
Abstract
Germline mutations in genes coding for molecules involved in the RAS/RAF/MEK/ERK pathway are the hallmarks of a newly classified family of autosomal dominant syndromes termed RASopathies. Myeloproliferative disorders (MPDs), in particular, juvenile myelomonocytic leukemia, can lead to potentially severe complications in children with Noonan syndrome (NS). We studied 27 children with NS or other RASopathies and 35 age-matched children as control subjects. Peripheral blood (PB) cells from these patients were studied for in vitro colony-forming units (CFUs) activity, as well as for intracellular phosphosignaling. Higher spontaneous growth of both burst-forming units-erythroid (BFU-E) and CFU-granulocyte/macrophage (CFU-GM) colonies from RAS-mutated patients were observed as compared with control subjects. We also observed a significantly higher amount of GM-colony-stimulating factor-induced p-ERK in children with RASopathies. Our findings demonstrate for the first time that PB cells isolated from children suffering from NS or other RASopathies without MPD display enhanced BFU-E and CFU-GM colony formation in vitro. The biological significance of these findings clearly awaits further studies. Collectively, our data provide a basis for further investigating of only partially characterized hematological alterations present in children suffering from RASopathies, and may provide new markers for progression toward malignant MPD in these patients.
Similar content being viewed by others

Introduction
The presence of germline mutations in genes coding for molecules involved in the RAS/RAF/MEK/ERK mitogen-activating protein kinase (MAPK) cascade is the molecular basis for a newly classified family of autosomal dominant syndromes termed ‘RASopathies’. These disorders that include Noonan syndrome (NS), LEOPARD syndrome, Costello syndrome, cardiofaciocutaneous syndrome, neurofibromatosis 1 and Legius syndrome are developmental syndromes affecting ~1 in 1000 live birth.1, 2, 3 Each of them exhibits distinctive phenotypic features, although there are numerous overlapping clinical manifestations including dysmorphic craniofacial features, congenital cardiac defects, skin, skeletal and ocular abnormalities, varying degrees of intellectual disability and increased cancer risk.4, 5, 6 NS, the most common among these disorders, is characterized by heterozygous germline mutation in the PTPN11 gene in approximately half of the cases.7 The PTPN11 proto-oncogene encodes Src-homology tyrosine phosphatase-2 (SHP-2), a protein tyrosine phosphatase with a role in signal transduction and hematopoiesis.8
Progression toward myeloproliferative and myelodysplastic disorders, which can cause severe medical problems, may occur in NS patients, although spontaneous remission has often been described.9, 10, 11 Juvenile myelomonocytic leukemia (JMML) and JMML-like myeloproliferative disorder may also occur in some NS patients with favorable outcome.12 JMML is an aggressive myeloproliferative neoplasm of childhood characterized by uncontrolled proliferation of monocytic and granulocytic cells. JMML-derived hematopoietic precursor cells often show hypersensitivity to granulocyte-macrophage colony-stimulating factor (GM-CSF) in vitro.13, 14 PTPN11 somatic missense mutations have been identified in approximately one-third of JMML cases. In this regard, analysis of the mutational spectra observed in NS vs JMML patients clearly indicates that germline PTPN11 NS-associated mutations have only a mild effect on development and hematopoiesis when compared with JMML-associated somatic PTPN11 lesions.15, 16, 17, 18, 19
Although the molecular mechanism underlying the progression of hematopoietic disorders observed in patients bearing PTPN11 mutations is not fully understood, several in vitro and in vivo studies indicate that hyperactivation of the tyrosine phosphatase SHP-2, encoded by PTPN11, may have a role in aberrant activation of both the JAK/STAT and RAS/MAPK/ERK signaling pathways favoring the pathogenesis of NS and JMML.20, 21, 22
On the basis of these findings, here we have systematically evaluated the extent of RAS/MAPK signaling activation in a cohort of RAS-mutated patients in order to explore the effect of aberrant RAS activation in circulating progenitors and mature monocytes, and possibly to determine genotype–phenotype correlations.
Materials and Methods
Patients and samples
In this study, a total of 27 patients, admitted to Pediatrics Clinical Genetics of San Gerardo Hospital, were enrolled with a clinical diagnosis of NS or related disorder (hereinafter referred to as RAS-mutated patients). As control cohort, we examined 35 age-matched children, which included both patients without onco-hematological diseases and patients with others genetic defects that could not affect the JAK/STAT and RAS/RAF/ERK pathways. Peripheral blood (PB) cells (n=22) were cultured in vitro in order to evaluate both spontaneous and growth factor-stimulated colony-forming unit (CFU) activity. In addition, PB monocytes were assessed for IL (interleukin)-6 (50 ng/ml)-induced response of p-STAT3 (n=15), GM-CSF (0.1 and 10 ng/ml)-induced response of p-STAT5 (n=15) and p-ERK (n=16) by phosphoflow cytometry, as previously described.23 Clinical and biological characteristics of the patients enrolled in this study are shown in Table 1. This study was approved by the local institutional ethics committees and carried out with the informed consent of the patients’ guardians.
