
Abstract
When a ribosome translates mRNA sequences, the ribosome often stalls at certain codons because it is hard to translate. Consecutive proline codons are such examples that induce ribosome stalling and elongation factor P (EF-P) is required for the stalled ribosome to continue translation at those consecutive proline codons. We found that EF-P is required for translation of the mgtB gene encoding a Mg2+ transporter in the mgtCBR virulence operon from the intracellular pathogen Salmonella enterica serovar Typhimurium. Salmonella lacking EF-P decreases MgtB protein levels in a manner dependent on consecutive proline codons located in the mgtB coding region despite increasing transcription of the mgtCBR operon via the mgtP open reading frame in the leader RNA, resulting in an altered ratio between MgtC and MgtB proteins within the operon. Substitution of the consecutive proline codons to alanine codons eliminates EF-P-mediated control of the mgtB gene during infection and thus contributes to Salmonella’s survival inside macrophages where Salmonella experiences low levels of EF-P. This suggests that this pathogen utilizes a strategy to coordinate expression of virulence genes by an evolutionarily conserved translation factor.
Similar content being viewed by others

Introduction
Mg2+ is involved in many important biological processes including coordinating nucleotides, stabilizing ribosome or membrane, and participating in many enzymatic reactions1. For pathogens, the ability to acquire Mg2+ or maintain Mg2+ homeostasis is critical to survive within a host and cause diseases. In the intracellular pathogen Salmonella enterica serovar Typhimurium, Mg2+ transport is tightly regulated by three distinct loci-mgtA, mgtCBR, and corA encoding MgtA, MgtB, and CorA Mg2+ transporters respectively2. A strain lacking all three Mg2+ transporters could not support growth unless medium was supplemented with at least 10 mM Mg2+ 3. Among them, MgtB orthologs occur in most limited phylogenetic distribution and appear to be associated with enteric bacteria replicating within warm-blooded host during infection cycle2,4,5. Moreover, the fact that the mgtB gene is located in the Salmonella pathogenicity islands 3 (SPI-3)6 supports the notion that MgtB might contribute to Salmonella’s pathogenicity. More recently, it was reported that the mgtB gene is required for virulence in Yersinia pestis7.
Salmonella MgtB is a Mg2+ -transporting P-type ATPase8 that typically induces a conformation change of the protein itself mediated by phosphorylation at a conserved aspartic acid residue during transport cycle1,2. MgtB is an inner membrane protein with 10 transmembrane domains and the conserved aspartic acid critical for the P-type ATPase protein family is located at the cytosolic face between 4th and 5th transmembrane segments9. In agreement with the notion that MgtB mainly mediates Mg2+ influx, transcription of the mgtB gene is induced in a low Mg2+ environment by the PhoP/PhoQ two-component system10, which is a major transcriptional regulator for Salmonella virulence11. In fact, the mgtB gene is a part of the mgtCBR operon and cotranscribed with two other genes, mgtC and mgtR, encoding the MgtC protein required for macrophage survival and mouse virulence and the MgtR short-peptide regulator for MgtC proteolysis respectively12,13,14. Because the mgtB gene is transcribed from a single promoter located upstream of the mgtC gene as a part of the mgtCBR polycistronic messages15, one might expect that Salmonella produces MgtC and MgtB proteins at similar levels. However, this is clearly not the case for the mgtCBR operon because the AmgR RNA is transcribed from the mgtC-mgtB intergenic region and alters MgtC and MgtB protein levels by preferentially degrading the mgtC part of the polycistronic messages15. Here we report that Salmonella achieves altered MgtC and MgtB protein levels in the mgtCBR operon by another strategy and the altered MgtC and MgtB protein levels contribute to Salmonella virulence.
During protein translation, a ribosome often slows down or stalls on transcripts depending on mRNA sequences. Consecutive proline codons are such sequences because the α-imino group of the proline codons constrains peptide bond formation between the two proline codons16 and thus induces ribosome stalling when it appears in a consecutive fashion17,18. Elongation factor P is specifically required for the stalled ribosome to resume translation17,18 by promoting peptide bond formation between peptidyl-prolyl-tRNA at the ribosomal P site and incoming prolyl-tRNA at the A site19. Therefore, a presence or absence of EF-P controls expression of genes harboring consecutive proline codons by limiting translation and the proteins with the consecutive proline codons are expected to be less abundant in a strain lacking EF-P. Alternatively or in addition, EF-P controls transcription of genes preceded by a leader RNA if the leader RNA harbors a short open reading frame with consecutive proline codons and the sequences associated with the short ORF have a potential to form two sets of stem-loop structures, one of which functions as a transcription attenuator. Although bacterial transcription and translation are usually coupled, the presence of the consecutive proline codons that induce ribosome stalling could uncouple transcription and translation. In this particular case, the presence or absence of EF-P determines whether or not the ribosome stalls at the consecutive proline codons of the ORF in the leader RNA, thereby coupling/uncoupling between transcription of the leader RNA and translation of the short ORF within the leader. By doing so, EF-P controls the formation of the transcription attenuator that prevents transcription elongation into the downstream genes. EF-P controls transcription elongation of the mgtCBR operon by such a transcription attenuation-like mechanism via the 296 nt-long leader RNA harboring mgtP with three consecutive proline codons (Fig. 1)20,21. Salmonella lacking EF-P induces ribosome stalling at the consecutive proline codons in mgtP and allows the formation of stem-loop E structure, which enhances mgtCBR transcription (Fig. 1)21. This makes physiological sense because Salmonella indeed decreases EF-P mRNA levels during infection21 and it explains why the mgtC gene is highly expressed among other PhoP-regulated genes inside macrophages22,23.

