
Abstract
In advanced breast cancers, TP53 mutation is highly predictive of complete response to high-dose epirubicin/cyclophosphamide chemotherapy. In these tumours with an altered control of genomic stability, accumulation of chemotherapy-induced genetic alterations may contribute to cell death and account for complete response. To explore the effects of chemotherapy on stability of the tumour genome, allelic profiles were obtained from microdissected tumour samples before and after chemotherapy in 29 unresponsive breast cancers (9 with TP53 mutation). Ninety-four per cent allelic profiles remained unchanged after treatment. Interestingly, 11 profiles (6%) showed important changes after treatment; allelic imbalances significantly increased (four cases) or decreased (seven cases) after chemotherapy in three distinct experiments, two of which using laser microdissected tumour cells. These genetic changes were not linked to the TP53 status, but one tumour showed complete disappearance of TP53-mutated cells in the residual tumour after treatment. Altogether, these observations carry important implications for the clonal evolution of breast cancers treated with DNA-damaging agents, as they point both to the importance of tumour heterogeneity and chemotherapy-driven selection of subclones.
Similar content being viewed by others


Main
Breast cancers are a heterogeneous group of tumours. Whereas most advanced breast cancer patients receive neoadjuvant chemotherapy, less than 20% of them will fully benefit from this type of treatment and reach complete pathological response, which is closely linked with longer survival (Bertheau et al, 2005; Kaufmann et al, 2005). The identification of predictive markers of response to chemotherapy is therefore a critical challenge for advanced breast cancer patients.
In previous studies (Bertheau et al, 2002, 2007), we have shown in non-inflammatory locally advanced breast cancers that TP53 mutations are highly predictive of complete pathological response to high-dose epirubicin/cyclophosphamide chemotherapy. TP53 is known to control genomic integrity (Vogelstein et al, 2000), and is induced by cytotoxic drugs (Lowe et al, 1993; Bunz et al, 1999), whereas its potential role as a prognostic and a predictive marker in cancer patients remains controversial (Vousden and Prives, 2005; Berns, 2006). We hypothesised that breast tumours with TP53 mutations could accumulate genetic alterations leading to complete tumour response through mitotic catastrophes and tumour cell death (Huang et al, 2005). Whereas obviously, we cannot explore the genome of tumours that underwent complete response, those that were not eradicated can be analysed. Note that we have previously shown that TP53 mutant tumours that did not undergo complete remissions have a worse prognosis than the other groups (Bertheau et al, 2007). This might be caused by chemotherapy-induced additional genetic changes that favour progression.
To test this hypothesis, we further explored the effects of chemotherapy on the tumour genome. Early tumour genetic changes after chemotherapy have been poorly studied in patients. We analysed here, in 29 locally advanced breast cancers, the allelic profiles obtained with ten polymorphic microsatellite markers known to be often involved in breast carcinoma (Bertheau et al, 2001). Allelic profiles obtained after chemotherapy were compared to those obtained before, using laser-microdissected surgical specimen.
Patients and methods
Patients and tumours
Among 80 non-inflammatory locally advanced breast cancers treated in a single institution from 1997 to 2001 with first-line chemotherapy and already studied for their TP53 status and response to treatment (Bertheau et al, 2007), we included 29 patients who fulfilled all the following criteria: patient informed of the study according to our Institutional Review Board recommendations; identical treatment (6 cycles of a dose–dense regimen associating 75 mg m−2 epirubicin and 1200 mg m−2 cyclophosphamide, given every 14 days with G-CSF support in case of febrile neutropenia; Cottu et al, 1999); and incomplete pathological response to treatment. The patients were women aged 23 to 87 (median age 43). Patients and tumours characteristics are given in Table 1. TNM tumour stages are given according to the WHO criteria (Sobin, 2002) and tumour grades are given according to the modified Scarff Bloom and Richardson system (Elston and Ellis, 1991). ESR1 and ERBB2 status were determined with immunohistochemistry (primary antibodies, respectively, clone 6F11 and clone CB11, Novocastra, Newcastle, UK). All patients underwent an initial open incisional biopsy, followed by chemotherapy and, three months after diagnosis, mastectomy and axillary lymph node dissection were performed. Only tumours with at least 5000 remaining cells on the mastectomy specimen tissue sections were included in the analysis. Three different categories of incomplete pathological response were individualised according to a methodology previously validated (Bertheau et al, 2005): tumours with post-treatment cellular density less than 50% compared to pre-treatment tumours defined ‘major pathological response’, tumours with post-treatment cellular density only slightly decreased, but with post-treatment cellular alterations defined ‘minor pathological response’, whereas tumours that remained unchanged after treatment defined ‘absence of response’.
