-INI receptor (Panel A) and h5-HT2C-VGV receptor (Panel B). The values plotted are the mean ± standard error of triplicate determinations and are representative of four to sixteen independent analyses performed in triplicate. The EC50 values (mean ± SEM) for 5-HT were 2.3 ± 0.95 nM and 59.3 ± 15.7 nM for the h5-HT2C-INI receptor and h5-HT2C-VGV receptor, respectively. The EC50 value for LSD at the h5-HT2C-INI receptor was 9.8 ± 1.3 nM. Receptor densities were 250 and 1200 fmol/mg protein for the h5-HT2C-INI receptor and h5-HT2C-VGV receptor, respectively
To assess the effects of post mortem interval on RNA editing status, rats were sacrificed and brains removed immediately or 18 hours after storing the body in a cold room. Editing status at the A-, C-, and D-sites was examined as described earlier. These experiments revealed no alterations in editing percentages, indicating that there is not a preferential degradation of either edited or nonedited RNA.
Statistical Analyses
Data were analyzed by multivariate analyses of covariance with diagnosis, race and sex as independent variables. Age and post mortem interval were included in all analyses. To determine if editing differed in individuals who committed suicide, the subjects were subdivided into suicide and non-suicide groups (regardless of diagnosis) and re-analyzed.
RESULTS
The h5-HT2C-VGV R was Unresponsive to the Hallucinogenic Agent (+) Lysergic Acid Diethylamide
In earlier studies using fibroblasts transiently expressing the receptor isoforms, the h5-HT2C-VGVR exhibited the most dramatic reduction in agonist potency relative to the non-edited h5-HT2C-INIR (Niswender et al. 1999); therefore, the current studies have focused on these two receptor isoforms. (+) LSD, a hallucinogenic ergot derivative, has been shown to have agonist properties in a neuroblastoma cell line expressing human 5-HT2C-INI receptors (Newton et al. 1996). In the present studies, LSD produced a robust phosphoinositide hydrolysis response in fibroblasts stably expressing the h5-HT2C-INIR (Figure 1); however, the hallucinogen failed to elicit a reproducible response in fibroblasts expressing the h5-HT2C-VGVR (Figure 1). In contrast, the maximum 5-HT signal was nearly double in cells expressing the h5-HT2C-VGV R, likely reflecting the higher density of receptors in this cell line. Radioligand binding analyses revealed that LSD bound with similar high affinity to both receptor isoforms (18 ± 7 nM and 14 ± 2 nM for h5-HT2C-INIR and h5-HT2C-VGVR, respectively). These data are in agreement with those presented by Fitzgerald et al. (1999) after expression of the h5-HT2C-INI and h5-HT2C-VGV receptors in HEK 293 cells, showing that the lack of an LSD response is not dependent on the cellular expression system. Further analysis of LSD interaction with these receptor variants indicated that preincubation with LSD completely blocked 5-HT-mediated phosphoinositide hydrolysis in cells expressing the 5-HT2C-VGV R, but not in cells expressing the h5-HT2C-INI R (Table 1). These results indicate that LSD may function as a highly effective antagonist of certain 5-HT2CR isoforms in the brain.
Distinction in The Ability of Typical and Atypical Antipsychotic Drugs to Inhibit Basal Activity of The h5-HT2C-INIR
We have shown that the h5-HT2C-INIR exhibits an enhanced ability to spontaneously isomerize to a G-protein coupling-competent conformation when compared to the h5-HT2C-VGVR variant (Niswender et al. 1999). The spontaneous G-protein interaction results in high basal activity of the h5-HT2C-INIR, a phenomenon termed constitutive activity. The atypical antipsychotic agent, clozapine, is classified as an inverse agonist at the rat 5-HT2CR, blocking the ability of the receptor to signal in a constitutive, agonist-independent manner (Westphal and Sanders-Bush 1994). To evaluate further the properties of clozapine and other antipsychotic drugs, h5-HT2C-INI and h5-HT2C-VGV receptors were transiently expressed in COS-7 cells, a system that reproducibly produces the high levels of receptor expression necessary for accurate determination of basal phosphoinositide hydrolysis. Transfected cells expressing a density of approximately 2500 fmol/mg were treated with chlorpromazine, thioridazine, clozapine, or loxapine and examined for repression of basal inositol monophosphate generation (Figure 2). Both loxapine and clozapine were able to significantly reduce basal inositol phosphate production of the h5-HT2C-INIR while chlorpromazine and thioridazine had no effect (Figure 2). In additional experiments, 1 μM concentrations of thioridazine and chlorpromazine were found to attenuate 5-HT-mediated inositol phosphate formation (data not shown), demonstrating that the drugs are active at this concentration. Due to an inherent lack of constitutive activity, the h5-HT2C-VGVR had low basal activity, which was not altered by either typical or atypical antipsychotic agents (Figure 2).