EF-P controls expression of the mgtCBR operon in two different ways. On the one hand, EF-P controls transcription elongation of the mgtCBR operon by the leader RNA harboring the proline-rich short open reading frame mgtP. Three consecutive proline codons in mgtP induce ribosome stalling and require EF-P to continue translation. EF-P-mediated control in translating mgtP allows to from one of two alternative stem-loop structures (stem-loops D versus E) that control transcription elongation into the downstream region. On the other hand, EF-P controls translation of the mgtB part of the mgtCBR polycistronic messages by two consecutive proline codons located in the mgtB coding region. Therefore, when EF-P is absent or present in low levels (B), Salmonella promotes mgtCBR transcript levels by inducing ribosome stalling at the proline codons in mgtP and thus allowing the formation of stem-loop E structure. However, at the same time, Salmonella lacking EF-P decreases MgtB production by inducing ribosome stalling at the proline codons located in the mgtB coding region, thereby resulting in high MgtC: MgtB ratios compared to Salmonella producing EF-P (A). The sequences of the mgtP and mgtB variants used in this work are indicated above the Pro codons.
In this paper, we described another layer of regulation in the mgtCBR operon mediated by EF-P. Salmonella lacking EF-P decreases MgtB protein levels via two consecutive proline codons located in the mgtB coding region despite increasing transcription of the entire mgtCBR operon by the leader RNA. Removal of EF-P-mediated control in the MgtB protein levels promotes Salmonella’s pathogenicity, implying that the ability to transport Mg2+ must be compromised during the course of infection.
Results
Inactivation of efp derepresses transcription of the mgtCBR operon by the leader mRNA harboring mgtP
The mgtCBR leader RNA harbors mgtP, an 18 amino-acid long ORF with three consecutive proline codons (Fig. 1). Because mgtP is located adjacent to a sequence adopting stem-loop E structure (Fig. 1), the presence of the consecutive proline codons that induce ribosome stalling allows to form stem-loop E structure and promotes transcription into the downstream coding region. Therefore, efp inactivation was expected to induce transcription of the mgtC and mgtB genes in a manner dependent on consecutive proline codon at mgtP. Indeed, when grown in media containing 500 μM Mg2+ for 1 h to initiate transcription from the PhoP-dependent promoter, the efp mutant increased mRNA levels of both the mgtC and mgtB genes by ~100 fold (Fig. S1) and derepression in mRNA levels were not detected in a derivative strain with the mgtP proline codons substituted by threonine codons (Fig. S1).
Loss of efp decreases MgtB protein levels in a PhoP/PhoQ-inducing condition
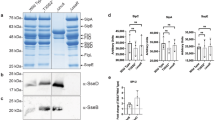
We assumed that Salmonella lacking EF-P would produce higher amounts of MgtB proteins based on the findings that efp inactivation increased transcription of the mgtC and mgtB genes (Fig. S1) and produced higher amounts of the MgtC proteins21 and that the mgtC and mgtB genes are transcribed from a single promoter located upstream of the mgtC gene15. To test this assumption, we measured the amounts of the MgtB proteins in low Mg2+ media to activate transcription of the mgtCBR operon by the PhoP/PhoQ two-component system10. Strikingly, MgtB proteins were barely detected in the efp mutant compared to those in the wild-type Salmonella (Fig. 2A). This is in contrast with the MgtC proteins that were produced in a higher amount in the efp mutant (Fig. 2B)21. Control experiments were carried out as follows: Both MgtB and MgtC proteins were not detected in a non-inducing media containing 10 mM Mg2+ (Fig. 2A and B), efp inactivation eliminates EF-P production (Fig. 2C), and Fur protein levels, which were used as loading control, were unaffected in all growth conditions (Fig. 2D).

Western blot analysis of crude extracts prepared from either wild-type (14028s) or the efp mutant Salmonella (DN337). Blots were probed with anti-MgtB (A), anti-MgtC (B), anti-EF-P (C), or anti-Fur (D) antibodies to detect MgtB, MgtC, EF-P, and Fur proteins respectively. Bacteria were grown for 5 h in N-minimal media containing 10 mM or 0.01 mM Mg2+ as described in Methods.