TP53 typing
After tumour RNA extraction (Chomczynski and Sacchi, 1987), TP53 status was determined by the yeast functional assay (Flaman et al, 1995; Waridel et al, 1997). Tumours were considered TP53 mutant when (i) more than 15% of the yeast colonies were red and (ii) analysis using the split versions of the test could identify the defect in the 5′ or 3′ part of the gene, confirming the initial determination and (iii) sequence analysis from mutant yeast colonies could identify an unambiguous genetic defect (mutation, deletion, splicing defects…).
We determined the TP53 status with the yeast functional assay in all 29 tumours before and after chemotherapy, whenever sufficient frozen material was available for RNA extraction (18 tumours).
For the nine cases with TP53 mutation before treatment, frozen tumour tissue was available after treatment in six cases and used to perform the yeast functional test as well as TP53 gene sequencing. For the other three mutated cases, we tried to use formalin-fixed paraffin-embedded tissue, but failed to obtain good enough RNA to perform the yeast functional assay or long enough DNA fragments (>300 bp) to perform gene sequencing.
For all nine mutated cases, TP53 immunostainings on paraffin-embedded samples were obtained before and after treatment using DO-7 monoclonal mouse anti-human antibody (Dako, Trappes, France), diluted 1/50 in an automated immunostainer (Ventana Medical Systems SA, Illkirch, France). The percentage of stained cells (nuclear staining) was noted.
Laser-microdissection and pressure catapulting (LMPC)
Using a PALM Microbeam/Olympus system, LMPC was performed on all 29 tumours before and after chemotherapy: 7 μm thick paraffin sections were spread on membrane-coated slides and stained with H&E (Figure 1A). A pulsed UV-A nitrogen laser (337 nm) was used to cut and catapult small tissue fragments directly into the buffer-containing cap of a microfuge tube. For each tumour, at least 5000 cell sections obtained in several tumour areas were lysed in 30 μl of buffer (50 mM Tris-HCl (pH 7.5), 1 mM EDTA, 0.5% Tween 20, 0.2 mg ml−1 proteinase K). After an incubation of 24–48 h at 56°C, proteinase K was inactivated at 95°C for 10 min. No further DNA extraction was performed before PCR analysis for these microdissected samples (Bertheau et al, 2001).

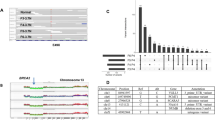
(A) Laser-microdissection of breast carcinoma paraffin sections stained with H&E. The left panels show the selected areas (surrounded by a black line) before microdissection and the right panels show the same areas after microdissection. Tumour cell density is much lower after chemotherapy (lower panels). (B) Examples of unchanged profiles in two patients: retention of heterozygosity (left panels) or loss of heterozygosity (right panels) before and after chemotherapy. (C and D) Examples of decreased AI in two patients. (C) For patient 7 marker D8S261, LOH is present before chemotherapy with complete allelic loss on microdissected tumour (left panel) and important AI on not microdissected tumour (right panel). After chemotherapy, AI is strongly reduced with both methods (lower panels). (D) For patient 3 marker D7S490, LOH is present before chemotherapy on microdissected tumour (left upper panel) and LOH has disappeared after chemotherapy (left lower panel). Use of not microdissected tissue is not conclusive since tumour cells are contaminated with numerous inflammatory cells (inset: tumour cells (T), inflammatory cells (Inf), necrosis (Nec)). (E and F) Examples of increased AI in two patients. (E) For patient 32 marker D11S1356, there is no AI before chemotherapy with both microdissected and not microdissected tumours (upper panels), whereas complete LOH is present after chemotherapy with microdissected tumour (lower left panel). Use of whole tumour tissue after chemotherapy (lower right panel) is not conclusive (does not reach -50%), as tumour cells are contaminated with inflammatory cells (inset: tumour cells (T), inflammatory cells (Inf)). (F) For patient 39 marker D16S496, LOH is present on the short allele before chemotherapy and on the long allele after chemotherapy with microdissected tumour (left panels). Profiles with whole tumour tissue (right panels) are close. For all allelic profiles, normal DNA profiles are shown in black (alleles N1 and N2), tumour DNA profiles are shown in colour (blue or green) (alleles T1 and T2). x-axis shows allelic size in base pairs, y-axis shows fluorescence intensity.