Blockade of constitutive activity of transiently expressed h5-HT2C-INI and h5-HT2C-VGV receptors by antipsychotic drugs. cDNAs for the 5-HT2C-INIR and 5-HT2C-VGVR were transiently expressed in COS-7 cells. Cells were pretreated with 1-μM concentrations of each antipsychotic agent for 15 minutes and [3H]-inositol monophosphate formation (±1 μM 5-HT) was measured after a 35 minute incubation. Data are represented as the mean ± standard error (minus vector alone control) of four independent experiments. * indicates that these values were significantly different from basal (p < .01 in all cases)
Edited Receptors Exhibit Unique Affinities for Clozapine and Loxapine but Not Chlorpromazine and Thioridazine
The extended ternary complex model of receptor:G-protein coupling predicts that constitutively active receptors exist in two states, presumably reflecting different receptor conformations: an inactive R state and an active R* conformation (Samama et al. 1993). Constitutively active receptors spontaneously isomerize from the R to the R* form, which is competent to interact with G-proteins and exhibits a higher affinity for agonists. Inverse agonists are so named due to their ability to decrease constitutive activity by binding to and stabilizing the receptor in an unproductive R conformation. We have previously shown that the enhanced ability of the h5-HT2C-INIR to spontaneously isomerize to an active conformation results in higher affinities of agonists for this isoform compared to the 5-HT2C-VGVR (Niswender et al. 1999). Conversely, it would be predicted that inverse agonists should have higher affinity for the predominantly uncoupled h5-HT2C-VGVR. To correlate blockade of constitutive activity with potential differences in affinity, we examined the binding profiles of chlorpromazine, thioridazine, clozapine, and loxapine at the h5-HT2C-INIR and h5-HT2C-VGVR isoforms. Chlorpromazine and thioridazine bound with similar affinity to both receptor isoforms (p = .852 and .553, respectively, Table 2). In contrast, loxapine, an inverse agonist, bound with significantly higher affinity to the predominantly uncoupled h5-HT2C-VGVR isoform (p = .0003; Table 2). The competition binding curve for the other inverse agonist clozapine was best fit by a two-site model when interacting with the h5-HT2C-INIR but not the h5-HT2C-VGVR. The Ki value for the single low affinity site at the h5-HT2C-VGVR was significantly lower than was the value at the low affinity site for h5-HT2C-INIR (p < .05, Table 2). These results indicate that, at concentrations close to their binding affinity, atypical antipsychotic drugs might preferentially interact with receptor isoforms with low constitutive activity.
5-HT2CR Editing Status in Prefrontal Cortex of Control Subjects and Subjects with Schizophrenia or Major Depression
The profile of 5-HT2CR isoforms was determined in samples of control brain by subcloning the PCR amplified fragments and sequencing 100 clones (Table 3). In prefrontal cortex, the most abundant isoform, VSV, was edited at four (A, B, C, and D) sites and the next most abundant isoform, VGV, was edited at all five sites. A similar profile was seen in the hippocampus, although in the hypothalamus (Fitzgerald et al. 1999), the profile differs significantly with the VNV isoform most prominent. This region-specific pattern of editing suggests that the isoforms may have unique functions within specific brain regions.
