
Abstract
We mapped the inferred long-distance migrations of four species of Amazonian goliath catfishes (Brachyplatystoma rousseauxii, B. platynemum, B. juruense and B. vaillantii) based on the presence of individuals with mature gonads and conducted statistical analysis of the expected long-distance downstream migrations of their larvae and juveniles. By linking the distribution of larval, juvenile and mature adult size classes across the Amazon, the results showed: (i) that the main spawning regions of these goliath catfish species are in the western Amazon; (ii) at least three species—B. rousseauxii, B. platynemum, and B. juruense—spawn partially or mainly as far upstream as the Andes; (iii) the main spawning area of B. rousseauxii is in or near the Andes; and (iv) the life history migration distances of B. rousseauxii are the longest strictly freshwater fish migrations in the world. These results provide an empirical baseline for tagging experiments, life histories extrapolated from otolith microchemistry interpretations and other methods to establish goliath catfish migratory routes, their seasonal timing and possible return (homing) to western headwater tributaries where they were born.
Similar content being viewed by others

Introduction
The Amazon has two main groups of migratory fish species, and they belong to the orders Siluriformes (catfishes) and Characiformes (characins)1,2,3. Major fisheries in the Amazon are based on knowledge of the seasonal upstream and downstream movements of fish, although the life cycles are poorly known by fishermen because the species enter and leave local fishing areas4,5,6. Long-distance fish migrations (>1,000 km) that exclusively or partially involve freshwater are known for salmon7 and eels8 but have also been inferred for Amazon goliath catfishes of the family Pimelodidae9,10,11,12. Although the migrations of most catfish species are poorly known, the general pattern reported is upstream movement to spawn, downstream passive drifting and even active migration of young juveniles to enter nursery habitats in river channels, floodplains or estuaries7,13,14,15. In the Amazon, goliath catfishes are major river channel and estuarine predators that are represented by a paraphyletic group of six extant and one fossil species of the genus Brachyplatystoma16,17,18 with a maximum known adult fork length (FL) of 60–280 cm9 (Fig. 1).

(A) B. vaillantii (piramutaba in Portuguese, pirabutón in Spanish); (B) B. rousseauxii (dourada in Portuguese, dorado in Spanish); (C) B. platynemum (babão in Portuguese, mota flemosa in Spanish); (D) B. juruense (zebra in Portuguese, zebra in Spanish); (E) Dorado migrations exploited by fishermen. The Santo Antônio Dam on the Madeira River now drowns the Teotônio Rapids (shown here) where B. rousseauxii (species in photo) and B. platynemum were previously exploited and easily detected when migrating. Photos by M. Goulding.
The first hypotheses of goliath catfish migration focused on B. rousseauxii and B. vaillantii, the most important commercial species, and their dependence on the Amazon River estuary as their nursery and inland rivers as feeding and spawning areas9. The spawning areas, however, were only identified as being located in the western Amazon, a vast region of at least 2 million km2 that includes parts of Colombia, Ecuador, Peru, Bolivia and Brazil. More recent biological studies based on commercial fisheries in all major western Andes-Amazon tributary basins19,20,21,22,23 established the seasonal, and in some cases year-round, presence of mature goliath catfishes in the western Amazon to at least a few hundred km downstream of the Andes. However, these investigations could not verify the spawning areas, leaving the possibility that they are located farther upriver and closer to the Andes.
We present data on the distribution of goliath catfish (Brachyplatystoma) adults, larvae and juveniles across the Amazon Basin, including areas in or near the Andes. Even without tagging experiments, the general differential distribution of sub-adult (downstream) and adult (upstream) goliath catfish size-classes9, as well as otolith microchemistry data10, strongly indicates that long-distance upstream goliath catfish migration occurs. Seasonal upstream goliath catfish movements are also visually obvious at some cataracts, such as those of the Madeira River (Fig. 1) in Brazil, before dams were built4, and those of the Caquetá River in Colombia6. Therefore, it is reasonable to hypothesize that long-distance downstream migrations of young fish occurs, otherwise there would be no recruitment to nurseries, in some cases as far downstream as the Amazon River estuary.
To test the goliath catfish migratory hypothesis related to western spawning regions, we mapped the presence of mature adults complemented by the seasonal and geographical variation in abundance and length of larvae and juveniles in river channels across the Amazon, including in the estuary. Two complementary geographical and temporal perspectives were used. The first focused on the Madeira Basin, the Amazon’s largest sub-basin with headwaters in the Andes, and where year-round collections were performed. The second included most of the Amazon and available data from all years. Our study shows that spawning for at least B. rousseauxii, B. juruense and B. platynemum occurs in or near the Andes and demonstrates conclusively that long-distance downstream migration of their larvae and juveniles occurs. The great variation in B. platynemum larvae size across the Amazon indicates a much wider nursery area than just the Andean region. Size-class data for B. vaillantii indicate long-distance migration and spawning in the western Amazon but do not confirm it near the Andes. Our results are also discussed in light of published goliath life history hypotheses derived from genetic and otolith isotope signatures.
Results
Adult Distribution and Gonadal Stage
Fish length and gonadal data derived from specimens captured in fisheries in major Andes-Amazon sub-basins in Brazil, Bolivia and Peru, complemented by published data from studies based in Colombia19,23 and Ecuador24, show the wide distribution of mature B. rousseauxii, B. platynemum and B. juruense in all major turbid rivers with headwaters in the Andes, including the Amazon River main channel (Fig. 2 and Table 1). Mature fish are defined as ripe individuals, that is, individuals with fully developed ova or testes25. Brachyplatystoma rousseauxii has the widest distribution. In addition to whitewater (turbid) rivers arising in the Andes, it is found in many clearwater and blackwater tributaries that arise on the Brazilian and Guiana Shields. With the exception of the lower Tocantins River26, which is part of the Amazon River estuary, B. rousseauxii is rarely registered in the fisheries of blackwater and clearwater rivers, an indication of its rarity in these drainages. The only Brazilian or Guiana shield rivers where mature long-distance migratory goliath catfishes were found was the Branco, a semi-turbid tributary of the Negro River. Of the goliath species considered, only B. rousseauxii was present in the Branco River, but it is of minimal importance in fisheries there and is reported by fishermen to be relatively rare.

(a) Sites investigated by the authors and others from published data. (b–e) Locations of mature goliath catfishes by species. Figure was created by authors with ArcGIS for Desktop Advanced 10.2, MapPublisher 9.6 tool inside Adobe Illustrator CC, and Adobe Illustrator CC, 2.0.