DNA extraction
Control allelic profiles were obtained with DNA extracted from blood cells: after cell lysis and proteinase K degradation, DNA was obtained with phenol chloroform extraction followed by DNA precipitation with ethanol and solved in TE buffer (Tris 10 mM (pH 7.5), EDTA 1 mM) (Muniz et al, 1994).
Tumours that showed post-treatment allelic changes with microdissected samples (see Results) were also analysed with DNA extracted from frozen whole tumour tissue. Briefly, frozen tumour sections were immersed in a buffer containing 8 M urea, 0.3 M NaCl, 10 mM EDTA, 2% SDS and 10 mM Tris-HCL, pH 7.5, and then submitted to phenol chloroform DNA extraction.
PCR
For microdissected cells, a volume of lysis buffer accounting for 500 cells was added in each PCR vial. For DNA extracted from blood cells, 10 ng DNA was used for each PCR. The following 10 microsatellite dinucleotide repeats were used (Table 2): D3S1573; D7S490; D8S1820; D8S261; D11S860; D11S1356; D13S171; D16S496; IGP53; and D17S855. The PCR mix contained 1 U Taq Gold (Applied Biosystems, Foster City, CA, USA), 2.5–4 mM MgCl2, 0.2 mM dNTP, 0.2 μ M labelled forward primers (NED™ for normal, FAM (6-carboxyfluorescein) or VIC™ for tumour) and 0.2 μ M non-labelled reverse primers. The PCR final volume was 20 μl. Thirty-five cycles of PCR were performed.
After denaturation, the PCR products were run on ABI PRISM 310 Genetic Analyzer. The analysis of the migration data were performed with Genescan 3.1 software (Applied Biosystems).
Allelic profiles analysis
Fluorescent allelic ratios obtained from microdissected tumour tissue were compared with fluorescent allelic ratios obtained from control DNA, allowing the measurement of allelic imbalances (AI). Loss of heterozygosity (LOH) was defined as AI greater than 50% for one tumour allele.
AI observed after chemotherapy were then compared to AI observed before chemotherapy, showing locus with unchanged AI, decreased AI or increased AI. Decreased or increased AI were considered significant only when allelic peak heights differed by at least 50%. This threshold was chosen to minimize the risk of false-positive results induced by PCR variations. All these significant pre-post-treatment differences were verified with a second round of PCR performed with microdissected tissue and a third round of PCR performed with frozen whole tumour DNA.
Results
Allelic profiles before chemotherapy
Among a total of 290 ‘before chemotherapy’ PCR (10 markers performed in 29 tumours), 63 allelic profiles were homozygote (non-informative) and 15 PCR could not be analysed due to technical issues. The rates of informativity for these markers were similar to those expected from databases (Tables 2 and 3). The 212 informative profiles revealed 87 (41%) LOH and 125 (59%) retentions of heterozygosity. Rates of LOH at each locus were similar to those observed in the literature in the corresponding chromosomal regions (Kerangueven et al, 1997; Braga et al, 1999; Katsama et al, 2000; Osborne and Hamshere, 2000; Shen et al, 2000; Hirano et al, 2001; Miyakis and Spandidos, 2002; Wang et al, 2004; Mao et al, 2005).
Allelic profiles after chemotherapy
Among a total of 290 ‘after chemotherapy’ PCR (29 tumours, 10 markers), 63 allelic profiles were homozygote (non-informative) and 24 PCR could not be analysed due to technical issues. The 203 informative profiles revealed 78 (38%) LOH and 125 (62%) retentions of heterozygosity. (Tables 2 and 3)
Comparison of allelic profiles before and after chemotherapy
Altogether, 191 pairs of PCR were informative and analysable before and after chemotherapy. In 180 pairs (94%), allelic profiles before and after chemotherapy were not different (Figure 1B; Table 3, green cells). However, in 11 pairs, pre- and post-chemotherapy profiles were significantly different. In seven pairs (4%), AI observed before chemotherapy had significantly decreased after chemotherapy (Figure 1C and D; Table 3, blue cells). In two pairs (1%), almost normal profiles observed before chemotherapy were clearly imbalanced after chemotherapy (Figure 1E; Table 3, orange cells). In 2 pairs (1%), AI observed for one allele before chemotherapy was found for the other allele after chemotherapy (Figure 1F; Table 3, orange* cells).