The consecutive proline codons at positions 555 and 556 are required for EF-P-mediated MgtB production
Given that EF-P’s role is limited to rescue stalled ribosome at consecutive proline codons17,18, low levels of MgtB in the efp mutant indicate that the mgtB gene might have consecutive proline codons responsible for EF-P-mediated control. We searched for the presence of consecutive proline codons in the mgtB coding region. Amino acid sequence analysis revealed that the mgtB gene has two pairs of consecutive proline codons in the coding region. One is located at positions 73 and 74 in the N-terminal cytoplasmic region of the MgtB protein and the other is located at positions 555 and 556 in the cytoplasmic loop region between 4th and 5th transmembrane domains (Fig. 3A). To determine which consecutive proline codons are required for EF-P-mediated MgtB expression, we created chromosomal mutant strains where respective two consecutive proline codons were replaced with alanine codons. Then, we compared MgtB protein levels of each mutant in either the wild-type and efp mutant background. When grown in low Mg2+ media, the Pro 555, 556 Ala substitution resisted to decrease MgtB levels despite efp inactivation, indicating that proline codons at positions 555 and 556 are critical for EF-P-mediated control of the mgtB gene (Fig. 3B). By contrast, a Salmonella mutant strain with the Pro 73, 74 Ala substitution behaved just like the isogenic strain with the wild-type mgtB gene (Fig. 3B). It is consistent with the previous notion that adjacent or neighboring sequences affect how strongly the ribosome stalls at those consecutive proline codons because Pro 555, 556 codons preceded by Asp 554 were strongly affected by EF-P while Pro 73, 74 codons preceded by Val 72 were not24,25,26. Control experiments proved that substitutions at consecutive proline codons in the mgtB gene did not affect the expression behavior of MgtC (Fig. 3C, EF-P (Fig. 3D, and Fur (Fig. 3E) protein levels.

(A) Schematic diagram of the MgtB protein and location of proline codons used in this work. (B–D) Western blot analysis of crude extracts prepared from a strain with the wild-type mgtB gene (14028s), the efp mutant (DN337), an mgtB derivative with the Pro 73, 74 codons substituted by Ala codons (EN793), a mutant with both the Pro 73, 74 substitution and the efp insertion (EN794), an mgtB derivative with the Pro 555, 556 codons substituted by Ala codons (EN873), or a mutant with both the Pro 555, 556 substitution and the efp insertion (EN877). Blots were probed with anti-MgtB (B), anti-MgtC (C), anti-EF-P (D) or anti-Fur (E) antibodies to detect MgtB, MgtC, EF-P and Fur proteins respectively. Bacteria were grown for 5 h in N-minimal media containing 0.01 mM Mg2+ as described in Methods.
Salmonella with the mgtB Pro 555, 556 substitution carries a Mg2+ -transporting activity in a strain lacking other Mg2+ transporters
The mgtB gene encodes a Mg2+ transporting P-type ATPase with 10-transmenbrane segments and the Pro 555 and 556 residues are located in the large cytoplasmic loop that lies between transmembrane 4 and 59. Because previous amino acid sequence analysis implicated that Asp 554 seems to be involved in ATP binding necessary for Mg2+ transport8, we wondered whether the Pro 555, 556 Ala substitution affects MgtB’s ability to transport Mg2+. To explore this, we created isogenic strains where two other Mg2+ transporter genes, corA and mgtA, were inactivated in either the wild-type mgtB or the mgtB derivative Salmonella with the Pro 555, 556 Ala substitution in its original chromosomal location. Given that growth of the corA and mgtA double knockout strain would be dependent on only functional MgtB, we tested MgtB’s activity to transport Mg2+ by determining ability to grow in low Mg2+ media. The mgtB derivative with the Pro 555, 556 Ala substitution grew well just like the isogenic strain with the wild-type mgtB gene in low Mg2+ media (Fig. 4A), demonstrating that MgtB with the Pro 555, 556 Ala substitution is functional for transporting Mg2+ to support growth in low Mg2+ media. By contrast, an introduction of mgtB deletion to the isogenic strain generated a triple knockout strain lacking all three Mg2+ transporters in Salmonella and Salmonella lacking all three Mg2+ transporters could not support growth in low Mg2+ media (Fig. 4A). As a control, 100 mM Mg2+ supplement in the liquid media restored growth of all strains (Fig. 4B).

(A,B) Growth curves of strains with the wild-type mgtB gene (EL496), the mgtB derivative with the Pro 555, 556 codons substituted by Ala codons (EN943), or the mgtB insertion (EL498) in a genetic background where both the corA and mgtA genes are deleted. Bacteria were grown in N-minimal medium containing 0.01 mM (A) or 100 mM Mg2+ (B) at 37 °C for 6 h in a 96-well plate with orbital shaking and measured absorbance at OD600 every 2.5 min.