In contrast to the other three widely distributed species discussed in this paper, B. vaillantii is rare or naturally absent above the Madeira Rapids in the southwestern Amazon of Bolivia and Peru. The new fishways around the Madeira dams, however, could allow B. vaillantii to migrate more easily to Bolivian and Peruvian waters in the southwestern Amazon, which would alter the population dynamics of this species and perhaps other species on which it preys. Mature B. rousseauxii, B. platynemum and B. juruense are only abundant, as indicated by fisheries catches, between approximately 55–250 m of elevation and only in turbid rivers at least 3,500 km upstream from the Amazon River mouth (Fig. 2 and Table 1). Considering only adult stages, B. vaillantii has the widest distribution of the long-distance migratory goliath catfishes because of its presence in the far eastern Amazon, including the estuary9,27. In the estuary, however, B. vaillantii adults are not mature as defined above. Mature B. vaillantii adults are known as far upstream as the Pongo de Manseriche, an Andean gorge of the Marañón River at 246 m elevation in northern Peru and 4,847 km upstream of the Amazon River mouth. Year-round gonadal stage data reported for B. rousseauxii, B. platynemum and B. juruense captured over a 12-month period by local fishermen in the upper Madeira (2003–2004) and Ucayali (2004–2005) basins of Peru provided unequivocal proof of the presence of only mature goliath catfishes in or near the Andes to at least 198 m elevation and 5,788 km from the Amazon River mouth (Figs 2 and 3 and Table 2). Mature B. rousseauxii and B. platynemum were abundant in commercial fisheries in both areas during our sampling periods, whereas B. juruense was only captured in the Ucayali River. Other goliath catfishes (B. filamentosum, B. capapretum, and B. tigrinum) are also present in these rivers but in smaller quantities, as indicated by commercial fisheries28,29.

Monthly precipitation and monthly capture of mature B. rousseauxii, B. platynemum, and B. juruense between April 2002 and April 2004 in Puerto Maldonado (Madre de Dios River, Upper Madeira Basin) and between July 2004 and July 2005 at Atalaya (near the confluence of the Ucayali and Urubamba Rivers). B. juruense was not captured in the Madre de Dios River during our study period. Precipitation was used as a proxy for river level since there were few data for the latter in the Andean region.
All 5,348 B. rousseauxii, 2,985 B. platynemum and 528 B. juruense specimens we examined from commercial fisheries near the Andean Piedmont or in the immediate pre-Andean area were sexually mature adults with fully developed gonads or recently spawned fish (Table 2). In contrast to the downstream regions, all goliath catfish captured near the Andean Piedmont had empty stomachs, suggesting that their presence in these areas was not related to trophic migration but to spawning. Although goliath catfishes were present in every month in the far western Amazon, the commercial captures from which our data were derived indicated strong seasonal variation (Fig. 3). The seasonal differences between commercial captures in the Madre de Dios (upper Madeira Basin) and Ucayali and Urubamba Rivers (Ucayali Basin) (Fig. 3) may reflect distinct headwater migration patterns or differences in fishing, considering that fishing was virtually halted in the Madre de Dios during the high water discharge period (November-February) due to the large quantities of downstream-moving wood that are a danger to nets.
In the Ucayali and Urubamba Rivers, ready-to-spawn B. rousseauxii, B. juruense and B. platynemum were most abundant in commercial fisheries during the rainy and warmer months corresponding to the higher river discharge period from October to March (Fig. 3). Since monthly fishing effort was approximately equivalent, landings can be considered a proxy for seasonal migratory fish abundance. Furthermore, goliath catfishes were captured by commercial fisheries in river channels with downriver drifting gill nets during all months. Since drifting gill nets only capture fish moving upstream, they reveal the direction of movement.
Seasonal Larval Densities as Spawning Indicators
Only B. rousseauxii larvae were captured throughout most of the year in standardized density sampling, including during the low water period, but in much lower quantities than during the December floods (Fig. 4). The other two species present in the sampling area, B. platynemum and B. juruense, were poorly or not represented during 6–8 months in density samples that included parts of the high, low and rising water periods, and only the latter species was captured during the low water period. The highest larval densities of all species combined occurred between November and January, months when river discharge, depth and current velocity were rapidly increasing30. Mature B. platynemum were also fairly common in Madre de Dios commercial fisheries from May to August, which included the falling and low water period, verifying their presence in the region (Fig. 3). Although B. juruense larvae were captured between November 2004 and August 2005, no adults were reported in the commercial fisheries of the Madre de Dios for this period, although the species is known in the study area31 (Fig. 3).

The monthly drifting larvae densities of goliath catfishes in the Madre de Dios River of the upper Madeira Basin in relation to total monthly precipitation (mm) levels. Larvae densities were estimated based on ichthyoplankton samples from the Madre de Dios River between November 2004 and August 2005. Precipitation data were used as a proxy for river level since data for the latter were not available. Precipitation data are for the city of Puerto Maldonado were supplied by the Servicio Nacional de Meteorología e Hidrología del Peru (SENAMHI).
Growth of Larvae and Juveniles
If the goliath catfish spawning areas and nurseries are separated by thousands of kilometers as we hypothesize, it should be reflected in the larvae and juvenile growth in the downstream direction. We tested this hypothesis for the Madeira Basin, for which seasonally sequential data were available, and for the Amazon as whole, including data from various years. The growth of fish in their early phase may be described by Brody’s equation, which relates fish size (Lt) to time (t) in the exponential equation Lt = a × eK×t 32, where a and K are constants. We used Brody’s equation but substituted the time variable with distance to fit the relationship between larvae/juvenile length and their distance from the mouth of the Amazon River. The size distributions of B. rousseauxii and B. juruense in the Madeira Basin (Fig. 5) and the Amazon as a whole (Fig. 6) fit exponential curves (p < 0.01 for both species). By the time B. rousseauxii enters the Amazon River from the Madeira River, it has reached juvenile size (approximately 20 mm). For B. juruense, the available data indicate an increase in median length from the headwaters to the upper Madeira in Brazil, after which the median length decreases, indicating a wide spawning region for the species (Fig. 6). Juvenile stages of B. juruense are most common in the central Amazon, approximately 2,000–2,500 km from the Amazon River mouth, though much less is known about this species than the other goliath species considered and more collections are needed in the lower Amazon River, where juveniles are also known.