For these 11 important changes detected after chemotherapy, we reproduced once the analysis with microdissected samples. A further analysis with non-microdissected extracted DNA showed very close results (Figure 1C and F) in five pairs, and non-conclusive results (Figure 1D and E) in the other six pairs.
Allelic profiles and TP53 status
The overall rates of LOH at all loci were similar in tumours with or without TP53 mutations, before (47 and 38%, respectively) or after treatment (41 and 37%, respectively).
LOH at the TP53 intragenic IGP53 locus before treatment was observed in 4 out of these 9 tumours with TP53 mutation (44%) and in 5 out of 20 tumours without TP53 mutation (20%). Eighteen tumours had sufficient post-treatment frozen material to allow analysis with the yeast functional assay after chemotherapy. Among them, all 12 tumours that did not bear TP53 mutation before treatment did not show post-treatment TP53 mutation.
Six out of nine tumours with TP53 mutation before treatment had good quality post-treatment tissue available and have been further analysed for post-treatment TP53 status (Table 4). Among these six tumours, two still bore a mutation after treatment (patients 3 and 12), whereas no mutation could be detected by the functional test in four others patients (patients 7, 8, 9 and 11). For these four patients with absence of detectable TP53-mutated cells after treatment, note that three of them had either a dense inflammatory stroma (patients 9 and 11) or a single small (2 mm) residual focus of tumour cells (patient 8).
Allelic profiles and clinicopathological data
The distribution of the 11 post-chemotherapy profile changes was not linked to any other clinicopathological parameter (age, tumour grade, tumour stage, ESR1 status, ERBB2 status and type of incomplete pathological response) and was not linked to the pre-treatment TP53 status of the tumours.
Discussion
We found that genetic changes are detectable early after chemotherapy in breast cancers.
All 29 patients had poor prognosis breast tumours treated with the same dose–dense epirubicin/cyclophosphamide regimen. The large amount of tumour tissue obtained before (open incisional biopsy) and after (mastectomy) treatment allowed reliable assessment of response to treatment. Obtaining such homogeneous pre- and post-chemotherapy tissue samples is difficult and explains why we could study only 29 patients.
Advanced breast cancers are often high grade with necrotic areas, and become more heterogeneous after treatment, with inflammatory infiltrate, oedema, fibrosis and necrosis. To overcome tissue heterogeneity, we used laser tissue microdissection, which allows a targeted sampling of tumour cells with very little contamination. In this regard, 20/29 tumours in our study had at least one allelic loss greater than 80%, indicating that tumour tissue samples were made of almost pure tumour cells. Although laser-microdissection is a highly efficient sampling method, it yields limited amounts of tissue material, and we took special care of sampling at least 5000 cells for each tumour. Indeed, allelic profiling must take into account allelic dropout artefacts that are artificial allelic imbalances occurring when DNA concentration is too low (Coulet et al, 2000; Miller et al, 2002). Several tumour areas were sampled in each tumour to obtain a full representativity of the tumour cell population and to avoid the sampling of genetically different subclones. In addition, all PCR showing important differences between pre- and post-treatment allelic profiles were confirmed with a second round of PCR using microdissected material and a third PCR with non-microdissected tumour tissue.
The chemotherapeutic regimen used in this study contained epirubicin and cyclophosphamide that are both well known for their cytotoxic and mutagenic effects in vitro. Anthracyclins are topoisomerase II inhibitors that induce DNA double-strand breaks (Ferguson and Baguley, 1996). In animal or human cell cultures, anthracyclins induce chromosomal alterations rather than mutations (El-Mahdy Sayed Othman, 2000; Baumgartner et al, 2004), often through homologous recombination mechanisms (Lehmann et al, 2003). The altered cells then either engage their apoptotic programme or accumulate genetic alterations (Morris et al, 1995). Alkylating agents transfer alkyl groups to cellular constituents, inducing DNA adducts as well as DNA crosslinking that may lead to point mutations (Sanderson and Shield, 1996; Hunter et al, 2006). A recent study using microarray-based CGH on 21 breast tumours before and after epirubicin/cyclophosphamide chemotherapy showed a significant acquired copy number gain on 11p15.2–11p15.5 after treatment (Pierga et al, 2007). In 30 patients with breast cancers treated with alkylating agents, microsatellite instability and LOH were found in peripheral blood mononuclear cells, respectively, in 27 and in 6 patients (Fonseca et al, 2005), illustrating how acute chemotherapy-induced DNA damage can also affect normal cells.