EF-P affects normal MgtB expression when Salmonella is inside macrophages
The results described above indicate that EF-P controls translation of the mgtB gene via consecutive proline codons at positions 555 and 556. We wondered what the physiological relevance of EF-P-mediated control in the mgtB gene would be. Because the mRNA levels of EF-P decrease during the course of infection21, one might expect that Salmonella would decrease MgtB protein levels inside macrophages. And, if this is the case, the mgtB derivative with the Pro 555, 556 Ala substitution would not be subject to EF-P-mediated control and the levels of MgtB will remain high even inside host cells. To explore this possibility, we measured MgtB protein amounts in the wild-type or the mgtB derivative Salmonella inside J774 A.1 macrophages. As expected, the mgtB derivative strain with the Pro 555, 556 Ala substitution produced higher levels of MgtB proteins compared to those of the wild-type at 21 h after infection (Fig. 5A). Consistent with previous observations, EF-P appeared to control MgtB expression at the level of translation because the mRNA levels of the wild-type and mgtB derivative with the Pro 555, 556 substitution were similar inside macrophages (Fig. S2). Control experiments proved that: first, mgtB inactivation abolished MgtB production (Fig. 5A). Second, the levels of MgtC and CorA proteins were similar in the wild-type and mgtB derivative strains (Fig. 5B and C). Therefore, this suggests that, under normal circumstances, Salmonella decreases MgtB expression inside macrophages by limiting translation of mgtB mRNAs via low levels of EF-P.

Western blot analysis of crude extracts prepared from wild-type (14028s), the mgtB derivative with the Pro 555, 556 codons substituted by Ala codons (EN873) or an mgtB deletion mutant (EN481) Salmonella inside J774 A.1 macrophage-like cells 21 h after infection. The amounts of MgtB (A) and MgtC (B) proteins were determined by anti-MgtB or anti-MgtC antibodies. Anti-CorA antibodies were used to detect CorA proteins (C) as loading controls.
The mgtB Pro 555, 556 Ala mutant Salmonella displays a hypervirulent phenotype in intramacrophage survival and mouse virulence
If Salmonella decreases EF-P mRNA levels during infection, low levels of EF-P would induce ribosome stalling at three consecutive proline codons of mgtP located in the leader RNA and thus increase transcription of the mgtCBR operon. This explains in part why transcription of the mgtC gene is highly induced inside macrophages20,22. It also makes sense because the mgtC gene is required for survival inside macrophages and virulence in mice6,14. At the same time, however, low levels of EF-P would decrease translation of the mgtB part of the mgtCBR polycistronic messages via two consecutive proline codons located in the mgtB coding region (Fig. 3). If this is the case, it will create a situation where Salmonella decreases MgtB production while still increases MgtC production inside host cells. We wondered whether the decrease in MgtB levels via EF-P has a physiological impact on Salmonella’s ability to replicate within macrophages. To address this question, we measured survival inside macrophage-like J774A. 1 cell line using the Salmonella mgtB derivative with the Pro 555, 556 Ala substitution because it is not subjected to EF-P-mediated control and it produced higher amounts of MgtB protein inside macrophages (Fig. 5). The Pro 555, 556 Ala substitution increased Salmonella’s replication inside macrophages by nearly 200% relative to that of wild-type Salmonella (Fig. 6A), suggesting that enhanced production of MgtB proteins promotes Salmonella’s survival inside a host, presumably by increasing the ability to uptake Mg2+ from the host. By contrast, the mgtB deletion mutant did not have a significant defect in intramacrophage survival (Fig. 6A). As a control, the mgtC mutant showed a remarkable defect in intramacrophage survival (Fig. 6A) as previously described6,27. Similar to those from intramacrophage survival, when we injected Salmonella strains listed above into mice intraperitoneally, the mgtB Pro 555, 556 Ala substitution renders Salmonella hypervirulent (Fig. 6B and C). These results showed that EF-P-mediated control of MgtB protein levels is critical for Salmonella virulence.

(A) Replication inside J774 A.1 macrophages of wild-type (14028s), the mgtB derivative with Pro codons replaced by Ala codons (EN873), the mgtB deletion mutant (EN481), or an mgtC deletion mutant (EL4) Salmonella at 21 h after infection. (B,C) Survival of C3H/HeN mice inoculated intraperitoneally with ~200 (B) or ~3000 (C) colony forming units of the Salmonella strains listed above.