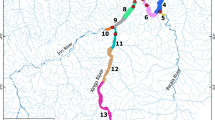
Box-plots of the lengths of goliath catfish drifting larvae from near the Andes in the Madre de Dios River to the Madeira River in Brazil, approximately 1,600 km downriver. Collection sites are indicated by the numbers above the box-plots: 1- Madre de Dios River at Los Amigos River confluence, 2- Madre de Dios River at Puerto Maldonado, 3- Madeira River at Porto Velho, and 4- Madeira River at Humaitá. Data were fit to an exponential curve, where Lmm is the size in millimeters and km is the distance in kilometers: B. rousseauxii: Lmm = 269.1−0.00101×km, r2 = 0.67, F(1, 327) = 670, p < 0.001; B. juruense: Lmm = 18.5−0.00034×km, r2 = 0.22, F(1, 76) = 23.1, p < 0.001; B. platynemum: r2 = 0.17 F(1, 108) = 1.93 p > 0.05; B. vaillantii: r2 = 0.08, F(1, 34) = 2.9, p > 0.05.

Fish length scatterplots of goliath catfish drifting larvae from near the Andes to the Amazon River estuary in Brazil. Data were fit to an exponential curve, where Lmm is the size in millimeters and km is the distance in kilometers: B. rousseauxii: Lmm = 81.8−0.00065×km, r2 = 0.84, F(1, 491) = 670, p < 0.001; B. juruense: Lmm = 33.9−0.00048×km, r2 = 0.57, F(1, 130) = 172.8, p < 0.001; B. platynemum: r2 = 0.04, F(1, 151) = 6.3, p > 0.05; B. vaillantii: Lmm = 50.9−0.00085×km, r2 = 0.64, F(1, 1035) = 1818, p < 0.001. Maps in figure was created by authors with ArcGIS for Desktop Advanced 10.2, MapPublisher 9.6 tool inside Adobe Illustrator CC, and Adobe Illustrator CC, 2.0.
A seasonal factor for all species is that varying current speeds during sampling periods could affect the larval size distribution along the downstream movement. If current speed greatly influenced the distribution of juvenile size, then a much less precise pattern would be expected for B. rousseauxii since it spawns during varying periods of the year with different current speeds. However, B. rousseauxii shows the best fit of all species to the downstream migratory growth model.
The wide distribution of small larvae of B. platynemum in the Madeira Basin and the Amazon as a whole and their highly mixed length-class distribution do not fit exponential curves, strongly suggesting that spawning for this species is widespread, that more than one population exists33 and that the nurseries include a large area in the western and central, and perhaps even eastern, Amazon (Figs 5 and 6).
Larvae and juveniles of B. vaillantii have been captured in the lower stretches of the Madeira River and upriver near the Madeira Rapids, at least 3,129 km upstream of the estuary. The presence of small larvae (<5 mm) in this region suggests the proximity of a spawning area, but we have too few sample points to detect the growth of B. vaillantii larvae during their downriver migration (Fig. 5). Nevertheless, the data for B. vaillantii larvae and juveniles for the entire Amazon fit an exponential curve (p < 0.01), corroborating the long-distance downstream larval and juvenile migration hypothesis (Fig. 6). Given the presence of ripe adults of this species in the Marañón River in Peru at its outlet from the Andes, it is possible that some spawning occurs much farther upstream in the western Amazon than our data indicate. No small B. vaillantii larvae (<5 mm) were found in the Amazon River within 1,500 km of the estuary.
Discussion
We argue that the distribution of mature size classes and general downriver movement, concurrent growth and regional size-class differences of goliath catfish larvae and juveniles of B. rousseauxii, B. juruense and B. vaillantii indicate that the western spawning areas and downstream nurseries of these species are widely separated in the Amazon (Figs 2, 3 and 6). The greatest distances measured between spawning and nursery areas, as determined by larvae and juvenile presence, were 5,786 km for B. rousseauxii, 4,238 km for B. juruense and 3,129 km for B. vaillantii.
The most extreme migration is undertaken by B. rousseauxii, which spawns in the far western Amazon but uses the estuary as its nursery, for a maximum known life history migratory cycle of all size classes of approximately 11,600 km (Figs 5 and 6). The size-class distribution of older juveniles and adults indicates that it takes 1–2 years to reach the Andes during upstream migration from the estuary9,22,34. Based on B. rousseauxii size classes recorded from fisheries catches in the western Amazon19,22 and farther downstream9, adults do not return to the estuary but remain in a large area of the western Amazon. The annual migration of adults subsequent to reaching the western Amazon for the first time, however, is probably much shorter; however, based on the distribution of adult size classes, it could still be 1,000–2,000 km or more. Because no other strictly freshwater long-distance fish migrations7 close to those discussed in this paper have been reported, B. rousseauxii undertakes the longest migration in the world, considerably longer than previously hypothesized9. Brachyplatystoma rousseauxii migration also surpasses the maximum life cycle migration (6,000 km) reported for anadromous salmon (Oncorhynchus)35 and is nearly as long as that of the European eel (Anguilla anguilla Linnaeus 1758), including the freshwater and marine phases8. It is also possible that B. platynemum undertake migration from the estuary to the Andean region similar to B. rousseauxii, as juveniles of the former are relatively common in artisanal fisheries in the estuary36, but much less is known about them37.
The general life history patterns of B. platynemum and B. juruense are similar to those of B. rousseauxii, with a more restricted, but still wide, separation of spawning and nursery areas. The more mixed larvae and juvenile size classes of B. platynemum across the Amazon indicate that their spawning areas are neither exclusively in the far western Amazon nor are their nurseries restricted to or mostly in the eastern Amazon. In contrast to B. rousseauxii and B. platynemum, neither adult nor larval B. juruense are known in the estuary, and the nursery of the latter species appears to be in the central Amazon. Of the long-distance migratory goliath catfishes, B. vaillantii is the only species whose adults and young share the estuary. To date, the maximum distance from the estuary that B. vaillantii larvae have been captured is 3,129 km. However, the presence of mature B. vaillantii adults in the Marañón River at the Pongo de Manseriche Gorge (4,847 km upstream) and in the Napo of Ecuador (4,754 km upstream) suggests the possibility of spawning in the far western Amazon. Based on commercial catches, B. vaillantii is most abundant in the Amazon River mainstem and was rare, if at all present, in Bolivia and Peru above the Madeira Rapids in Brazil. As mentioned above, the new dam bypasses might allow the species to pass the Madeira Rapids, which were previously a barrier4.