Cytokine stimulation and phospho-specific flow cytometry
Freshly isolated PB mononuclear cells were suspended in serum-free media ex vivo at a concentration of 1–2 × 106 cells per ml, rested for 1 h at 37 °C, and assessed for total cell count and viability by trypan blue dye exclusion before being subject to phosphoflow assay. Samples were then stimulated with 0.1 and 10 ng/ml of GM-CSF or 50 ng/ml of IL-6 for 10 and 15 min, respectively, to allow signal transduction. Cells were then fixed with paraformaldehyde and permeabilized with ice-cold methanol according to published methods23 and then incubated with anti-phosphoprotein-directed monoclonal antibodies, or isotype-matched IgG, and surface antigen-directed monoclonal antibodies, previously tested for their maintenance of expression after fixation and permeabilization.
Samples were stained with antibodies such as CD33 PE (clone P67.6), CD14 APC-H7 (clone MϕP9), CD45 PerCP (clone 2D1), p-STAT5 Alexa 647 (pY694), p-STAT3 Alexa 488 (pY705), p-ERK1/2 Alexa 488 (T202/Y204) or isotypes IgG1k Alexa 488 and Alexa 647 (clone MOPC-21) (BD Bioscience, San José, CA, USA), and data were acquired on a FACSaria flow cytometer (BD Bioscience). At least 100 000 events were collected and analyzed using DIVA software (San José, CA, USA). Basal levels of each phosphoprotein were calculated as percentage of phospho-positive cells in unstimulated conditions. Response to stimulation upon cytokine treatment (for example, recombinant human GM-CSF or IL-6) was calculated by subtracting the percentage of unstimulated phospho-positive cells under basal conditions. Each phosphoprotein reactivity was evaluated in CD14+/CD33+/CD45+ cells as assessed by the flow cytometric gating strategy described in Figure 1. GM-CSF-induced p-STAT5 signaling in monocytic cells contained was considered as an internal positive control of functional signaling.

Mononuclear cells were identified on the basis of physical parameters (a) and then selected according to their CD45 reactivity (b). Within the CD45+ cells, monocytes (in green), non-myeloid and myeloid cells could be distinguished by means of their CD33 and CD14 reactivity (c). Phosphoprotein expression was then measured on monocytic gated cells (d–f).
Spontaneous colony growth assay
Freshly isolated PB mononuclear cells were cultured at a density of 5 × 105/ml in methylcellulose medium (Methocult H4230, Stem Cell, Vancouver, BC, Canada) in the presence or absence of 10% conditioned medium from the 5637 tumor cell line and 2 U/ml erythropoietin. Spontaneous and stimulated growth of CFU-GM and BFU-E was determined after 14–21 days of incubation as previously described.24
Statistics
The unpaired t-test (P-value) was used to compare the values of both CFU-GM and BFU-E growth between RAS-mutated patients (n 27) and control subjects (n 35), as well as to compare p-STAT3 and p-ERK values between the same two groups. All statistical tests were performed at a significance level of P<0.05 (two-tailed).
Results
Clonogenic assay
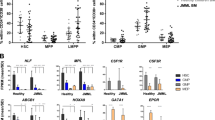
Clonogenic assays from PB precursor cells showed a significantly higher spontaneous growth rate of both BFU-E and CFU-GM in RAS-mutated patients (n=22) than control subjects (n=22). We observed an increased number of BFU-E colonies (mean±s.e.m.): 84.45±7.37% vs 30.50±4.32%, P<0.0001, as well as enhanced growth of CFU-GM, (21.14±3.15% vs 7.23±1.38%, P=0.0002; Figures 2a and b, respectively). Augmented colony formation activity of RAS-mutated cells was also observed upon incubation in growth factor-supplemented medium: BFU-E (78.41±6.60% vs 31.14±4.80%, P<0.0001) and CFU-GM (21.59±2.93% vs 8.95±1.77%, P=0.0006).

Representative quantitative BFU-E (a) and CFU-GM (b) colony numbers as assessed by clonogenic assays. t test, ***P<0.001.