Discussion
Here we established that Salmonella decreases MgtB protein levels by lowering EF-P levels during infection. Such a decrease in MgtB levels mediated by EF-P is dependent on the consecutive proline codons located in the mgtB coding region (Fig. 3) and the decrease in MgtB protein levels is required for Salmonella’s ability to survive within macrophages and mouse virulence (Fig. 6). It is interesting to discover the decrease in MgtB levels inside host cells because low levels of EF-P during Salmonella infection actually induce transcription of the entire mgtCBR operon via consecutive proline codons at mgtP located in the mgtCBR leader RNA (Fig. 1)21. Then, one can easily imagine that Salmonella increases MgtC protein production but decreases MgtB’s during the course of infection, establishing high MgtC: MgtB protein ratios inside host cells. Salmonella seems to limit proliferation inside macrophages by decreasing the production of the Mg2+-importing MgtB transporter because substitution of the consecutive proline codons in the mgtB coding region that prevents downregulation of MgtB protein levels promotes Salmonella’s pathogenicity both in terms of macrophage survival and mouse virulence (Fig. 6). We wondered why Salmonella achieves differential protein levels between MgtC and MgtB within the same operon. We speculated that it might be due to homeostasis between ATP and Mg2+ levels during infection because the MgtC protein inhibits proton translocation of F1Fo ATP synthase within an acidified phagosome to promote Salmonella’s pathogenicity, resulting in a decrease in ATP production27. Given that Mg2+ is required to coordinate ATP for neutralizing its negative charges, the decrease in ATP production via enhanced MgtC protein levels could be compromised by the decrease in the ability to transport Mg2+ ion via the decrease in MgtB protein levels in the host environment.
EF-P-mediated differential regulation that takes place in the mgtCBR operon has several properties in common with those achieved by the AmgR RNA in the same operon. First, differential regulation within the operon is mediated by the same proteins. We described here that EF-P controls transcription elongation of the mgtCBR operon and, at the same time, controls translation of the mgtB part of the mRNA message. Similarly, PhoP controls transcription of the mgtCBR full-length messages as well as the AmgR antisense transcript initiated from mgtC-mgtB intergenic region toward the mgtC gene15. Second, mutations that prevent differential regulation promote Salmonella’s pathogenicity. The substitution that eliminates EF-P-mediated control in the mgtB gene increases MgtB protein levels, resulting in elevated proliferation inside macrophages and hypervirulence in mice. Likewise, the substitution that removes the promoter region of the amgR accumulates preferentially MgtC proteins and, to a lesser extent, MgtB proteins, leading to a hypervirulence phenotype in mice15. However, they clearly differ from each other in terms of how they achieve differential regulation within the same operon because EF-P does its task by controlling ribosome stalling on the mRNA messages whereas AmgR by degrading mRNA in an RNase E-mediated antisense mechanism. Moreover, PhoP binds the promoter of the mgtCBR operon with a higher affinity than that of the amgR15, gradually establishing different levels between MgtC and MgtB proteins inside host cells. This is in contrast with the fact that low levels of EF-P enable to maintain a steady-state MgtC: MgtB ratio during infection.
For Salmonella Typhimurium, ability to survive and replicate within a phagosome inside host macrophages is critical to cause diseases28. EF-P seems to be required for Salmonella virulence based on following: Inactivation of two genes specifying proteins required for EF-P’s activity17,18,29 is highly attenuated for mouse virulence29. And also a previous proteomic approach identified that many virulence genes were upregulated in a strain lacking functional EF-P29. Because EF-P is involved in translation of many genes with a variety of physiological functions17,18,30, it has been an intriguing question to figure out which genes are responsible for the virulence phenotype of strains lacking functional EF-P. Given that EF-P’s role in virulence could be limited to genes with consecutive proline codons in the coding region or in the short ORF located in the preceding leader RNA, the simplest scenario is that a particular virulence gene(s) controlled by EF-P represents EF-P’s virulence phenotype. In other words, EF-P controls translation of the particular virulence gene with consecutive proline codons and thus the strain lacking a functional EF-P would decrease the abundance of the corresponding proteins, in turn, resulting in an avirulent phenotype. In this case, one can expect that a deletion mutant of the particular gene would behave similarly to the mutants of genes encoding EF-P itself or EF-P modifying enzymes. Moreover, substitution of consecutive proline codons in the particular gene that eliminates EF-P-mediated control is expected to suppress the defect in the virulence phenotype of the strain lacking functional EF-P. However, the findings how EF-P controls expression of the mgtCBR operon and contributes to Salmonella’s pathogenicity imply that the virulence phenotype of the strain lacking functional EF-P is not so simple as we hypothesized. Low levels of EF-P during infection promote Salmonella pathogenicity by inducing production of the MgtC virulence protein via consecutive proline codons at mgtP in the leader RNA21 but, at the same time, limit the pathogen’s proliferation inside host by decreasing production of the MgtB Mg2+ transporter via another consecutive proline codons in the mgtB coding region. Substitution of proline codons in mgtP attenuates Salmonella virulence31 whereas substitution of proline codons in the mgtB coding region renders Salmonella hypervirulent (Fig. 6). Therefore, the virulence phenotype displayed by the strain lacking functional EF-P must be a reflection of the summation between two opposing effects caused by MgtC and MgtB proteins. And there are possibly more responsible genes out there.