The presence and abundance of mature goliath catfish in commercial fisheries in the Andean region is a reliable indicator of upstream movement to spawn. The absence of these fish in commercial fisheries during some or all months near the Andes, however, should not be interpreted as direct evidence that they are not present in the region, as fishing activity must also be considered. The striking differences between the monthly relative captures in the Madre de Dios and Ucayali-Urubamba Rivers is most likely due to the absence of fishing in the former rather than the absence of mature fish. The high catches in the Madre de Dios River during the falling river level period during all three years for which data are available should not be interpreted as a greater abundance at this time of year but rather that fishing is possible during these months (Fig. 3). The presence of spawning B. rousseauxii and B. platynemum in the Madre de Dios River channel during the high water period is corroborated by the relative abundance of their larvae (Fig. 4). The higher density of fish larvae during the rising water period is thus a better indicator of the main reproductive period of goliath catfishes than monthly commercial captures. The best indicator, however, is larval flux, which is the absolute density value of larvae per unit of time in a river section38. Drift densities of young fish generally decrease with increased river depth and flow velocity due to the dilution effect caused by higher turbulence in a much greater volume of water39. The larval flux index was not used due to the difficulty of obtaining accurate river discharge data near the Andes because of the few hydrological stations that exist. Considering that goliath catfish larvae densities were calculated without discharge calculations and were highest at the beginning of the rainy season and decreased during the January and March peak discharge periods30, it is possible that the highest larval flux would be in the latter period if river discharge was considered in the larvae density algorithms.
Fisheries data from the middle Caquetá River in Colombia also indicate that upstream-moving mature goliath catfish are most abundant during the high water period40. While upstream areas near the Andes have yet to be investigated as spawning sites in the Caquetá Basin, large B. rousseauxii have been reported farther upstream to at least 170 m elevation in the neighboring Putumayo River23. In contrast to our data and that of the Caquetá, a gonadal review of 15,000 B. rousseauxii specimens captured by commercial fisheries during a 5-year period near Iquitos, Peru at 76 m elevation and approximately 4,000 km upstream of the Amazon River mouth hypothesized that breeding occurs mostly during the low water period and ends at the beginning of rising water22. The fisheries in both areas are intensive throughout the year, and fishing effort bias is minimal22,40. However, those studies analyzed the monthly proportions of mature females by combining maturation stages 3 (advanced maturation) and 4 (ripe)25. The uncertainty of the time between stages 3 and 4 raises doubts about combining the two to identify exactly when spawning occurs. It is unknown how far downstream of the Andes or immediate pre-Andean area that long-distance migratory goliath catfish spawn, and the Caquetá and Iquitos data alone do not confirm this. Nevertheless, the presence of maturing populations throughout the year in the Iquitos Region, an area of large rivers and floodplains where prey are more abundant9 (Fig. 2, Table 1), suggests that migratory adult goliath fish, especially B. rousseauxii, remain in this area to feed and develop their gonads for subsequent reproductive cycles after their first arrival.
A second study in the Iquitos area that included only larval abundance concluded that breeding occurs mostly during receding or low water41. Considering that spawning fish are present during all months in the Ucayali headwaters upriver of this site (Fig. 3), it is reasonable to expect the presence of fish larvae downstream near Iquitos during all months. The larval results are based only on abundance without reference to water volume, the latter of which is needed to calculate the absolute abundance index to compare high and low water periods.
A similar but slightly later peak in drifting larval abundance than that of the Madre de Dios River was found for B. rousseauxii in the Madeira River, 1,613 km downstream from the former sampling site, with maximum fluxes (larvae or juveniles/second) in January and February and minimal downstream migration in September38. Another study using bottom-trawl sampling in the same region of the Madeira River reported different results for B. rousseauxii, with relatively high larvae and juvenile abundance (number of fish/haul) during the low water period42. However, the study did not consider the bias introduced by the inverse effect of decreased water volume on larval density; thus, seasonal comparisons may not be accurate because the absolute number of larvae may be greater during the high water period but in a much greater volume of water.
As indicated by the upper watershed areas in the Madeira and Ucayali Basins, the goliath catfish spawning zone is located in a mountainous to lowland transition area above about 170 m with relatively high channel slope (declivity = 0.16 m/km) and a lowland downstream area with a much lower declivity (generally less than 0.01 m/km) (Table 1). The river channels in the spawning zone, and in the Andes in general below 300 m, are characterized by gravelly bottoms, as opposed to muddy and soft substrates farther downstream, greater turbidity than downstream, shallower river channel depth (7–20 m versus 40 + m downstream), higher pH (up to 7.9 versus up to 7.1 downstream), higher conductivity (up to 287 microsiemens/cm versus 70 microsiemens/cm downstream), highly saturated O2 levels (up to 8.2 mg/l versus up to 6.4 mg/l downstream), and lower average water temperature (26.8 °C versus 28 °C downstream)20,43. Despite these striking physical differences, there are too few data to support a hypothesis of spawning site selection by long-distance migratory goliath catfishes. Furthermore, limnological and geomorphological data alone cannot be used as explanatory variables because biological factors, such as avoidance of egg and larvae predators, could also play a critical role9.
The evolution of long-distance migratory goliath catfish life histories in connection with the western Amazon and western Orinoco, where they also occur, could reflect an ancient evolutionary spawning association with the Andes, as Andean fossils of their genus are known from at least the Miocene 12–11 Ma16, although the elevations at that time were probably not much higher than 200 m. The paleo-Amazon-Orinoco in which the ancient catfish species lived flowed north in the low foreland basin of the Andes44. Brachyplatystoma vaillantii, B. rousseauxii, and B. platynemum range widely beyond the Amazon, with all three found throughout the Orinoco Basin and the first two also in the large, short rivers of the Guianas. Juveniles have also been reported in river channels of the Orinoco Basin45.
The geographical genetics of three goliath catfish species have been studied, but only B. platynemum presented clear population segregation for the Amazon River mainstem and Madeira River33, whereas B. vaillantii showed no genetic segregation46,47. Genetic studies of B. rousseauxii presented mixed results but indicated a relatively homogenous population in the Amazon Basin, although homing behavior could not be totally excluded48,49. Regional genetic differentiation of B. rousseauxii in the large headwater region of the upper Madera Basin in Bolivia could also indicate homing behavior49. The present genetic evidence indicates that long-distance migratory goliath catfish species have few distinct populations in the Amazon Basin, but it does not convincingly eliminate the possibility of homing.