In our study, at least two different mechanisms could explain the increase of AI at 11p15 and 11q23.3 after chemotherapy: a pre-existent undetectable small population of tumour cells bearing one AI could have been selected by the treatment that would have preferentially targeted the tumour cells without AI. Alternatively, new AI could be directly induced by the treatment in a sufficient number of tumour cells to become detectable through molecular analyses. In this regard, changes induced by anthracyclins at the 11q23 MLL locus lead to therapy-related acute leukaemias (Allan and Travis, 2005). We also observed in two tumours, at D13S171 (13q13.1) (patient 6) and D16S496 (16q22.1) (patient 39), imbalance on one allele before chemotherapy followed by imbalance on the other allele after chemotherapy. These ‘inversed profiles’ could be explained by selection of a minor subclone bearing one allelic loss through a preferential effect of the drug on a major subclone bearing loss of the other allele. In seven PCR (six tumours, five markers: D7S490 (7q31.32), D8S261 (8p22), D11S1356 (11q23.3), D13S171 (13q13.1) and IGP53 (17p13.1), AI present before treatment strongly decreased after treatment, leading to almost normal allelic profiles in five cases. Contamination by non-tumour cells may hide AI in tumour cells, but we purposely used laser microdissection to prevent this bias. The most likely explanation for these ‘improvements’ is chemotherapy-driven selection of tumour subclones without AI.
One purpose of our study was to determine whether post-chemotherapy genetic abnormalities could occur at different rates in TP53-mutated versus TP53-nonmutated breast tumours. TP53 mutations are observed in 20 to 30% of breast carcinomas and are linked to poor prognosis (Turpin et al, 1999; Olivier et al, 2006). Loss of TP53 function may result in increased genetic instability with higher frequency of mutations, chromosomal abnormalities, gene amplifications, LOH and abnormal chromosome segregation (Fukasawa et al, 1996; Morris, 2002). Our study failed to demonstrate strong link between the pre-treatment TP53 status of the tumours and the occurrence of post-treatment genetic events. Further investigations using xenografted human breast tumours would allow sequential analyses of the genetic alterations during chemotherapy-induced tumour regression. Yet in four cases (patients 7, 8, 9 and 11), the TP53 mutation found before chemotherapy was not found after (Table 4). This could indicate, as in our previous observations, that the TP53-mutated cells were targeted by the chemotherapy, especially in one case (patient 7) that showed a low post-treatment stromal cell density as well as an important decrease of the proportion of cells with TP53 LOH (Table 3). The other three post-treatment tumours (patients 9, 11 and 8) had either a high stromal cell density (patients 9 and 11) or consisted of a single small residual nodule (patient 8) that might have impeded detection of the TP53 mutation in these non-microdissected frozen samples. Immunohistochemistry showed either negative stainings or a trend to a decrease in the proportion of TP53-overexpressing cells after treatment, pointing to higher chemosensitivity of TP53-mutated cells, even if TP53 immunohistochemistry is a less reliable indicator of the TP53 gene status than the yeast functional assay.
In this study, we demonstrate that genetic changes do occur early after chemotherapy in breast carcinomas. The most likely explanation for our observations is chemotherapy-driven selection of tumour subclones: breast cancer tumour cells are well known to be highly heterogeneous (Shipitsin et al, 2007) and those already bearing genetic alterations may be more sensitive to chemotherapy and enter apoptosis more rapidly than unaltered cells. Another recent report also showed that such post-treatment changes do exist but, as in our study, seem to be rare (Pierga et al, 2007). Whether specific molecular abnormalities can facilitate or impede response to therapy is a crucial issue in oncology (Tanner et al, 2006). However, exquisite chemosensitivity linked with specific allelic losses in tumours has not been reported yet. The disappearance of TP53-mutated cells after chemotherapy in one case (patient 7) is clearly in favour of clonal selection. Yet, whatever the mechanism, these treatment-induced events should favour the emergence of therapy-resistant tumour subclones and subsequent tumour recurrences.