To investigate the potential relationship between edited 5-HT2CR isoforms and psychiatric diagnosis, we performed primer extension assays of the editing status of 5-HT2CR RNA in post-mortem tissue from subjects diagnosed with schizophrenia or major depression compared to control subjects. Total RNA was isolated from the prefrontal cortex (Brodmann areas 8 and 9) of 13 control subjects, 13 subjects diagnosed with major depression, and 13 individuals with various classifications of schizophrenia. There were no significant differences in age or post mortem interval between any of the groups (Tables 4, 5, 6 ). The RNA samples were subjected to RT-PCR amplification and editing efficiency at the A-, C- and D-sites was examined using primer extension analyses (Figure 3). As shown in Table 4, editing levels at the A-, C- and D-sites were equal in all three groups of subjects. Editing percentages were also not significantly different as a function of age, race, sex, or post mortem interval. These studies suggest that there is not a global alteration in editing that correlates with major depression or schizophrenia in the populations that we examined. A separate analysis of suicide (n = 18) vs. non-suicide (n = 21) samples revealed that the suicide population exhibited significantly higher levels of editing at the A-site (Figure 4; p < .05). Editing at the C-site and D-site were not significantly different (p = .235 and .157, respectively). Exclusion of control subjects from the analysis and comparison of suicide (n = 18) vs. non-suicide psychiatric samples (n = 8) also revealed a significant elevation of editing at the A-site (p = .029) and a trend for an increase at the D-site (p = .063).

Primer extension analyses of RT-PCR samples. Panel A: Schematic of the genomic DNA sequence of the human 5-HT2CR; the position of the five editing sites and the amino acid changes induced by editing at these positions is shown. Panel B: Primer extension assay design to assess editing at the A-site. A radiolabeled 18 nucleotide primer (designated with an asterisk) is annealed to the antisense strand of the RT-PCR template and extended in the presence of the indicated deoxy (d) or dideoxy (dd) nucleotides with T7 RNA polymerase (Burns et al. 1997). The sizes of predicted products produced by edited (21 nucleotides) and nonedited (22 nucleotides) templates are indicated. Panel C: Primer extension assay design to assess editing at the E-, C-, and D- sites. Degenerate positions within the extending oligonucleotide are indicated as R and the predicted sizes of products are shown. While an assessment of editing at the E-site is theoretically possible with this design, the close proximity of this site to the primer coupled with the low editing efficiency at this position precluded a quantitative measurement of editing at this site in all samples. Editing at the internal B-site also could not be accurately assessed using multiple primer designs; for these reasons, B- and E-site editing was only assessed when RT-PCR products were individually subcloned and sequenced (Table 3). Panel D: Primer extension analysis of editing at the A-, C-, and D-sites within RT-PCR samples prepared from the J series of subjects (J1 [major depression group], J2 [schizophrenic group] and J3 [control group]). The positions and sizes of the edited and nonedited products for each site are indicated. * edited E products could not be accurately quantitated in all samples. # indicates a nonspecific product; the appearance of this fragment is irregular and does not appear to correspond to edited or nonedited fragments at any site. The ratio of edited to nonedited bands was determined using phosphorimager analyses and Imagequant® software

Editing efficiencies at the A-, C- and D-sites in prefrontal cortex of suicide vs. nonsuicide subjects. The level of editing at the A-, C- and D- positions was determined by primer extension analysis. Each subject sample was analyzed at least twice and a mean of these repetitive assays used as the value for that sample. Black bars represent individuals who committed suicide; white bars represent nonsuicide subjects. Results were analyzed by one-way ANOVA. * indicates p < .05
DISCUSSION
RNA editing is an important mechanism for the generation of molecular diversity (Simpson and Emeson 1996). Adenosine-to-inosine conversion at the level of 5-HT2CR pre-mRNA alters amino acids in the predicted second intracellular loop (Burns et al. 1997), a region of Gq/11-coupled receptors important for G-protein coupling (Moro et al. 1993; Arora et al. 1995; Blin et al. 1995; Verrall et al. 1997). We have previously shown that certain rat and human edited 5-HT2CR isoforms couple less efficiently to the phospholipase C signal transduction cascade (Burns et al. 1997). This coupling deficiency is evident both in the spontaneous activity and in the activity generated by agonist binding (Niswender et al. 1999; Fitzgerald et al. 1999; Herrick-Davis et al. 1999).