Recent studies have reconstructed the theoretical movement and migration of individuals of the Brachyplatystoma species by comparing strontium isotope signatures (87Sr/86Sr) along transverse sections of their otoliths with the isotope signatures of major river water types10,11. The main results show that the juvenile phases of B. rousseauxii and B. vaillantii have strontium isotope signatures of the western Andean tributaries and the Amazon River mainstem, and few present signatures for other water types. The estimated mean size (37 ± 16 mm (mean ± S.D.) of B. rousseauxii when it reaches the Amazon River during its downriver drift migration in the Madeira Basin has also been calculated using strontium isotope signatures and the relationship between otolith radius and body length10. The length range of B. rousseauxii larvae and juveniles captured in the Madeira River in our study varied from 5 to 42 mm (Fig. 5), in agreement with that extrapolated from otoliths. The otolith data corroborate that the nursery habitat of these two species is mostly in turbid rivers, as directly indicated by our study and indirectly indicated by fisheries data9. Although considering the eastern Amazon as an obligate region of early goliath catfish life cycles, otolith isotope interpretations also point to the possibility that some of the large eastern clearwater tributaries might be used as nurseries for B. rousseauxii and B. vaillantii. The eastern estuary (Marajó Bay, Pará River and the lower Tocantins) is heavily influenced by the clearwater Tocantins River, especially during the Amazon River’s low water period50; thus, it should be expected that at least part of the estuary goliath catfish nursery population would have a clearwater river otolith strontium signature. Our field studies do not indicate the presence of young (<20 cm) B. rousseauxii and B. vaillantii in the Xingu or Tapajós Rivers, and they are absent or extremely rare in the fisheries of those areas. A multi-element otolith microchemistry study also indicated that B. rousseauxii resided in the Amazon estuary for the first 1.5–2.0 years of life based on Sr:Ca and Ba:Ca ratios12.
By presenting the convergence of three lines of evidence—the distribution of mature size classes, downstream migration of larvae/juveniles and otolith signatures—that strongly suggest long-distance goliath catfish migration in the Amazon, this study presents a significant step towards achieving a holistic understanding of the longest freshwater fish migrations in the world and at a mega-basin scale. Furthermore, it sets the hypothetical stage for eventual tagging experiments to understand the exact environmental cues that the fishes use during various life history movements and to test empirically recent homing hypotheses10 and at what sub-basin level they occur. The methods used in this paper also raise uncertainties that the present data alone cannot address, such as why the goliath catfishes that use the eastern or central Amazon as nurseries migrate thousands of kilometers upstream to spawn in the far western Amazon associated with the Andes or nearby uplands rather than reproduce much closer in the middle or even lower reaches of the Amazon River mainstem or in the eastern tributaries arising on the Brazilian and Guiana Shields.
Finally, the previous limitations imposed by inferring migration based on the wide separation of nurseries and spawning areas are largely eliminated in this paper by proof of long-distance downstream larvae/juvenile movements in the river channels. Of special relevance is the expected infrastructure development in the Andes, especially the combination of dams, headwater deforestation and mining activity51,52, which could present major threats to important spawning areas ranging from Colombia in the north to Bolivia in the south. Andean large dams will most likely be much different than those already constructed elsewhere in the Amazon, specifically their high walls. Even if high-wall dams are located upstream of spawning sites, they would greatly alter sediment and nutrient cycles downriver where spawning occurs. The long-distance migratory goliath catfishes provide a profound biological indicator of ecosystem health from the Andes to the freshwater Amazon River plume in the Atlantic, and the impacts on them should be considered in all major infrastructure development.
Methods
Definition of Long-Distance Migratory Goliath Catfishes
A modern cladistic classification of the goliath catfish genus Brachyplatystoma includes seven species17, of which five of the extant six species are easily recognized by commercial fishermen. We consider five (rousseauxii, vaillantii, platynemum, juruense and tigrinum) to be long-distance (>1,000 km) migratory species. We did not include B. tigrinum in this study because it is relatively rare in fisheries, and our data for it were minimal, although small numbers of individuals are commonly present in association with the upstream movement of B. rousseauxii. The largest goliath catfishes are B. filamentosum and B. capapretum, both commonly reaching more than 2.5 m in length and 100 kg in weight. We found no field evidence that they perform long-distance migration (>1,000 km), and their mixed size classes across the Amazon indicate that they spawn widely and in various river types, although shorter migrations are probably involved12. They are also the only species that commonly enter floodplain waters53, whereas the long-distance migratory species are largely confined to river channels and the estuary.
Adult Goliath Catfish Distribution and Size Classes
Data for the presence of migratory goliath catfishes, their size classes and maturity stages were obtained by extensive field surveys of local fisheries conducted by the authors since the late 1970 s in six major regions: (1) the Amazon River mainstem from its estuary in Brazil to the confluence of the Ucayali and Marañón Rivers in Peru; (2) major tributary basins (Madeira, Ucayali and Marañón) with headwaters in the Andes; (3) the Purus and Juruá Basins, whose headwaters are associated with the Fitzcarrald Arch, which has a post-Andean uplift origin that produced their low hilly areas, such as the Serra do Divisor54; (4) Guiana Shield basins (Negro, Branco and Trombetas); (5) Brazilian Shield basins (Trombetas, Xingu and Tapajós); and (6) the Amazon estuary, including Marajó Bay and offshore fresh waters in the Atlantic. The data from the Andean tributaries in Colombia and Ecuador were obtained from the literature. Published goliath catfish data, including fisheries catches, maturity stages and size classes, were especially important for the Caquetá and Putumayo basins, which have been investigated by Colombian scientists19,23. Photographic proof and coordinates of goliath catfishes for the Napo Basin near the Andes was provided in a recent thesis24.
The presence of the migratory goliath catfishes and their maturity stages were verified by the authors from specimens in urban markets and complemented by local interviews. The maturation stages of females and males were based on macroscopic gonadal characteristics: ripe, ready-to-spawn individuals, spent or very recently spent25. Wherever present, goliath catfishes are known to the local people, and nearly all oral reports we received were eventually verified by actual specimens either from fishers or our own observations. Mature fish were considered abundant in a fishing area when they were among the 20 most important food species, frequent when they regularly appeared in markets in small quantities, and rare when observed by the authors but their presence was not well known by local fishers.
Andean surveys based on interviews to establish the maximum elevations that goliath catfishes reach were conducted in the Amazon River’s two largest Andes-Amazon sub-basins, the Madeira and Ucayali. For the Madeira Basin, the Madre de Dios sub-basin in Peru was surveyed from the city of Puerto Maldonado in the lowlands (<200 m) to tributaries 4,000 m upstream, including the Manu, Alto Madre de Dios and Inambari Rivers, whose headwaters rise in the high Andes. Surveys in the Ucayali Basin ranged from near Cusco at 3,300 m to Atalaya at 200 m and included the Vilcanota, Urubabamba, Tambo and Ucayali Rivers. No evidence of long-distance migratory goliath catfishes was found above 250 m elevation in the Andes (Table 1).
Two large datasets of unpublished goliath catfish fork length and gonadal maturity stages are reported here for the first time: Madre de Dios River commercial fisheries based in the city of Puerto Maldonado between April 2002 and April 2004 in the Upper Madeira Basin of Peru and Urubamba and Ucayali commercial fisheries between July 2004 and August 2005 based in the city of Atalaya (Table 2). The Madre de Dios fisheries are within 150 km of the Andes, and those of Atalaya are at the edge of the Andes.