Change history
16 November 2011
This paper was modified 12 months after initial publication to switch to Creative Commons licence terms, as noted at publication
References
Allan JM, Travis LB (2005) Mechanisms of therapy-related carcinogenesis. Nat Rev Cancer 5: 943–955
Baumgartner A, Schmid TE, Cemeli E, Anderson D (2004) Parallel evaluation of doxorubicin-induced genetic damage in human lymphocytes and sperm using the comet assay and spectral karyotyping. Mutagenesis 19: 313–318
Berns A (2006) Cancer biology: can less be more for p53? Nature 443: 153–154
Bertheau P, Lerebours F, Mounier N, de Roquancourt A, Espie M, Clot P, Servant JM, Misset JL, Marty M, Janin A (2005) Prognostic significance of a combined clinicopathologic score for response to primary systemic therapy in locally advanced breast cancer. Oncol Rep 14: 513–520
Bertheau P, Plassa F, Espie M, Turpin E, de Roquancourt A, Marty M, Lerebours F, Beuzard Y, Janin A, de The H (2002) Effect of mutated TP53 on response of advanced breast cancers to high-dose chemotherapy. Lancet 360: 852–854
Bertheau P, Plassa LF, Lerebours F, de Roquancourt A, Turpin E, Lidereau R, de The H, Janin A (2001) Allelic loss detection in inflammatory breast cancer: improvement with laser microdissection. Lab Invest 81: 1397–1402
Bertheau P, Turpin E, Rickman D, Espié M, de Reyniès A, Feugeas J, Plassa L, Soliman H, Varna M, de Roquancourt A, Lehmann-Che J, Beuzard Y, Marty M, Misset J, Janin A, de Thé H (2007) Exquisite sensitivity of TP53 mutant and basal breast cancers to a dose-dense epirubicin/cyclophosphamide chemotherapy regimen. PLoSMed 4: e90
Braga E, Pugacheva E, Bazov I, Ermilova V, Kazubskaya T, Mazurenko N, Kisseljov F, Liu J, Garkavtseva R, Zabarovsky E, Kisselev L (1999) Comparative allelotyping of the short arm of human chromosome 3 in epithelial tumors of four different types. FEBS Lett 454: 215–219
Bunz F, Hwang PM, Torrance C, Waldman T, Zhang Y, Dillehay L, Williams J, Lengauer C, Kinzler KW, Vogelstein B (1999) Disruption of p53 in human cancer cells alters the responses to therapeutic agents. J Clin Invest 104: 263–269
Chomczynski P, Sacchi N (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 162: 156–159
Cottu PH, Zelek L, Extra JM, Espie M, Mignot L, Morvan F, Marty M (1999) High-dose epirubicin and cyclophosphamide every two weeks as first-line chemotherapy for relapsing metastatic breast cancer patients. Ann Oncol 10: 795–801
Coulet F, Blons H, Cabelguenne A, Lecomte T, Lacourreye O, Brasnu D, Beaune P, Zucman J, Laurent-Puig P (2000) Detection of plasma tumor DNA in head and neck squamous cell carcinoma by microsatellite typing and p53 mutation analysis. Cancer Res 60: 707–711
El-Mahdy Sayed Othman O (2000) Cytogenetic effect of the anticancer drug epirubicin on Chinese hamster cell line in vitro. Mutat Res 468: 109–115
Elston CW, Ellis IO (1991) Pathological prognostic factors in breast cancer. I. The value of histological grade in breast cancer: experience from a large study with long-term follow-up. Histopathology 19: 403–410
Ferguson LR, Baguley BC (1996) Mutagenicity of anticancer drugs that inhibit topoisomerase enzymes. Mutat Res 355: 91–101
Flaman JM, Frebourg T, Moreau V, Charbonnier F, Martin C, Chappuis P, Sappino AP, Limacher IM, Bron L, Benhattar J, Tada M, Van Meir EG, Estreicheru A, Iggo RD (1995) A simple p53 functional assay for screening cell lines, blood, and tumors. Proc Natl Acad Sci USA 92: 3963–3967
Fonseca FL, Sant Ana AV, Bendit I, Arias V, Costa LJ, Pinhal AA, del Giglio A (2005) Systemic chemotherapy induces microsatellite instability in the peripheral blood mononuclear cells of breast cancer patients. Breast Cancer Res 7: R28–R32
