
Abstract
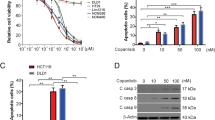
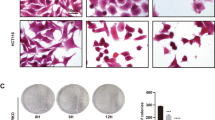
The tumor suppressor phosphatase and tensin homologue deleted from chromosome 10 (PTEN) gene is a negative regulator of the phosphatidylinositol-3-kinase (PI3K)/protein kinase B (Akt/PKB) signaling pathway. Overexpression of PTEN in cancer cells results in cell-cycle arrest and cell death through inhibition of PI3K. Caffeine, a xanthine analogue, is well known to enhance the cytocidal and growth-inhibitory effects of DNA-damaging agents such as radiation, UV light, and anticancer agents on tumor cells by abrogating DNA-damage checkpoints through inhibition of ataxia-telangiectasia-mutated (ATM), and ATM and Rad3-related (ATR) kinase activity. In this study, we demonstrate that treatment with a combination of adenovirus-mediated transfer of PTEN (Ad-PTEN) and caffeine synergistically suppressed cell growth and induced apoptosis in colorectal cancer cells but not in normal colorectal fibroblast cells. This synergistic effect was induced through abrogation of G2/M arrest, downregulation of the Akt pathway, and modulation of the p44/42MAPK pathway. Thus, combined treatment with Ad-PTEN and caffeine is a potential therapy for colorectal cancer.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others

References
Li J, Yen C, Liaw D, et al. PTEN, a putative protein tyrosine phosphatase gene mutated in human brain, breast, and prostate cancer. Science. 1997;275:1943–1947.
Cairns P, Okami K, Halachmi S, et al. Frequent inactivation of PTEN/MMAC1 in primary prostate cancer. Cancer Res. 1997;57:4997–5000.
Tashiro H, Blazes MS, Wu R, et al. Mutations in PTEN are frequent in endometrial carcinoma but rare in other common gynecological malignancies. Cancer Res. 1997;57:3935–3940.
Wang SI, Puc J, Li J, et al. Somatic mutations of PTEN in glioblastoma multiforme. Cancer Res. 1997;57:4183–4186.
Yokomizo A, Tindall DJ, Drabkin H, et al. PTEN/MMAC1 mutations identified in small cell, but not in non-small cell lung cancers. Oncogene. 1998;17:475–479.
Cheney W, Johnson D, Vaillancourt M-T, et al. Suppression of tumorigenicity of glioblastoma cells by adenovirus-mediated MMAC1/PTEN gene transfer. Cancer Res. 1998;58:2331–2334.
Davies MA, Lu Y, Sano T, et al. Adenoviral transgene expression of MMAC/PTEN in human glioma cells inhibits Akt activation and induces anoikis. Cancer Res. 1998;58:5285–5290.
Furnari FB, Huang HJ, Cavenee WK . The phosphoinositol phosphatase activity of PTEN mediates a serum-sensitive G1 growth arrest in glioma cells. Cancer Res. 1998;58:5002–5008.
Li DM, Sun H . PTEN/MMAC1/TEP1 suppresses the tumorigenicity and induces G1 cell cycle arrest in human glioblastoma cells. Proc Natl Acad Sci USA. 1998;95:15406–15411.
Minaguchi T, Mori T, Kanamori Y, et al. Growth suppression of human ovarian cancer cells by adenovirus-mediated transfer of the PTEN gene. Cancer Res. 1999;59:6063–6067.
Weng LP, Gimm O, Kum JB, et al. Transient ectopic expression of PTEN in thyroid cancer cell lines induces cell cycle arrest and cell type-dependent cell death. Hum Mol Genet. 2001;10:251–258.
Saito Y, Swanson X, Mhashilkar A, et al. Adenovirus-mediated transfer of the PTEN gene inhibits human colorectal cancer growth in vitro and in vivo. Gene Therapy. (in press).
Elledge SJ . Cell cycle checkpoints: preventing an identity crisis. Science. 1996;274:1664–1672.
O'Connor PM . Mammalian G1 and G2 phase checkpoints. Cancer Surv. 1997;29:151–182.
Hartwell LH, Kastan MB . Cell cycle control and cancer (review). Science. 1994;266:1821–1828.
IARC. IARC monograph evaluative carcinogen risks humans. 1991.
Mirvish SS, Cardesa A, Wallcave L, Shubik P . Induction of mouse lung adenomas by amines or ureas plus nitrite and by N-nitroso compounds: effect of ascorbate, gallic acid, thiocyanate, and caffeine. Nat Cancer Inst. 1975;55:633–636.