Methods
Bacterial strains, plasmids, oligodeoxynucleotides and growth conditions
Bacterial strains and plasmids used in this study are listed in Table S1. All Salmonella enterica serovar Typhimurium strains are derived from the wild-type strain 14028s32 and were constructed by phage P22-mediated transductions as described33. All DNA oligonucleotides are listed in Table S2. Bacteria were grown at 37 °C in Luria-Bertani broth (LB), N-minimal media (pH 7.7)8 supplemented with 0.1% casamino acids, 38 mM glycerol and the indicated concentrations of MgCl2. Escherichia coli DH5α was used as the host for preparing plasmid DNA. Ampicillin was used at 50 μg ml−1, chloramphenicol at 25 μg ml−1, kanamycin at 50 μg ml−1, or tetracycline at 10 μg ml−1.
Effect of efp inactivation on gene expression
Gene expression upon efp inactivation was measured as described previously21. Briefly, bacteria were grown overnight in N-minimal medium containing 10 mM Mg2+. 1/100 dilution of the overnight culture was used to inoculate 20 ml of the same medium and grown for 3 h. Cells were then washed and transferred to 20 ml of N-minimal medium containing 500 μM Mg2+ and grown for 1 h. Bacteria were stabilized using RNAprotect Bacteria Reagent (Qiagen) and RNA was isolated for further analysis.
Quantitative real time-polymerase chain reaction (RT-PCR)
Total RNA was isolated using RNeasy Kit (Qiagen) according to the manufacturer’s instructions. The purified RNA was quantified using a Nanodrop machine (NanoDrop Technologies). cDNA was synthesized using PrimeScriptTM RT reagent Kit (TaKaRa). The mRNA levels of the mgtC, mgtB, efp, and rrsH genes were measured by quantification of cDNA using SYBR Green PCR Master Mix (TOYOBO) and appropriate primers (mgtC: 7530/7531, mgtB:7763/7764 and efp: qrt-efp-F/qrt-efp-R) and monitored using a 7300 Real Time PCR system (Applied Biosystems, Foster City). The mRNA levels of each target genes were calculated using a standard curve of 14028s genomic DNA with known concentration and data were normalized to the levels of 16S ribosomal RNA amplified with primers 6970 and 6971.
Western Blot Analysis
Cells were grown for 5 h in 35 ml of N-minimal medium containing 10 μM Mg2+. Cells were normalized by measuring optical density at 600 nm (OD600). Crude extracts were prepared in PBS (Phosphate-buffered saline) buffer by sonication and analyzed as described15. The data are representative of two independent experiments, which gave similar results.
Measuring growth of strains lacking Mg2+ transporters
Salmonella lacking all three Mg2+ transporters requires 100 mM Mg2+ in the medium to support growth3. To address whether proline to alanine substitution of the mgtB gene could support growth in a Salmonella strain lacking other two Mg2+ transporters, MgtA and CorA34, bacteria were grown in N-minimal medium containing 0.01 mM (Mg2+ limiting) or 100 mM Mg2+ (Mg2+ in excess). Growth was determined at 37 °C for 6 h in a 96-well plate with orbital shaking and absorbance measured at OD600 every 2.5 min.
Examining survival inside macrophages
Intramacrophage survival assays were conducted with the macrophage-like cell line J774 A.1 as described6.
Examining gene expression inside macrophages
Gene expression inside macrophages was measured as described previously23.
Mouse virulence assays
Six- to eight-week-old female C3H/HeN mice were inoculated intraperitoneally with ~102 or ~103 colony-forming units. Mouse survival was followed for 21 days. Virulence assays were conducted twice with similar outcomes, and data correspond to groups of five mice. All procedures were performed according to approved protocols by the Institutional Animal Care and Use Committee from Kangwon National University.
Construction of chromosomal mutant strains with the mgtB proline codons substituted by alanine codons
To generate strains with chromosomal mutations in the mgtB coding region, we used the fusaric acid-based counterselection method as described previously15. First, we introduced TetR cassettes in two different regions of the mgtB gene as follows: we generated PCR products harboring the tetRA genes using as template chromosomal DNA from strain MS7953s and primers KHU336/KHU337 (for 73rd and 74th proline codons) and KH472/KH473 (for 555th and 556th proline codons). The product was purified using a QIAquick PCR purification kit (QIAGEN) and used to electroporate Salmonella 14028s containing plasmid pKD4635. The resulting mgtB NT::tetRA (EN786) and mgtB 900 nt::tetRA (EN821) strains containing plasmid pKD46 were kept at 30 °C. Then, we replaced the tetRA cassettes by preparing DNA fragments carrying proline to alanine codons substitutions in mgtB at positions 73 and 74 or 555 and 556 were prepared by a two-step PCR process. For the first PCR, we used two sets of primer pairs 7554/KH379 and KH378/12605R (for 73rd and 74th proline codons) and KH476/KH475 and KH474/KH477 (for 555th and 556th proline codons), and 14028s genomic DNA as template. For the second PCR, we mixed the two PCR products from the first PCR as templates and amplified DNA fragments using primers 7554/12605R (for 73rd and 74th proline codons) and KH476/KH477 (for 555th and 556th proline codons). The resulting PCR products were purified and integrated into the EN786 and EN821 chromosome and selected against tetracycline resistance with media containing fusaric acid to generate EN793 (mgtBPro 73,74 Ala) and EN873 (mgtBPro 555,556 Ala), tetracycline-sensitive, ampicillin-sensitive (TetS AmpS) chromosomal mutants, respectively. The presence of the expected nucleotide substitutions was verified by DNA sequencing. A P22 phage lysate grown in strain DN337 was used to transduce strains EN793, and EN873 Salmonella selecting for chloramphenicol resistance to generate EN794 (mgtBPro 73,74 Ala, efp::CmR), and EN877 (mgtBPro 555,556 Ala, efp::CmR), respectively.