Phosphoflow assay
A significantly enhanced amount of GM-CSF-stimulated p-ERK-positive cells was observed in monocytes isolated from RAS-mutated patients (n=16) as compared with control subjects (n=19): 28.84±5.192% vs 12.74±3.52% (mean±s.e.m.) P=0.0128, respectively (Figure 3a). This enhanced p-ERK response was also observed at lower dose of GM-CSF (0.1 ng/ml) (4.13±2.34% vs 0.28±0.15%) or under basal conditions (3.39±0.94% vs 1.42±0.54%) in RAS-mutated patients (n=9) as compared with control subject (n=12), although these differences were not statistically significant. When we tested the GM-CSF-induced p-STAT5 response in monocytic cells, we failed to find significant differences between patients (n=13) and control subjects (n=17): 91.15±2.5% vs 84.39±3.63% (mean±s.e.m.) at 0.1 ng/ml GM-CSF and 96.65±0.62% vs 96.91±0.49% at 10 ng/ml GM-CSF (Figure 3b). Finally, IL-6-stimulated p-STAT3 levels were also measured in monocytes from patients (n=15) and controls (n=20). Both subgroups responded to such stimuli in a similar fashion (mean±s.e.m. at 50 ng/ml IL-6: 50.49±7.38% vs 50.30±6.55%, P=NS; Figure 3c).

Scatter diagram of p-ERK+ cells treated with 10 ng/ml GM-CSF (a), p-STAT5+ cells treated with 0.1 ng/ml GM-CSF (b) and p-STAT3+ cells treated with 50 ng/ml IL-6 (c). Data are shown as percentages with mean (central bars) and ranges (whiskers). t test, *P<0.05.
Discussion
It has been shown that individuals with NS are at increased risk of developing a myeloproliferative disorder (NS/MPD) that may resolve spontaneously or progress to JMML.25, 26, 27 neuro cardio facial cutaneous syndromes, in particular NS, are related to JMML in terms of both germline and somatic mutations of genes involved in the RAS/MAPK signaling pathway.
Here we demonstrate for the first time that RAS-mutated non-JMML patients are characterized by a significantly higher spontaneous growth rate, in vitro, of both BFU-E- and CFU-GM-circulating progenitors than control subjects. This observation is in good agreement with previous studies using mice carrying germline mutations of PTPN11 (for example, D61Y, E76K and D61G), which showed an increased number of BFU-E and GM-CFU colonies with aberrant differentiation and MAPK-induced GM-CSF hyperactivation.25, 26, 28 Such enhanced growth of circulating hematopoietic progenitors, which we have consistently observed in our cohort of RAS-mutated patients, although not to the same extent of that measured in JMML patients, may reflect a constitutional predisposition to cell hyperproliferation of subjects, such as NS patients, who display a moderately activated RAS/MAPK signaling pathway.
Timeus et al.29 have recently reported a significant decrease in the apoptotic rate of circulating CD34+ hematopoietic progenitors in NS patients suggesting an increased CD34+ cell survival, despite the lack of evidence of an abnormal pattern in both CD34+ cells and CFU-GMs. Thus, this evidence supports a model whereby NS patients displaying aberrant behavior of hematopoietic progenitors are predisposed to developing NS/MPD or JMML.
Mutations of PTPN11 have been shown to promote RAS activation, which in turn induces activation of the Raf/MEK/MAPK cascade.30, 31 Here we demonstrate that such activation can be consistently evaluated in RAS-mutated patients by simply measuring p-ERK levels through accurate single-cell phosphoflow analysis. Thus, our findings seem to indicate that the extent of ERK phosphorylation in hematopoietic progenitors might be a potential new marker for diagnosis and monitoring of NS patients at risk of developing MPDs.
Some authors have hypothesized that Shp-2-mediated STAT3 hypophosphorylation might also have a role in the phenotypic abnormalities observed in NS and/or during the pathogenesis of JMML.22 Contrary to the results by Zhang et al., we did not observed any altered pattern of STAT3 phosphorylation in our series of PB cells from RAS-mutated patients. However, we could not measure p-STAT3 on circulating CD34+/CD33+ myeloid progenitors due to the low number of such cells suitable for phosphoflow analysis. Thus, further studies are clearly needed to shed light on the role played by JAK/STAT signaling in NS/MPDs.
Finally, in good agreement with previous data in JMML patients,23 we could not detect any significant difference between the constitutive STAT5 phosphorylation in RAS-mutated patients vs control subjects.
These data together with the findings by Kotecha et al.,32 who failed to detect an increase in p-STAT5 in NS patients, strongly suggest that lack of p-STAT5 hyperactivation may be related to the hematological benign course of most syndromic cases with Ras mutations.