Ichthyoplankton Data
Given the lack of tagging experiments to detect upstream movements in the days or weeks before spawning, a more substantial understanding of goliath catfish reproductive periods in the western Amazon is provided by the seasonal downstream drift of their larvae, which indicates when spawning occurs. We define larvae as the fish phase between hatching to complete loss of embryonic and larval organs55, with the juvenile stage beginning at approximately 20 mm56. Fish larvae larger than 2.7 mm were measured, and they were identified by the number of caudal and pre-caudal myomeres56. Year-round larvae and juvenile collections were conducted in the Madeira, the Amazon’s largest sub-basin, and these results were compared to all the data available for the Amazon, including all specimens captured in various years, seasons and locations. For the Madeira Basin, the Madre de Dios River in southeastern Peru was selected as the site near the Andes to test for the presence of headwater larvae because commercial fisheries based in the city of Puerto Maldonado revealed the at least seasonal presence of mature goliath catfishes near the Andes20.
Total length and abundance data of goliath catfish larvae and juveniles were investigated near the Andes in the Madre de Dios, Ucayali and Urubamba Rivers; in the lowlands in the large channels of the Madeira and Amazon rivers; and in the freshwater areas of the Amazon River estuary. Samples were collected during daylight hours with ichthyoplankton nets with a mesh size range of 300–500 μm and trawl nets with a mesh size of 5 mm30,56,57. The fish larvae collection method was approved by the Brazilian Institute of the Environment - IBAMA (SISBIO- permission number 4419) in Brazil and the Dirección General de Extracción y Producción Pesquera para Consumo Humano Directo from the Ministry of Production in Peru, with in-country legal deposit protocols of the Museo de Historia Natural, Lima.
Standardized larval density data30 were collected weekly using ichthyoplankton nets in the Madre de Dios River between November 2004 and August 2005 (Fig. 4). The sampling was conducted at five points across five cross-channel transects along 12 km of the river. Two samples were taken at each site: near the surface (1 m depth) and near the bottom (70% of the maximum depth). A mechanical flowmeter (General Oceanics 2030 R) was installed at the mouth of the net to estimate the volume of water sampled. The monthly drifting larvae densities were estimated based on the average of all samples.
River Distance Calculations
The distances from the Amazon River mouth to the studied areas were determined using the Barrier Analysis Tool (BAT) as an extension of ArcMap 10.2, which was developed for The Nature Conservancy (Software Developer: Duncan Hornby of the University of Southampton’s GeoData Institute). The tool uses point data to divide a routed river network (polylines with node to node coding) into connected networks from which a direct path distance calculation can be made.
To test the long-distance downstream larvae and juvenile migration hypothesis, the size of larvae and juveniles in the ichthyoplankton samples from the Madre de Dios, Madeira and Amazon Rivers and the estuary were related to the distance of the sampling points to the Amazon River mouth by the exponential equation L = a × eb×D, where L is the size in millimeters and D is the distance in kilometers from the Amazon River mouth. An exponential model was chosen because it had biological significance and provided the best data fit.
Climate Data
Hydrological data were not available for most of the Andean areas we surveyed; thus, we used local precipitation data as a proxy for river level seasonality. Precipitation data were supplied by the Servicio Nacional de Meteorología e Hidrología del Peru (SENAMHI). Because there were no precipitation records for Puerto Maldonado (Madre de Dios) in November 2004, we used the average November precipitation registered in a historical series of 43 years.
Note
Herewith the authors declare that the study (manuscript no: SREP1622389T) was completed in accordance with the laws of Brazil and Peru and the fish larvae collection method was approved by the Brazilian Institute of the Environment - IBAMA (SISBIO- permission number 4419) in Brazil and the Dirección General de Extracción y Producción Pesquera para Consumo Humano Directo from the Ministry of Production in Peru, with in-country legal deposit protocols of the Museo de Historia Natural, Lima.
Additional Information
How to cite this article: Barthem, R. B. et al. Goliath catfish spawning in the far western Amazon confirmed by the distribution of mature adults, drifting larvae and migrating juveniles. Sci. Rep. 7, 41784; doi: 10.1038/ srep41784 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Barthem, R. & Goulding, M. An Unexpected Ecosystem: The Amazon as Revealed by Fisheries. (Gráfico Biblos & Missouri Botanical Garden, 2007).
Goulding, M. In Tropical Rainforests: Diversity and Conservation (eds F. Almeda & C. M. Pringle ) 71–86 (California Academy of Sciences, 1988).
Araujo-Lima, C. A. R. M. & Ruffino, M. L. In Migratory Fishes of South America: Biology, Fisheries and Conservation Status (eds J. Carolsfield, B. Harvey, C. Ross & A. Baer ) 62–75 (World Fisheries Trust/World Bank/IDRC, 2004).
Goulding, M. Man and Fisheries on an Amazon Frontier. 121 (Kluwer Academic Publishers/Springer-Verlag, 1981).
Ribeiro, M. C. L. B. & Petrere-Jr, M. Fisheries ecology and management of the jaraqui (Semaprochilodus taeniurus, S. insignis) in Central Amazonia. 5, 195–215 (1990).
Rodríguez-Fernandez, C. A. Bagres, Malleros y Cuerderos en El Bajo Río Caquetá. Estudios en la Amazonía Colombiana. Vol. 2 (Tropenbos-Colombia, 1991).
Lucas, M. C. & Baras, E. Migration of Freshwater Fishes. 440 (Blackwell Science Ltd, 2001).
Aarestrup, K. et al. Oceanic spawning migration of the European eel (Anguilla anguilla). Science 325, 1660 (2009).
Barthem, R. B. & Goulding, M. The Catfish Connection: Ecology, Migration, and Conservation of Amazon Predators. (Columbia University Press, 1997).
Duponchelle, F. et al. Trans-Amazonian natal homing in giant catfish. J. Appl. Ecol., doi: 10.1111/1365-2664.12665 (2016).
Hegg, J. C., Giarrizzo, T. & Kennedy Brian, P. Diverse early life-history strategies in migratory Amazonian catfish: Implications for conservation and management. PLoS ONE 10 (2015).
Hermann, T. W., Stewart, D. J., Limburg, K. E. & Castello, L. Unravelling the life history of Amazonian fishes through otolith microchemistry. Royal Society Open Science 3 (2016).
Carolsfeld, J., Harvey, B., Ross, C. & Baer, A. (International Development Research Center, Ottawa, Canada, 2003).
Cowx, I. G. et al. Vol. 49 (ed Mekong River Commission) 100 (MRC Technical Paper, Phnom Penh, 2015).