Construction of strains with chromosomal deletions of the mgtB or corA genes
Salmonella strains deleted for the mgtB or corA genes were generated by the one-step gene inactivation method35. A KmR cassette for the mgtB gene and a CmR cassette for the corA gene were PCR amplified from plasmid pKD4 or pKD3 using primers DE-mgtB-F/DE-mgtB-R (for mgtB), and del-corA-F/del-corA-R (for corA) and the resulting PCR products were integrated into the 14028s chromosome to generate EN480 (mgtB::KmR), and YS166 (corA::CmR), respectively. The mgtB strain (EN481) was generated by removing the KmR cassette from EN480 using plasmid pCP20 as described35. A P22 phage lysate grown in strain YS166 was used to transduce EG9521 Salmonella selecting for chloramphenicol resistance to generate EL496 (corA::CmR, mgtA::MudJ). A P22 phage lysate grown in strain EL496 was used to transduce strains EN481, and EN873 Salmonella selecting for chloramphenicol and kanamycin resistance to generate EL498 (corA::CmR, mgtA::MudJ, ΔmgtB), and EN943 (corA::CmR, mgtA::MudJ, mgtBPro 555,556 Ala), respectively.
Additional Information
How to cite this article: Choi, E. et al. Elongation factor P restricts Salmonella’s growth by controlling translation of a Mg2+ transporter gene during infection. Sci. Rep. 7, 42098; doi: 10.1038/srep42098 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Smith, R. L. & Maguire, M. E. Microbial magnesium transport: unusual transporters searching for identity. Mol Microbiol 28, 217–226 (1998).
Groisman, E. A. et al. Bacterial Mg2+ homeostasis, transport, and virulence. Annu Rev Genet 47, 625–646 (2013).
Hmiel, S. P., Snavely, M. D., Florer, J. B., Maguire, M. E. & Miller, C. G. Magnesium transport in Salmonella typhimurium: genetic characterization and cloning of three magnesium transport loci. J Bacteriol 171, 4742–4751 (1989).
Perez, J. C. et al. Evolution of a bacterial regulon controlling virulence and Mg(2+) homeostasis. PLoS genetics 5, e1000428 (2009).
Stubben, C. J. et al. Steps toward broad-spectrum therapeutics: discovering virulence-associated genes present in diverse human pathogens. BMC genomics 10, 501 (2009).
Blanc-Potard, A. B. & Groisman, E. A. The Salmonella selC locus contains a pathogenicity island mediating intramacrophage survival. Embo J 16, 5376–5385 (1997).
Ford, D. C., Joshua, G. W., Wren, B. W. & Oyston, P. C. The importance of the magnesium transporter MgtB for virulence of Yersinia pseudotuberculosis and Yersinia pestis. Microbiology 160, 2710–2717 (2014).
Snavely, M. D., Miller, C. G. & Maguire, M. E. The mgtB Mg2+ transport locus of Salmonella typhimurium encodes a P-type ATPase. J Biol Chem 266, 815–823 (1991).
Smith, D. L., Tao, T. & Maguire, M. E. Membrane topology of a P-type ATPase. The MgtB magnesium transport protein of Salmonella typhimurium. J Biol Chem 268, 22469–22479 (1993).
Soncini, F. C., Garcia Vescovi, E., Solomon, F. & Groisman, E. A. Molecular basis of the magnesium deprivation response in Salmonella typhimurium: identification of PhoP-regulated genes. J Bacteriol 178, 5092–5099 (1996).
Groisman, E. A. The pleiotropic two-component regulatory system PhoP-PhoQ. J Bacteriol 183, 1835–1842 (2001).
Alix, E. & Blanc-Potard, A. B. MgtC: a key player in intramacrophage survival. Trends Microbiol 15, 252–256 (2007).
Alix, E. & Blanc-Potard, A. B. Peptide-assisted degradation of the Salmonella MgtC virulence factor. Embo J 27, 546–557 (2008).
Lee, J. W. & Lee, E. J. Regulation and function of the Salmonella MgtC virulence protein. Journal of microbiology 53, 667–672 (2015).
Lee, E. J. & Groisman, E. A. An antisense RNA that governs the expression kinetics of a multifunctional virulence gene. Mol Microbiol 76, 1020–1033 (2010).