The ability of distinct edited 5-HT2CR isoforms to interact uniquely with the intracellular signaling machinery led to the hypothesis that these isoforms would also exhibit differential responses to drugs. We have previously shown that the hallucinogenic drugs DOI and DMT exhibit lower affinity and potency when interacting with the edited h5-HT2C-VGV R vs. the non-edited h5-HT2C-INI R (Niswender et al. 1999). The agonist LSD, on the other hand, is unique in its inability to elicit a reproducible phosphoinositide hydrolysis response in cells expressing h5-HT2C-VGV receptors (Fitzgerald et al. 1999; present results). Mechanisms to explain this unique property of LSD are currently unknown. Subtle differences in the site of binding of LSD and 5-HT have been described for the 5-HT2A receptor (Almaula et al. 1996), a protein with high homology to the 5-HT2CR. LSD binding to a distinct site on the 5-HT2CR may propagate a signal effectively only when interacting with isoforms containing a specific combination of amino acids within the second intracellular loop. This phenotype would be independent of binding affinity, consistent with the indistinguishable affinity of LSD for the h5-HT2C-INI and h5-HT2C-VGV isoforms (current results; Fitzgerald et al. 1999). Our finding that LSD interaction with the h5-HT2C-VGVR isoform blocks, rather than mimics 5-HT, is also consistent with a subtle difference in LSD binding, which fails to propagate a coupling-competent receptor confirmation. Preliminary examination of LSD's ability to induce arachidonic acid signaling revealed an inability to stimulate arachidonic acid metabolism when interacting with the h5-HT2C-VGVR, while stimulation at the 5-HT2C-INIR remains intact (K. Berg, personal communication). These observations indicate that LSD is incapable of transducing an agonist signal when interacting with the h5-HT2C-VGVR. The capacity of RNA editing to eliminate the agonist activity of LSD and reveal a potent antagonist activity suggests that the repertoire of edited 5-HT2CR isoforms may dictate some of the actions of LSD.
Similar to the rat 5-HT2C-INI R expressed in NIH 3T3 cells (Barker et al. 1994), the human 5-HT2C-INIR expressed in COS-7 cells exhibits substantial constitutive activity (Niswender et al. 1999; Herrick-Davis et al. 1999; current results). However, little or no constitutive activity was found with the human 5-HT2C-INIR expressed in HEK cells (Fitzgerald et al. 1999), suggesting that the cellular microenviroment is important for detection of receptor constitutive activity. At the rat 5-HT2C-INIR, the atypical antipsychotic agent clozapine behaves as an inverse agonist, defined as an antagonist that not only blocks the effect of an agonist but also blocks the spontaneous, constitutive activity of the receptor (Westphal and Sanders-Bush 1994). Based on the compromised constitutive activity found in the extensively edited 5-HT2CR isoforms, we hypothesized that antipsychotic drugs might exhibit distinct properties depending upon the edited status of the receptor. In studies assessing receptor-dependent basal activity (termed constitutive activity), the antipsychotic drugs clozapine and loxapine decreased basal phosphoinositide hydrolysis by the 5-HT2C-INIR isoform, but did not have this effect at the 5-HT2C-VGVR. In contrast, the typical antipsychotic drugs chlorpromazine and thioridazine did not alter the constitutive activity of the 5-HT2C-INIR, even though they blocked the ability of 5-HT to elicit phosphoinositide hydrolysis. Thus, the typical antipsychotics behaved as neutral antagonists, blocking agonist stimulated activity, but not spontaneous activity. Clozapine and loxapine, on the other hand, function as inverse agonists at the 5-HT2C-INIR, blocking constitutive as well as agonist stimulated activity. Interestingly loxapine, like clozapine, has properties consistent with a classification as an atypical antipsychotic drug. For example, loxapine has a 5-HT2A/D2 ratio of affinities comparable to clozapine and other atypical antipsychotic drugs (Meltzer et al. 1989). In addition, studies in humans suggest that loxapine has atypical properties at low doses (Meltzer and Jayathilake 1999; Glazer 1999). Therefore, we hypothesize that atypical antipsychotic drugs behave as inverse agonists at the human 5-HT2CR, while typical antipsychotic drugs behave as neutral antagonists. Consistent with this hypothesis, the widely acknowledged atypical antipsychotic drug risperidone mimicked clozapine and loxapine, decreasing constitutive phosphoinositide hydrolysis as well as blocking agonist stimulated activity (data not shown). Although there is no experimental evidence for altered basal 5-HT2CR signaling in schizophrenia, it is possible that there may be different therapeutic outcomes for drugs that possess inverse agonism vs. drugs that simply block endogenous ligands.