Hogan, Z., Baird, I. G., Radtke, R. & Vander-Zanden, M. J. Long distance migration and marine habitation in the tropical Asian catfish, Pangasius krempfi. J. Fish. Biol. 71, 818–832 (2007).
Lundberg, J. G. Brachyplatystoma promagdalena, new species, a fossil goliath catfish (Siluriformes: Pimelodidae) from the Miocene of Colombia, South America. Neotrop. Ichthy. 3, 597–605 (2005).
Lundberg, J. G. & Akama, A. Brachyplatystoma capapretum: A new species of goliath catfish from the Amazon Basin, with a reclassification of allied catfishes (Siluriformes: Pimelodidae). Copeia 2005, 492–516 (2005).
Lundberg, J. G., Sullivan, J. P. & Hardman, M. Phylogenetics of the South American catfish family Pimelodidae (Teleostei: Siluriformes) using nuclear and mitochondrial gene sequences. Proc. Acad. Nat. Sci. Philadelphia 161, 153–189 (2011).
Agudelo-Córdoba, E. et al. Breeding, growth and exploitation of Brachyplatystoma rousseauxii Castelnau, 1855 in the Caquetá River, Colombia. Neotrop. Ichthyol. 11, 637–647 (2013).
Barthem, R., Goulding, M., Forsberg, B., Cañas, C. M. & Ortega, H. Aquatic Ecology of the Rio Madre de Dios: Scientific Bases for Andes-Amazon Headwaters Conservation. (Asociación para La Conseración de La Cuenca Amazónica (ACCA)/Amazon Conservation Association (ACA), 2003).
Coronel, J. S., Maes, G. E., Claus, S., Damme, P. A. V. & Volckaert, F. A. M. Differential population history in the migratory catfishes Brachyplatystoma flavicans and Pseudoplatystoma fasciatum (Pimelodidae) from the Bolivian Amazon assessed with nuclear and mitochondrial DNA markers. J. Fish Biol. 65, 859–868 (2004).
Garcia-Vasquez, A. et al. Life-history characteristics of the large Amazonian migratory catfish Brachyplatystoma rousseauxii in the Iquitos Region, Peru. J. Fish Biol. 75, 2527–2551, doi: 10.1111/j.1095-8649.2009.02444.x (2009).
Lasso, C. A. et al. In Serie Editorial Recursos Hidrobiológicos y Pesqueros Continentales de Colombia. 359 (Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá, Colombia, 2011).
Utreras-Bucheli, V. M. Caracterización de la pesca de grandes bagres en el Alto Río Napo (Ecuador), recomendaciones para su manejo y conservación Master’s Thesis thesis, Universidad Internacional de Andulucía (UNIA) (2010).
Núñez, J. & Duponchelle, F. Towards a universal scale to assess sexual maturation and related life history traits in oviparous teleost fishes. Fish. Phys. Biochem. 35, 167–180 (2009).
Merona, B. d., Juras, A. A., Santos, G. M. d. & Cintra, I. H. A. Os peixes e a pesca no baixo rio Tocantins: Vinte anos depois da UHE Tucuruí. 208p (Centrais Elétricas do Nort e do Brasil S.A – Eletrobr as Eletronort e, 2010).
Barthem, R. B. Ocorrência, distribuição e biologia dos peixes da baía de Marajó, estuário amazônico. Bol. Mus. paraense Emílio Goeldi 2, 49–69 (1985).
Cañas, C. M. La pesca en la provincia de Tambopata: Monitoreo y ordenamiento pesquero en el sureste peruano. 178–186 (Proyecto Aprovechamiento y Manejo Sostenible de la Reserva de Biosfera y Parque Nacional del Manu (PRO-MANU), Lima, Peru, 2000).
Riofrío-Quijandria, J. C. & Palma-Gonzales, C. Evaluación pesquera en Atalaya y alrededores (Ucayali). (Informe Técnico presentado a ERM, 2003).
Cañas, C. M. & Pine, W. E. Documentation of the temporal and spatial patterns of Pimelodidae catfish spawning and larvae dispersion in the Madre de Dios River (Peru): Insights for conservation in the Andean-Amazon headwaters. River Res. Appl. 27, 602–611, doi: 10.1002/rra.1377 (2011).
Cañas, C. Evaluación de los recuros pesqueros en la proviincia de Tambopata, Madre de Dios. Conservation International, Peru, Série Técnica 67, 1–34 (2000).
Ricker, W. E. Computation and interpretation of biological statistics of fish populations. Bull. Fish. Res. Board Canada 191, 1–382 (1975).
Ochoa, L. E. et al. Genetic structure and historical diversification of catfish Brachyplatystoma platynemum (Siluriformes: Pimelodidae) in the Amazon basin with implications for its conservation. Ecol. Evol. 5, 2005–2020 (2015).
Alonso, J. C. & Pirker, L. E. M. In O Manejo da Pesca dos Grandes Bagres Migradores (eds N. N. Fabré & R. B. Barthem ) 21–28 (Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis 2005).
Cooke, S. J., Crossin, G. T. & Hinch, S. G. In Encyclopedia of Fish Physiology: From Genome to Environment Vol. 3 (ed A.P. Farrell ) 1945–1952 (Academic Press, 2011).
Barthem, R. B., Ribeiro, M. & Petrere-Jr., M. Life strategies of some long-distance migratory catfish in relation of hydroelectric dams in the Amazon Basin. Biol. Conser. 55, 339–345 (1991).
Klautau, A. G. C. M. et al. Impacted biodiversity by industrial piramutaba fishing in the Amazon River mouth. Bol. Inst. Pesca 42, 102–111 (2016).
Barthem, R. B., Costa, M. C., Cassemiro, F., Leite, R. G. & Silva-Jr., N. In Biodiversity: The Dynamic Balance of the Planet (ed O. Grillo ) 137–158 (InTech, 2014).
Lechner, A. et al. Hydraulic forces impact larval fish drift in the free flowing section of a large European river. Ecohydrology 7, 648–658 (2014).
Agudelo-Córdoba, E. et al. Bagres de la Amazonia Colombiana: Un Recurso sin Fronteras. (SINCHI, 2000).
García-Dávila, C. et al. Using barcoding of larvae for investigating the breeding seasons of pimelodid catfishes from the Mara~non, Napo and Ucayali rivers in the Peruvian Amazon. J. Appl. Ichthyol. 31, 40–51 (2016).
Cella-Ribeiro, A. et al. Temporal and spatial distribution of young Brachyplatystoma spp. (Siluriformes: Pimelodidae) along the rapids stretch of the Madeira River (Brazil) before the construction of two hydroelectric dams. J. Fish Biol. 86, 1429–1437 (2015).