Nomura T . Diminution of tumorigenesis initiated by 4-nitroquinoline-1-oxide by post-treatment with caffeine in mice. Nature. 1976;260:547–549.
Nomura T . Comparative inhibiting effects of methylxanthines on urethane-induced tumors, malformations, and presumed somatic mutations in mice. Cancer Res. 1983;43:1342–1346.
Theiss JC, Shimkin MB . Inhibiting effect of caffeine on spontaneous and urethane-induced lung tumors in strain A mice. Cancer Res. 1978;38:1757–1761.
Welsch CW . Caffeine and the development of the normal and neoplastic mammary gland. Proc Soc Exp Biol Med. 1994;207:1–12.
Nishikawa A, Furukawa F, Imazawa T, Ikezaki S, Hasegawa T, Takahashi M . Effects of caffeine on glandular stomach carcinogenesis induced in rats by N-methyl-N′-nitro-N-nitrosoguanidine and sodium chloride. Food Chem Toxicol. 1995;33:21–26.
Nishikawa A, Furukawa F, Imazawa T, Yoshimura H, Mitsumori K, Takahashi M . Effects of caffeine, nicotine, ethanol and sodium selenite on pancreatic carcinogenesis in hamsters after initiation with N-nitrosobis(2-oxopropyl)amine. Carcinogenesis. 1992;13:1379–1382.
Huang MT, Xie JG, Wang ZY, et al. Effects of tea, decaffeinated tea, and caffeine on UVB light-induced complete carcinogenesis in SKH-1 mice: demonstration of caffeine as a biologically important constituent of tea. Cancer Res. 1997;57:2623–2629.
Zajdela F, Latarjet R . Inhibition of skin carcinogenesis in vivo by caffeine and other agents. Natl Cancer Inst Monogr. 1978;133–140.
Lau CC, Pardee AB . Mechanism by which caffeine potentiates lethality of nitrogen mustard. Proc Natl Acad Sci USA. 1982;79:2942–2946.
Matsuoka S, Huang M, Elledge SJ . Linkage of ATM to cell cycle regulation by the Chk2 protein kinase. Science. 1998;282:1893–1897.
Sarkaria JN, Busby EC, Tibbetts RS, et al. Inhibition of ATM and ATR kinase activities by the radiosensitizing agent, caffeine. Cancer Res. 1999;59:4375–4382.
Schlegel R, Pardee AB . Caffeine-induced uncoupling of mitosis from the completion of DNA replication in mammalian cells. Science. 1986;232:1264–1266.
Stewart AL, Mhashilkar AM, Yang XH, et al. PI3K blockade by Ad-PTEN inhibits invasion and induces apoptosis in radial growth phase and metastatic melanoma cells. Mol Med. 2002;8:451–461.
Roehm NW, Rodgers GH, Hatfield SM, Glasebrook AL . An improved colorimetric assay for cell proliferation and viability utilizing the tetrazolium salt XTT. J Immunol Methods. 1991;142:257–265.
Nishizaki M, Meyn RE, Levy LB, et al. Synergistic inhibition of human lung cancer cell growth by adenovirus-mediated wild-type p53 gene transfer in combination with docetaxel and radiation therapeutics in vitro and in vivo. Clin Cancer Res. 2001;7:2887–2897.
Taylor V, Wong M, Brandts C, et al. 5′ Phospholipid phosphatase SHIP-2 causes protein kinase B inactivation and cell cycle arrest in glioblastoma cells. Mol Cell Biol. 2000;20:6860–6871.
Gu J, Tamura M, Yamada KM . Tumor suppressor PTEN inhibits integrin- and growth factor-mediated mitogen-activated protein (MAP) kinase signaling pathways. J Cell Biol. 1998;143:1375–1383.
Tamura M, Gu J, Matsumoto K, Aota S, Parsons R, Yamada KM . Inhibition of cell migration, spreading, and focal adhesions by tumor suppressor PTEN. Science. 1998;280:1614–1617.
DeFrank JS, Tang W, Powell SN . p53-null cells are more sensitive to ultraviolet light only in the presence of caffeine. Cancer Res. 1996;56:5365–5368.
Powell SN, DeFrank JS, Connell P, et al. Differential sensitivity of p53(−) and p53(+) cells to caffeine-induced radiosensitization and override of G2 delay. Cancer Res. 1995;55:1643–1648.
Yao SL, Akhtar AJ, McKenna KA, et al. Selective radiosensitization of p53-deficient cells by caffeine-mediated activation of p34cdc2 kinase. Nat Med. 1996;2:1140–1143.