In addition to differences in the modulation of edited 5-HT2C receptors by agonists, drugs classified as inverse agonists would be predicted to show distinctions in binding affinity dependent upon the unique coupling profiles of receptor isoforms. Agonists and inverse agonists exhibit differential preferences for the inactive R and the active R* state of G-protein coupled receptors. Agonists bind with higher affinity to the R* form, whereas inverse agonists interact with higher affinity with the inactive R form; neutral antagonists have equal affinity for both forms (for review, see Milligan et al. 1995). This prediction was confirmed, since the inverse agonists clozapine and loxapine exhibit higher binding affinity at the 5-HT2C-VGVR than 5-HT2C-INIR. In contrast, the typical antipsychotics are equipotent at the two isoforms, consistent with the conclusion that they function as neutral antagonists. Recent studies suggest that inverse agonists at the 5-HT2CR, but not neutral antagonists, cause profound receptor supersensitivity after prolonged exposure (Berg et al. 1999). Receptor supersensitivity may explain the clinical phenomenon of clozapine-withdrawal psychosis (Meltzer et al. 1996), in which a high incidence of rapid relapse occurs in schizophrenic patients when clozapine treatment is discontinued. Based on the results of the current manuscript, receptor supersensitivity would not be predicted after typical neuroleptics, consistent with the clinical observation that withdrawal psychosis is not common after discontinuation of typical neuroleptics (Viguera et al. 1997).
The above studies suggest that the interaction of agonists and inverse agonists with edited 5-HT2C receptors may be regulated by the coupling status of the isoforms, resulting in differential abilities of compounds to stimulate or antagonize inositol phosphate signaling depending on the repertoire of expressed edited receptors. As a first step at understanding the clinical significance of this distinction, we compared the efficiency of RNA editing of 5-HT2C receptors in prefrontal cortex of subjects with schizophrenia, normal controls, and individuals diagnosed with major depression. While no global changes in editing were noted among these three subject groups, re-analysis of suicide vs. non-suicide subjects revealed that editing at the A-site was elevated in individuals who committed suicide, independent of diagnosis. Editing at the A-site predicts the formation of a receptor isoform with a single amino acid change (Iso to Val) within the putative second intracellular loop of the receptor (Burns et al. 1997). Although the functional significance of such an editing change is not entirely clear, the analogous position has been proposed to play a key role in the transduction of the binding signal for another Gq/11-coupled receptor, the gonadotropin-releasing hormone receptor (Ballesteros et al. 1998).
In summary, the present studies show that an extensively edited isoform of the 5-HT2CR responds uniquely to both the hallucinogenic drug LSD and antipsychotic medications. These observations prompted an examination of 5-HT2CR editing status in brains of subjects with schizophrenia vs. normal controls and individuals with psychotic depression. A comparison of the efficiency of editing revealed no statistically significant differences in RNA editing capacity between the three groups of subjects. However, a further statistical analysis revealed a small, but significant increase in editing capacity at the A-site in individuals who committed suicide, regardless of diagnosis. Further studies are warranted to assess the editing pattern of individual mRNAs and to compare the repertoire of edited receptors in suicidal vs. nonsuicidal subjects.