Devol, A. H., Forsberg, B., Richey, J. & Pimentel, T. P. Seasonal variation in chemical distributions in the Amazon (Solimões) river: A multiyear time series. Glob. Geochem. Cycle 9, 307–328 (1995).
Wesselingh, F. P. & Hoorn, C. In Historical Biogeography of Neotropical Freshwater Fishes (eds James Albert & Roberto E. Reis ) 59–67 (University of California Press, 2007).
Machado-Allison, A. In Reproductive Biology of South American Vertebrates 45–59 (Springer, 1992).
Formiga-Aquino, K. et al. In Biología de las Poblaciones de Peces Amazónicos y Piscicultura. Comunicaciones del Segundo Coloquio Internacional de la Red de Investigación sobre la Ictiofauna Amazónica. (eds Jesús Núñez, Fred Chu-Koo, Jorge Porto & Carmen R. García-Dávila ) 37–46 (IRD) (2011).
Rodrigues, F. C., Farias, I. P., Batista, J. & Alves-Gomes, J. A. Isolation and characterization of microsatellites loci for “piramutaba” (Brachyplatystoma vaillantii, Siluriformes: Pimelodidae), one of the commercially most important migratory catfishes in the Amazon Basin. Cons. Gen. Res. 1, 365–368 (2009).
Batista, J. S. & Alves-Gomes, J. A. Phylogeography of Brachyplatystoma rousseauxii (Siluriformes - Pimelodidae) in the Amazon Basin offers preliminary evidence for the first case of “homing” for an Amazonian migratory catfish. Gen. Mol. Res. 5, 723–740 (2006).
Carvajal-Vallejos, F. M. et al. Genetic structure in the Amazonian catfish Brachyplatystoma rousseauxii: influence of life history strategies. Genetica 142, 323–336, doi: 10.1007/s10709-014-9777-2 (2014).
Barthem, R. B. & Schwassmann, H. O. Amazon river influence on the seasonal displacement of the salt wedge in the Tocantins River Estuary, Brazil, 1983–1985. Bol. Mus. par. Emílio Goeldi, Zool. 10, 119–130 (1994).
Finer, M. & Jenkins, C. N. Proliferation of hydroelectric dams in the Andean Amazon and implications for Andes-Amazon connectivity. PLoS One 7, e35126, http://www.ncbi.nlm.nih.gov/pubmed/22529979 (2012).
Finer, M., Jenkins, C. N., Pimm, S. L., Keane, B. & Ross, C. Oil and gas projects in the western Amazon: Threats to wilderness, biodiversity, and indigenous peoples. PloS one 3, e2932, doi: 10.1371/journal.pone.0002932 (2008).
Goulding, M. The Fishes And The Forest: Explorations in Amazonian Natural History. (University of California Press, 1980).
Regarda, V. et al. Geomorphic evidence for recent uplift of the Fitzcarrald Arch (Peru): A response to the Nazca Ridge subduction. 107, 107–117 (2009).
Balon, E. K. Terminology of intervals in fish development. J. Fish. Res. Board Canada 32, 1663–1670 (1975).
Leite, R. G., Cañas, C., Forsberg, B., Barthem, R. & Goulding, M. Larvas dos Grandes Bagres Migradores. 127 (Instituto Nacional de Pesquisas da Amazônia/Asociación para la Conservación de la Cuenca Amazónica, 2007).
Cañas, C. M. & Waylen, P. R. Modelling production of migratory catfish larvae (Pimelodidae) on the basis of regional hydro-climatology features of the Madre de Dios Basin in Southeastern Peru. Hydrol. Processes 26, 996–1007, doi: 10.1002/hyp.8192 (2012).
Acknowledgements
The final synthesis that this paper represents was made possible as part of a Science for Nature and People Partnership (SNAPP - https://www.nceas.ucsb.edu/), and we thank the following partners: from the Wildlife Consrvation Society (WCS), Cristian Samper, John Robinson, Julie Kunen, Mariana Varese, Micaela Varese, Mariana Montoya, Carlos Durigán, Guillermo Estupiñán, Charo Lanao and Natalia Piland; from the Nature Conservancy, Craig Groves and Peter Kareiva; and from the National Center for Ecological Analysis and Synthesis (NCEAS), Frank Davis and Lee Ann French. For general foundational support, we thank Avecita Chicchón (Gordon & Betty Moore Foundation and previously the WCS and the MacArthur Foundation) and Adrian Forsyth and Enrique Ortiz (The Andes-Amazon Fund and previously of the Alton-Jones Foundation and Blue Moon Fund). We also thank the Museu Paraense Emilio Goeldi (MPEG) and the Instituto Nacional de Pesquisa da Amazônia (INPA) for their field and logistical support. Funding was provided by the David and Lucile Packard Foundation (Grant #2013-38757 & #2014-39828), Ward Woods (Grant #309519), the WCS and the Nature Conservancy (TNC). Field data reported for the first time were supported by the Gordon and Betty Moore Foundation (Grant 500) and the John D. and Catherine T. MacArthur Foundation (Grant 84377). The Brazilian National Research Council (CNPq) provided a productivity grant to E.M. Venticinque (Grant 309458/2013-7). Finally, we thank John Lundberg and three anonymous reviewers for carefully reviewing the manuscript and providing many useful suggestions.
Author information
Authors and Affiliations
Contributions
R.B., M.G., and M.L.B.R. elaborated the migratory concept; R.G.L., C.C., and R.B. analyzed the catfish larvae data; M.G., R.B., C.C., and J.C. analyzed the adult catfish distribution; E.V., B.F., P.P., and A.M. elaborated the GIS framework; B.F. analyzed the limnological data; R.B., M.G. and M.L.B.R. wrote the paper and designed the illustrations. All authors discussed the results and revised the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Barthem, R., Goulding, M., Leite, R. et al. Goliath catfish spawning in the far western Amazon confirmed by the distribution of mature adults, drifting larvae and migrating juveniles. Sci Rep 7, 41784 (2017). https://doi.org/10.1038/srep41784
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep41784
This article is cited by
-
‘Disentangling’ the advantages from gillnets in freshwater small-scale fisheries in the Brazilian Amazon
Reviews in Fish Biology and Fisheries (2023)
-
Enhancing our understanding of fish movement ecology through interdisciplinary and cross-boundary research
Reviews in Fish Biology and Fisheries (2022)
-
Semi-natural fishway efficiency for goliath catfish (Brachyplatystoma spp.) in a large dam in the Amazon Basin
Hydrobiologia (2022)
-
Identifying habitat preferences and core areas of Amazon River dolphin activity using spatial ecology analysis
Landscape Ecology (2022)
-
Mobile piscivores and the nature of top-down forcing in Upper Amazonian floodplain lakes
Hydrobiologia (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