Recently, Strullu et al. have reported a large prospective cohort of NS patients where ~3% of them met the consensus diagnostic criteria for JMML, and nearly half of them showed severe neonatal manifestations and died within the first month of life.33 As the incidence of childhood RASopathies and MPDs may be underestimated due to such early lethality, the authors recommended that all cases of childhood NS should be systematically evaluated for clinical signs of MPD and basic hematological parameters during the first year of life.33 In this regard, our work provides new insights into the molecular characterization of children with NS that could potentially be exploited for more advanced screening and monitoring of children with RASopathies and MPDs.
Taken together, our results allow us to hypothesize the existence of a ‘hematological phenotype’—in non-JMML NS or RAS-mutated syndromic patients—that is secondary to germline RAS mutations and able to induce a mild activation of the RAS/MAPK cascade. Further studies are needed to fully understand the role of such aberrant patterns during the pathogenesis of hematological diseases in these patients.
References
Gelb BD, Tartaglia M . Noonan syndrome and related disorders: dysregulated RAS-mitogen activated protein kinase signal transduction. Hum Mol Genet 2006; 15: R220–R226.
Kratz CP, Niemeyer CM, Zenker M . An unexpected new role of mutant Ras: perturbation of human embryonic development. J Mol Med 2007; 85: 227–235.
Schubbert S, Shannon K, Bollag G . Hyperactive Ras in developmental disorders and cancer. Nat Rev Cancer 2007; 7: 295–308.
Rauen KA . The RASopathies. Annu Rev Genomics Hum Genet 2013; 14: 355–369.
Roberts AE, Allanson JE, Tartaglia M, Gelb BD . Noonan syndrome. Lancet 2013; 381: 333–342.
Tartaglia M, Zampino G, Gelb BD . Noonan syndrome: clinical aspects and molecular pathogenesis. Mol Syndromol 2010; 1: 2–26.
Tartaglia M, Mehler EL, Goldberg R, Zampino G, Brunner HG, Kremer H et al. Mutations in PTPN11, encoding the protein tyrosine phosphatase SHP-2, cause Noonan syndrome. Nat Genet 2001; 29: 465–468.
Liu X, Qu CK . Protein tyrosine phosphatase SHP-2 (PTPN11) in hematopoiesis and leukemogenesis. J Signal Transduct 2011; 2011: 195239.
Bader-Meunier B, Tchernia G, Mielot F, Fontaine JL, Thomas C, Lyonnet S et al. Occurrence of myeloproliferative disorder in patients with Noonan syndrome. J Pediatr 1997; 130: 885–889.
Choong K, Freedman MH, Chitayat D, Kelly EN, Taylor G, Zipursky A . Juvenile myelomonocytic leukemia and Noonan syndrome. J Pediatr Hematol Oncol 1999; 21: 523–527.
Hasle H . Malignant diseases in Noonan syndrome and related disorders. Horm Res 2009; 72: 8–14.
Bastida P, Garcia-Minaur S, Ezquieta B, Dapena JL, Sanchez de Toledo J . Myeloproliferative disorder in Noonan syndrome. J Pediatr Hematol Oncol 2011; 33: e43–e45.
Chan RJ, Leedy MB, Munugalavadla V, Voorhorst CS, Li Y, Yu M et al. Human somatic PTPN11 mutations induce hematopoietic-cell hypersensitivity to granulocyte-macrophage colony-stimulating factor. Blood 2005; 105: 3737–3742.
Emanuel PD, Bates LJ, Castleberry RP, Gualtieri RJ, Zuckerman KS . Selective hypersensitivity to granulocyte-macrophage colony-stimulating factor by juvenile chronic myeloid leukemia hematopoietic progenitors. Blood 1991; 77: 925–929.
Kratz CP, Niemeyer CM, Castleberry RP, Cetin M, Bergstrasser E, Emanuel PD et al. The mutational spectrum of PTPN11 in juvenile myelomonocytic leukemia and Noonan syndrome/myeloproliferative disease. Blood 2005; 106: 2183–2185.
Mohi MG, Williams IR, Dearolf CR, Chan G, Kutok JL, Cohen S et al. Prognostic, therapeutic, and mechanistic implications of a mouse model of leukemia evoked by Shp2 (PTPN11) mutations. Cancer Cell 2005; 7: 179–191.
Schubbert S, Lieuw K, Rowe SL, Lee CM, Li X, Loh ML et al. Functional analysis of leukemia-associated PTPN11 mutations in primary hematopoietic cells. Blood 2005; 106: 311–317.