References
Almaula N, Ebersole BJ, Zhang D, Weinstein H, Sealfon SC . (1996): Mapping the binding site pocket of the serotonin 5-hydroxytryptamine2A receptor. Ser3.36(159) provides a second interaction site for the protonated amine of serotonin but not of lysergic acid diethylamide or bufotenin. J Biol Chem 271: 14672–14675
Arora KK, Sakai A, Catt KJ . (1995): Effects of second intracellular loop mutations on signal transduction, internalization of the gonadotropin-releasing hormone receptor. J Biol Chem 270: 22820–22826
Ballesteros J, Kitanovic S, Guarnieri F, Davies P, Fromme BJ, Konvicka K, Chi L, Millar RP, Davidson JS, Weinstein H, Sealfon SC . (1998): Functional microdomains in G-protein-coupled receptors. The conserved arginine-cage motif in the gonadotropin-releasing hormone receptor. J Biol Chem 273: 10445–10453
Barker EL, Westphal RS, Schmidt D, Sanders-Bush E . (1994): Constitutively active 5-hydroxytryptamine2C receptors reveal novel inverse agonist activity of receptor ligands. J Biol Chem 269: 11687–11690
Berg KA, Stout BD, Cropper JD, Maayani S, Clarke WP . (1999): Novel actions of inverse agonists on 5-HT2C receptor systems. Mol Pharmacol 55: 863–872
Blin N, Yun J, Wess J . (1995): Mapping of single amino acid residues required for selective activation of Gq/11 by the m3 muscarinic acetylcholine receptor. J Biol Chem 270: 17741–17748
Burns CM, Chu H, Rueter SM, Hutchinson LK, Canton H, Sanders-Bush E, Emeson RB . (1997): Regulation of serotonin-2C receptor G-protein coupling by RNA editing. Nature 387: 303–308
Burris KD, Breeding M, Sanders-Bush E . (1991): (+)Lysergic acid diethylamide, but not its nonhallucinogenic congeners, is a potent serotonin 5HT1C receptor agonist. J Pharmacol Exp Ther 258: 891–896
Cheng Y, Prusoff WH . (1973): Relationship between the inhibition constant (K1) and the concentration of inhibitor which causes 50 per cent inhibition (I50) of an enzymatic reaction. Biochem Pharmacol 22: 3099–3108
Egan CT, Herrick-Davis K, Miller K, Glennon RA, Teitler M . (1998): Agonist activity of LSD and lisuride at cloned 5HT2A and 5HT2C receptors. Psychopharmacology (Berl) 136: 409–414
Endicott J, Spitzer RL . (1978): A diagnostic interview: the schedule for affective disorders and schizophrenia. Arch Gen Psychiatry 35: 837–844
Fitzgerald LW, Iyer G, Conklin DS, Krause CM, Marshall A, Patterson JP, Tran DP, Jonak GJ, Hartig PR . (1999): Messenger RNA editing of the human serotonin 5-HT2C receptor. Neuropsychopharmacology 21: 82S–90S
Glazer WM . (1999): Does loxapine have “atypical” properties? Clinical evidence. J Clin Psychiatry 60: 42–46
Herrick-Davis K, Grinde E, Niswender CM . (1999): Serotonin 5-HT2C Receptor RNA editing alters receptor basal activity: implications for serotonergic signal transduction. Neurochemistry 73: 1711–1717
Herrick-Davis K, Egan C, Teitler M . (1997): Activating mutations of the serotonin 5-HT2C receptor. J Neurochem 69: 1138–1144
Holmes C, Arranz MJ, Powell JF, Collier DA, Lovestone S . (1998): 5-HT2A and 5-HT2C receptor polymorphisms and psychopathology in late onset Alzheimer's disease. Hum Mol Genet 7: 1507–1509