Pavlov, M. Y. et al. Slow peptide bond formation by proline and other N-alkylamino acids in translation. Proc Natl Acad Sci USA 106, 50–54 (2009).
Doerfel, L. K. et al. EF-P is essential for rapid synthesis of proteins containing consecutive proline residues. Science 339, 85–88 (2013).
Ude, S. et al. Translation elongation factor EF-P alleviates ribosome stalling at polyproline stretches. Science 339, 82–85 (2013).
Blaha, G., Stanley, R. E. & Steitz, T. A. Formation of the first peptide bond: the structure of EF-P bound to the 70S ribosome. Science 325, 966–970 (2009).
Lee, E. J. & Groisman, E. A. Tandem Attenuators Control Expression of the Salmonella mgtCBR Virulence Operon. Mol Microbiol 86, 212–224 (2012).
Nam, D., Choi, E., Shin, D. & Lee, E. J. tRNAPro -mediated downregulation of elongation factor P is required for mgtCBR expression during Salmonella infection. Mol Microbiol. doi: 10.1111/mmi.13454 (2016).
Eriksson, S., Lucchini, S., Thompson, A., Rhen, M. & Hinton, J. C. Unravelling the biology of macrophage infection by gene expression profiling of intracellular Salmonella enterica . Mol Microbiol 47, 103–118 (2003).
Lee, E. J. & Groisman, E. A. Control of a Salmonella virulence locus by an ATP-sensing leader messenger RNA. Nature 486, 271–275 (2012).
Peil, L. et al. Distinct XPPX sequence motifs induce ribosome stalling, which is rescued by the translation elongation factor EF-P. Proc Natl Acad Sci USA. 110, 15265–15270 (2013).
Lassak, J., Wilson, D. N. & Jung, K. Stall no more at polyproline stretches with the translation elongation factors EF-P and IF-5A. Mol Microbiol 99, 219–235 (2016).
Starosta, A. L. et al. Translational stalling at polyproline stretches is modulated by the sequence context upstream of the stall site. Nucleic Acids Res 42, 10711–10719 (2014).
Lee, E. J., Pontes, M. H. & Groisman, E. A. A Bacterial Virulence Protein Promotes Pathogenicity by Inhibiting the Bacterium’s Own F1Fo ATP Synthase. Cell 154, 146–156 (2013).
Ohl, M. E. & Miller, S. I. Salmonella: a model for bacterial pathogenesis. Annu Rev Med 52, 259–274 (2001).
Navarre, W. W. et al. PoxA, yjeK, and elongation factor P coordinately modulate virulence and drug resistance in Salmonella enterica. Mol Cell 39, 209–221 (2010).
Zou, S. B., Roy, H., Ibba, M. & Navarre, W. W. Elongation factor P mediates a novel post-transcriptional regulatory pathway critical for bacterial virulence. Virulence 2, 147–151 (2011).
Lee, E. J., Choi, J. & Groisman, E. A. Control of a Salmonella virulence operon by proline-charged tRNAPro. Proc Natl Acad Sci USA 111, 3140–3145 (2014).
Fields, P. I., Swanson, R. V., Haidaris, C. G. & Heffron, F. Mutants of Salmonella typhimurium that cannot survive within the macrophage are avirulent. Proc Natl Acad Sci USA 83, 5189–5193 (1986).
Davis, R. W., Bolstein, D. & Roth, J. R. Advanced Bacterial Genetics. (Cold Spring Harbor Lab, 1980).
Snavely, M. D., Florer, J. B., Miller, C. G. & Maguire, M. E. Magnesium transport in Salmonella typhimurium: expression of cloned genes for three distinct Mg2+ transport systems. J Bacteriol 171, 4752–4760 (1989).
Datsenko, K. A. & Wanner, B. L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci USA 97, 6640–6645 (2000).
Acknowledgements
We thank Dongwoo Shin for providing strains. This work was supported, in part, by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT & Future Planning (NRF-2013R1A1A2074505 and NRF-2016R1A2B2012424) to E.-J.L. and NRF-2015R1A4A1041105 to J.-S.L. The authors declare no conflict of interest.
Author information
Authors and Affiliations
Contributions
E.-J.L. designed research; E.C., S.C., D.N., Y.H. and S.P. performed research; J.-S.L. managed mouse experiments; E.L. and E.C. wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Choi, E., Choi, S., Nam, D. et al. Elongation factor P restricts Salmonella’s growth by controlling translation of a Mg2+ transporter gene during infection. Sci Rep 7, 42098 (2017). https://doi.org/10.1038/srep42098
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep42098
This article is cited by
-
Whole-Genome Sequence and Pathogenicity Analysis of Providencia Heimbachae Causing Diarrhea in Weaned Piglets
Current Microbiology (2023)
-
Pat- and Pta-mediated protein acetylation is required for horizontally-acquired virulence gene expression in Salmonella Typhimurium
Journal of Microbiology (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