Tartaglia M, Martinelli S, Stella L, Bocchinfuso G, Flex E, Cordeddu V et al. Diversity and functional consequences of germline and somatic PTPN11 mutations in human disease. Am J Hum Genet 2006; 78: 279–290.
Tartaglia M, Niemeyer CM, Fragale A, Song X, Buechner J, Jung A et al. Somatic mutations in PTPN11 in juvenile myelomonocytic leukemia, myelodysplastic syndromes and acute myeloid leukemia. Nat Genet 2003; 34: 148–150.
Loh ML, Vattikuti S, Schubbert S, Reynolds MG, Carlson E, Lieuw KH et al. Mutations in PTPN11 implicate the SHP-2 phosphatase in leukemogenesis. Blood 2004; 103: 2325–2331.
Matozaki T, Murata Y, Saito Y, Okazawa H, Ohnishi H . Protein tyrosine phosphatase SHP-2: a proto-oncogene product that promotes Ras activation. Cancer Sci 2009; 100: 1786–1793.
Zhang W, Chan RJ, Chen H, Yang Z, He Y, Zhang X et al. Negative regulation of Stat3 by activating PTPN11 mutants contributes to the pathogenesis of Noonan syndrome and juvenile myelomonocytic leukemia. J Biol Chem 2009; 284: 22353–22363.
Hasegawa D, Bugarin C, Giordan M, Bresolin S, Longoni D, Micalizzi C et al. Validation of flow cytometric phospho-STAT5 as a diagnostic tool for juvenile myelomonocytic leukemia. Blood Cancer J 2013; 3: e160.
Schiro R, Longoni D, Rossi V, Maglia O, Doni A, Arsura M et al. Suppression of juvenile chronic myelogenous leukemia colony growth by interleukin-1 receptor antagonist. Blood 1994; 83: 460–465.
Araki T, Mohi MG, Ismat FA, Bronson RT, Williams IR, Kutok JL et al. Mouse model of Noonan syndrome reveals cell type- and gene dosage-dependent effects of Ptpn11 mutation. Nat Med 2004; 10: 849–857.
Niemeyer CM . RAS diseases in children. Haematologica 2014; 99: 1653–1662.
Tartaglia M, Gelb BD . Germ-line and somatic PTPN11 mutations in human disease. Eur J Med Genet 2005; 48: 81–96.
Chan G, Kalaitzidis D, Usenko T, Kutok JL, Yang W, Mohi MG et al. Leukemogenic Ptpn11 causes fatal myeloproliferative disorder via cell-autonomous effects on multiple stages of hematopoiesis. Blood 2009; 113: 4414–4424.
Timeus F, Crescenzio N, Baldassarre G, Doria A, Vallero S, Foglia L et al. Functional evaluation of circulating hematopoietic progenitors in Noonan syndrome. Oncol Rep 2013; 30: 553–559.
De Rocca Serra-Nedelec A, Edouard T, Treguer K, Tajan M, Araki T, Dance M et al. Noonan syndrome-causing SHP2 mutants inhibit insulin-like growth factor 1 release via growth hormone-induced ERK hyperactivation, which contributes to short stature. Proc Natl Acad Sci USA 2012; 109: 4257–4262.
Xu D, Wang S, Yu WM, Chan G, Araki T, Bunting KD et al. A germline gain-of-function mutation in Ptpn11 (Shp-2) phosphatase induces myeloproliferative disease by aberrant activation of hematopoietic stem cells. Blood 2010; 116: 3611–3621.
Kotecha N, Flores NJ, Irish JM, Simonds EF, Sakai DS, Archambeault S et al. Single-cell profiling identifies aberrant STAT5 activation in myeloid malignancies with specific clinical and biologic correlates. Cancer Cell 2008; 14: 335–343.
Strullu M, Caye A, Lachenaud J, Cassinat B, Gazal S, Fenneteau O et al. Juvenile myelomonocytic leukaemia and Noonan syndrome. J Med Genet 2014; 51: 689–697.
Acknowledgements
This work was supported by Comitato M.L. Verga and Fondazione Tettamanti; we also thank Mariani Foundation (Milan) for supporting the clinical activity of U.O.S Genetica Clinica Pediatrica Fondazione MBBM. CB, JS, PB and PC performed the research; GG, AS and AB designed the research study; CB and JS analyzed the data; GG wrote the paper.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Gaipa, G., Bugarin, C., Cianci, P. et al. Peripheral blood cells from children with RASopathies show enhanced spontaneous colonies growth in vitro and hyperactive RAS signaling. Blood Cancer Journal 5, e324 (2015). https://doi.org/10.1038/bcj.2015.52
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/bcj.2015.52