Cross SM, Sanchez CA, Morgan CA, et al. A p53-dependent mouse spindle checkpoint. Science. 1995;267:1353–1356.
Kuerbitz SJ, Plunkett BS, Walsh WV, Kastan MB . Wildtype p53 is a cell cycle checkpoint determinant following irradiation. Proc Natl Acad Sci USA. 1992;89:7491–7495.
Tsuiki H, Nitta M, Tada M, Inagaki M, Ushio Y, Saya H . Mechanism of hyperploid cell formation induced by microtubule-inhibiting drug in glioma cell lines. Oncogene. 2001;20:420–429.
Mayo LD, Dixon JE, Durden DL, Tonks NK, Donner DB . PTEN protects p53 from Mdm2 and sensitizes cancer cells to chemotherapy. J Biol Chem. 2002;277:5484–5489.
Tsuchiya H, Mori Y, Ueda Y, Okada G, Tomita K . Sensitization and caffeine potentiation of cisplatin cytotoxicity resulting from introduction of wild-type p53 gene in human osteosarcoma. Anticancer Res. 2000;20:235–242.
Dunphy WG, Kumagai A . The cdc25 protein contains an intrinsic phosphatase activity. Cell. 1991;67:189–196.
Kastan MB, Onyekwere O, Sidransky D, Vogelstein B, Craig R . Participation of p53 protein in the cellular response to DNA damage. Cancer Res. 1991;51:6304–6311.
Peng CY, Graves PR, Thoma RS, Wu Z, Shaw AS, Piwnica-Worms H . Mitotic and G2 checkpoint control: regulation of 14-3-3 protein binding by phosphorylation of Cdc25C on serine-216. Science. 1997;277:1501–1505.
Sebastian B, Kakizuka A, Hunter T . Cdc25M2 activation of cyclin-dependent kinases by dephosphorylation of threonine-14 and tyrosine-15. Proc Natl Acad Sci USA. 1993;90:3521–3524.
Thompson DA, Belinsky G, Chang TH, Jones DL, Schlegel R, Munger K . The human papillomavirus-16 E6 oncoprotein decreases the vigilance of mitotic checkpoints. Oncogene. 1997;15:3025–3035.
Taylor WR, Stark GR . Regulation of the G2/M transition by p53. Oncogene. 2001;20:1803–1815.
Tsuneoka M, Mekada E . Ras/MEK signaling suppresses Myc-dependent apoptosis in cells transformed by c-myc and activated ras. Oncogene. 2000;19:115–123.
Cross TG, Scheel-Toellner D, Henriquez NV, Deacon E, Salmon M, Lord JM . Serine/threonine protein kinases and apoptosis. Exp Cell Res. 2000;256:34–41.
Seger R, Krebs EG . The MAPK signaling cascade. FASEB J. 1995;9:726–735.
Acknowledgements
We thank Kathryn Hale for editorial assistance and Alma Vega for help in the preparation of this manuscript. This study was partially supported by the Texas Higher Education Coordinating Board ATP/ARP Grant 003657-0078-2001 (RR), by Public Health Service Grant PO1-CA 78778-01A1 (JAR), by a Career Development award from The University of Texas SPORE in Lung Cancer P50CA70907-5 (RR), by M.D. Anderson Cancer Center Institutional Research Grant (RR), by M.D. Anderson Cancer Center Support Grant CA16672, by the W.M. Keck Foundation Fund for Human Cancer Gene Prevention and Therapy (RR), by a BESCT Lung Cancer Program Grant (DAMD17-01-1-0689), by a TARGET Lung Cancer Grant (DAMD17-02-1-0706), and by a sponsored research agreement with Introgen Therapeutics, Inc.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Saito, Y., Gopalan, B., Mhashilkar, A. et al. Adenovirus-mediated PTEN treatment combined with caffeine produces a synergistic therapeutic effect in colorectal cancer cells. Cancer Gene Ther 10, 803–813 (2003). https://doi.org/10.1038/sj.cgt.7700644
Received:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.cgt.7700644
Keywords
This article is cited by
-
Fibroblast-derived CXCL12 regulates PTEN expression and is associated with the proliferation and invasion of colon cancer cells via PI3k/Akt signaling
Cell Communication and Signaling (2019)
-
Synthesis, anticancer, anti-HIV-1, and antimicrobial activity of some tricyclic triazino and triazolo[4,3-e]purine derivatives
Medicinal Chemistry Research (2012)