Fukasawa K, Choi T, Kuriyama R, Rulong S, Vande Woude GF (1996) Abnormal centrosome amplification in the absence of p53. Science 271: 1744–1747
Hirano A, Emi M, Tsuneizumi M, Utada Y, Yoshimoto M, Kasumi F, Akiyama F, Sakamoto G, Haga S, Kajiwara T, Nakamura Y (2001) Allelic losses of loci at 3p25.1, 8p22, 13q12, 17p13.3, and 22q13 correlate with postoperative recurrence in breast cancer. Clin Cancer Res 7: 876–882
Huang X, Tran T, Zhang L, Hatcher R, Zhang P (2005) DNA damage-induced mitotic catastrophe is mediated by the Chk1-dependent mitotic exit DNA damage checkpoint. Proc Natl Acad Sci USA 102: 1065–1070
Hunter C, Smith R, Cahill DP, Stephens P, Stevens C, Teague J, Greenman C, Edkins S, Bignell G, Davies H, O'Meara S, Parker A, Avis T, Barthorpe S, Brackenbury L, Buck G, Butler A, Clements J, Cole J, Dicks E, Forbes S, Gorton M, Gray K, Halliday K, Harrison R, Hills K, Hinton J, Jenkinson A, Jones D, Kosmidou V, Laman R, Lugg R, Menzies A, Perry J, Petty R, Raine K, Richardson D, Shepherd R, Small A, Solomon H, Tofts C, Varian J, West S, Widaa S, Yates A, Easton DF, Riggins G, Roy JE, Levine KK, Mueller W, Batchelor TT, Louis DN, Stratton MR, Futreal PA, Wooster R (2006) A hypermutation phenotype and somatic MSH6 mutations in recurrent human malignant gliomas after alkylator chemotherapy. Cancer Res 66: 3987–3991
Katsama A, Sourvinos G, Zachos G, Spandidos DA (2000) Allelic loss at the BRCA1, BRCA2 and TP53 loci in human sporadic breast carcinoma. Cancer Lett 150: 165–170
Kaufmann M, von Minckwitz G, Rody A (2005) Preoperative (neoadjuvant) systemic treatment of breast cancer. Breast 14: 576–581
Kerangueven F, Noguchi T, Coulier F, Allione F, Wargniez V, Simony-Lafontaine J, Longy M, Jacquemier J, Sobol H, Eisinger F, Birnbaum D (1997) Genome-wide search for loss of heterozygosity shows extensive genetic diversity of human breast carcinomas. Cancer Res 57: 5469–5474
Lehmann M, Franco A, de Souza Prudente Vilar K, Lukza Reguly M, de Andrade HH (2003) Doxorubicin and two of its analogues are preferential inducers of homologous recombination compared with mutational events in somatic cells of Drosophila melanogaster. Mutat Res 539: 167–175
Lowe SW, Schmitt EM, Smith SW, Osborne BA, Jacks T (1993) p53 is required for radiation-induced apoptosis in mouse thymocytes. Nature 362: 847–849
Mao X, Hamoudi RA, Zhao P, Baudis M (2005) Genetic losses in breast cancer: toward an integrated molecular cytogenetic map. Cancer Genet Cytogenet 160: 141–151
Miller CR, Joyce P, Waits LP (2002) Assessing allelic dropout and genotype reliability using maximum likelihood. Genetics 160: 357–366
Miyakis S, Spandidos DA (2002) Allelic loss in breast cancer. Cancer Detect Prev 26: 426–434
Morris SM (2002) A role for p53 in the frequency and mechanism of mutation. Mutat Res 511: 45–62
Morris SM, Domon OE, McGarrity LJ, Chen JJ, Casciano DA (1995) Programmed cell death and mutation induction in AHH-1 human lymphoblastoid cells exposed to m-amsa. Mutat Res 329: 79–96
Muniz ES, Plassa F, Amselem S, Goossens M, Vernant JP (1994) Molecular analysis of polymorphic loci to study chimerism after allogeneic bone marrow transplantation. Heteroduplex analysis in denaturing gradient gel electrophoresis: a new approach to detecting residual host cells. Transplantation 57: 451–456
Olivier M, Langerod A, Carrieri P, Bergh J, Klaar S, Eyfjord J, Theillet C, Rodriguez C, Lidereau R, Bieche I, Varley J, Bignon Y, Uhrhammer N, Winqvist R, Jukkola-Vuorinen A, Niederacher D, Kato S, Ishioka C, Hainaut P, Borresen-Dale AL (2006) The clinical value of somatic TP53 gene mutations in 1794 patients with breast cancer. Clin Cancer Res 12: 1157–1167