Hoyer D, Clarke DE, Fozard JR, Hartig PR, Martin GR, Mylecharane EJ, Saxena PR, Humphrey PP . (1994): International Union of Pharmacology classification of receptors for 5- hydroxytryptamine (Serotonin). Pharmacol Rev 46: 157–203
Jope RS, Song L, Grimes CA, Pacheco MA, Dilley GE, Li X, Meltzer HY, Overholser JC, Stockmeier CA . (1998): Selective increases in phosphoinositide signaling activity and G protein levels in postmortem brain from subjects with schizophrenia or alcohol dependence. J Neurochem 70: 763–771
Kennett GA, Wood MD, Bright F, Trail B, Riley G, Holland V, Avenell KY, Stean T, Upton N, Bromidge S, Forbes IT, Brown AM, Middlemiss DN, Blackburn TP . (1997): SB 242084, a selective and brain penetrant 5-HT2C receptor antagonist. Neuropharmacology 36: 609–620
Lappalainen J, Zhang L, Dean M, Oz M, Ozaki N, Yu DH, Virkkunen M, Weight F, Linnoila M, Goldman D . (1995): Identification, expression, and pharmacology of a Cys23-Ser23 substitution in the human 5-HT2C receptor gene (HTR2C). Genomics 27: 274–279
Martin JR, Bos M, Jenck F, Moreau J, Mutel V, Sleight AJ, Wichmann J, Andrews JS, Berendsen HH, Broekkamp CL, Ruigt GS, Kohler C, Delf AM . (1998): 5-HT2C receptor agonists: pharmacological characteristics and therapeutic potential. J Pharmacol Exp Ther 286: 913–924
Meltzer HY, Jayathilake K . (1999): Low-dose loxapine in the treatment of schizophrenia: is it more effective and more “atypical” than standard-dose loxapine? J Clin Psychiatry 60: 47–51
Meltzer HY, Lee MA, Ranjan R, Mason EA, Cola PA . (1996): Relapse following clozapine withdrawal: effect of neuroleptic drugs and cyproheptadine. Psychopharmacology (Berl) 124: 176–187
Meltzer HY, Matsubara S, Lee JC . (1989): The ratios of serotonin2 and dopamine2 affinities differentiate atypical and typical antipsychotic drugs. Psychopharmacol Bull 25: 390–392
Milligan G, Bond RA, Lee M . (1995): Inverse agonism: pharmacological curiosity or potential therapeutic strategy? Trends Pharmacol Sci 16: 10–13
Moreau JL, Bos M, Jenck F, Martin JR, Mortas P, Wichmann J . (1996): 5HT2C receptor agonists exhibit antidepressant-like properties in the anhedonia model of depression in rats. Eur Neuropsychopharmacol 6: 169–175
Moro O, Lameh J, Hogger P, Sadee W . (1993): Hydrophobic amino acid in the i2 loop plays a key role in receptor-G protein coupling. J Biol Chem 268: 22273–22276
Newton RA, Phipps SL, Flanigan TP, Newberry NR, Carey JE, Kumar C, McDonald B, Chen C, Elliott JM . (1996): Characterization of human 5-hydroxytryptamine2A and 5-hydroxytryptamine2C receptors expressed in the human neuroblastoma cell line SH-SY5Y: comparative stimulation by hallucinogenic drugs. J Neurochem 67: 2521–2531
Niswender CM, Copeland SC, Herrick-Davis K, Emeson RB, Sanders-Bush E . (1999): RNA editing of the human serotonin 5-hydroxytryptamine 2C receptor silences constitutive activity. J Biol Chem 274: 9472–9478
Roth BL, Meltzer HY . (1995): The Role of Serotonin in Schizophrenia. In: Bloom FE, Kupfer DJ (eds), Psychopharmacology: The Fourth Generation of Progress. New York, Raven Press Ltd. pp 1215–1227 (Ch. 102).