Osborne RJ, Hamshere MG (2000) A genome-wide map showing common regions of loss of heterozygosity/allelic imbalance in breast cancer. Cancer Res 60: 3706–3712
Pierga JY, Reis-Filho JS, Cleator SJ, Dexter T, Mackay A, Simpson P, Fenwick K, Iravani M, Salter J, Hills M, Jones C, Ashworth A, Smith IE, Powles T, Dowsett M (2007) Microarray-based comparative genomic hybridisation of breast cancer patients receiving neoadjuvant chemotherapy. Br J Cancer 96: 341–351
Sanderson BJ, Shield AJ (1996) Mutagenic damage to mammalian cells by therapeutic alkylating agents. Mutat Res 355: 41–57
Shen CY, Yu JC, Lo YL, Kuo CH, Yue CT, Jou YS, Huang CS, Lung JC, Wu CW (2000) Genome-wide search for loss of heterozygosity using laser capture microdissected tissue of breast carcinoma: an implication for mutator phenotype and breast cancer pathogenesis. Cancer Res 60: 3884–3892
Shipitsin M, Campbell LL, Argani P, Weremowicz S, Bloushtain-Qimron N, Yao J, Nikolskaya T, Serebryiskaya T, Beroukhim R, Hu M, Halushka MK, Sukumar S, Parker LM, Anderson KS, Harris LN, Garber JE, Richardson AL, Schnitt SJ, Nikolsky Y, Gelman RS, Polyak K (2007) Molecular definition of breast tumor heterogeneity. Cancer Cell 11: 259–273
Sobin LH, Wittekind Ch (eds) (2002) TNM Classification of Malignant Tumors. John Wiley & Sons: New York, pp 145–156
Tanner M, Isola J, Wiklund T, Erikstein B, Kellokumpu-Lehtinen P, Malmstrom P, Wilking N, Nilsson J, Bergh J (2006) Topoisomerase IIalpha gene amplification predicts favorable treatment response to tailored and dose-escalated anthracycline-based adjuvant chemotherapy in HER-2/neu-amplified breast cancer: Scandinavian Breast Group Trial 9401. J Clin Oncol 24: 2428–2436
Turpin E, Dalle B, de Roquancourt A, Plassa LF, Marty M, Janin A, Beuzard Y, de The H (1999) Stress-induced aberrant splicing of TSG101: association to high tumor grade and p53 status in breast cancers. Oncogene 18: 7834–7837
Vogelstein B, Lane D, Levine AJ (2000) Surfing the p53 network. Nature 408: 307–310
Vousden KH, Prives C (2005) P53 and prognosis: new insights and further complexity. Cell 120: 7–10
Wang ZC, Lin M, Wei LJ, Li C, Miron A, Lodeiro G, Harris L, Ramaswamy S, Tanenbaum DM, Meyerson M, Iglehart JD, Richardson A (2004) Loss of heterozygosity and its correlation with expression profiles in subclasses of invasive breast cancers. Cancer Res 64: 64–71
Waridel F, Estreicher A, Bron L, Flaman JM, Fontolliet C, Monnier P, Frebourg T, Iggo R (1997) Field cancerisation and polyclonal p53 mutation in the upper aero-digestive tract. Oncogene 14: 163–169
Acknowledgements
We thank Christophe Leboeuf, Dominique Cuvillier, Evelyne Wittmer and Claire Bocquet for their technical assistance and Jessica Zucman-Rossi for her helpful advice. This work was supported by PHRC, ARC and ANRT.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
From twelve months after its original publication, this work is licensed under the Creative Commons Attribution-NonCommercial-Share Alike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Varna, M., Soliman, H., Feugeas, JP. et al. Changes in allelic imbalances in locally advanced breast cancers after chemotherapy. Br J Cancer 97, 1157–1164 (2007). https://doi.org/10.1038/sj.bjc.6603937
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.bjc.6603937
Keywords
This article is cited by
-
Clonal evolution in primary breast cancers under sequential epirubicin and docetaxel monotherapy
Genome Medicine (2022)
-
Immunohistochemical and molecular analyses of HER2 status in breast cancers are highly concordant and complementary approaches
British Journal of Cancer (2011)
-
A case of HER2-positive male breast cancer with lung metastases showing a good response to trastuzumab and paclitaxel treatment
Breast Cancer (2009)