Saltzman AG, Morse B, Whitman MM, Ivanshchenko Y, Jaye M, Felder S . (1991): Cloning of the human serotonin 5-HT2 and 5-HT1C receptor subtypes. Biochem Biophys Res Commun 181: 1469–1478
Samama P, Cotecchia S, Costa T, Lefkowitz RJ . (1993): A mutation-induced activated state of the beta 2-adrenergic receptor. Extending the ternary complex model. J Biol Chem 268: 4625–4636
Sanders-Bush E, Canton H . (1995): Serotonin Receptors: Signal Transduction Pathways. New York, Raven Press
Segman RH, Ebstein RP, Heresco-Levy U, Gorfine M, Avnon M, Gur E, Nemanov L, Lerer B . (1997): Schizophrenia, chronic hospitalization and the 5-HT2C receptor gene. Psychiatr Genet 7: 75–78
Simpson L, Emeson RB . (1996): RNA editing. Ann Rev Neurosci 19: 27–52
Stockmeier CA, Dilley GE, Shapiro LA, Overholser JC, Thompson PA, Meltzer HY . (1997): Serotonin receptors in suicide victims with major depression. Neuropsychopharmacology 16: 162–173
Verrall S, Ishii M, Chen M, Wang L, Tram T, Coughlin SR . (1997): The thrombin receptor second cytoplasmic loop confers coupling to Gq-like G proteins in chimeric receptors. Additional evidence for a common transmembrane signaling and G protein coupling mechanism in G protein-coupled receptors. J Biol Chem 272: 6898–6902
Viguera AC, Baldessarini RJ, Hegarty JD, van Kammen DP, Tohen M . (1997): Clinical risk following abrupt and gradual withdrawal of maintenance neuroleptic treatment. Arch Gen Psychiatry 54: 49–55
Vollenweider FX, Leenders KL, Scharfetter C, Maguire P, Stadelmann O, Angst J . (1997): Positron emission tomography and fluorodeoxyglucose studies of metabolic hyperfrontality and psychopathology in the psilocybin model of psychosis. Neuropsychopharmacology 16: 357–372
Vollenweider FX, Vollenweider-Scherpenhuyzen MF, Babler A, Vogel H, Hell D . (1998): Psilocybin induces schizophrenia-like psychosis in humans via a serotonin-2 agonist action. Neuroreport 9: 3897–3902
Westphal RS, Sanders-Bush E . (1994): Reciprocal binding properties of 5-hydroxytryptamine type 2C receptor agonists and inverse agonists. Mol Pharmacol 46: 937–942
Xie E, Zhu L, Zhao L, Chang LS . (1996): The human serotonin 5-HT-2C receptor complete cDNA, genomic structure and alternatively spliced variant. Genomics 35: 551–561
Acknowledgements
This research was supported by a postdoctoral fellowship from the Pharmaceutical Research and Manufacturers of America Foundation (C.M.N.) and National Institutes of Health research grants NS35891 (R.B.E.), MH57019 (K.H.D.), MH45488 (C.A.S.), MH 34007 (E.S.B.) and The American Foundation for Suicide Prevention. The authors acknowledge the assistance of Bryan L. Roth, M.D., Ph.D., in the psychiatric evaluation of three subjects and Elizabeth Balraj, in the Cuyahoga County Coroner's Office.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Niswender, C., Herrick-Davis, K., Dilley, G. et al. RNA Editing of the Human Serotonin 5-HT2C Receptor: Alterations in Suicide and Implications for Serotonergic Pharmacotherapy. Neuropsychopharmacol 24, 478–491 (2001). https://doi.org/10.1016/S0893-133X(00)00223-2
Received:
Revised:
Accepted:
Issue Date:
DOI: https://doi.org/10.1016/S0893-133X(00)00223-2
Keywords
This article is cited by
-
Ethanol deprivation and central 5-HT deficiency differentially affect the mRNA editing of the 5-HT2C receptor in the mouse brain
Pharmacological Reports (2023)
-
Serotonin 5-HT2C Receptor Cys23Ser Single Nucleotide Polymorphism Associates with Receptor Function and Localization In Vitro
Scientific Reports (2019)
-
A-to-I RNA editing in the rat brain is age-dependent, region-specific and sensitive to environmental stress across generations
BMC Genomics (2018)
-
The role of 5-HT receptors in depression
Molecular Brain (2017)
-
ADAR RNA editing in human disease; more to it than meets the I
Human Genetics (2017)
