
Abstract
The fossil record of Ginkgo leaf and reproductive organs has been well dated to the Mid-Jurassic (170 Myr). However, the fossil wood record that can safely be assigned to Ginkgoales has not yet been reported from strata predating the late Early Cretaceous (ca. 100 Myr). Here, we report a new fossil wood from the Mid-Late Jurassic transition deposit (153–165 Myr) of northeastern China. The new fossil wood specimen displays several Ginkgo features, including inflated axial parenchyma and intrusive tracheid tips. Because it is only slightly younger than the oldest recorded Ginkgo reproductive organs (the Yima Formation, 170 Myr), this fossil wood very probably represents the oldest bona fide fossil Ginkgo wood and the missing ancestral form of Ginkgo wood evolution.
Similar content being viewed by others


Introduction
The maidenhair tree or Ginkgo is often described as a “living fossil”. It is one of the very few extant plant genera that can be traced back to the Jurassic, at approximately 170 Myr ago. Since Ginkgo apodes Zhou and Zheng1, the “missing link” in Ginkgo evolution, has been reported from North-Eastern China, the genus has a quite complete fossil record lineage with different organs documented, including leaves, pollen, reproductive structures, and long and dwarf shoots. Such a complete record lineage is of great interest for phylogenetical studies and for deciphering the potential role of developmental heterochronies in the evolution of Ginkgoales2.
A tree is made of wood, which forms more than 80% of its biomass. Wood ecophysiology is recognized as a first-order factor in plant evolution3. Unfortunately, the fossil wood record for Ginkgo is very scanty4,5, with no established data predating the late Early Cretaceous, i.e., ca. 100 Myr6,7.
Some woods older than the Cretaceous with supposed and putative relationship links to the Ginkgoales were described as a variety of taxa, such as Baieroxylon Greguss, Plaeoginkgoxylon Feng, Wang et Roessler, Primoginkgoxylon Süß, Proginkgoxylon Zheng et Zhang and Protoginkgoxylon Zheng et Zhang ex Khudaiberdyev5,8,9,10,11,12,13. This profusion of works indicates the strong and lasting interest that the palaeobotany community has in Ginkgo wood evolution5,14,15.
These fossil records predating the Cretaceous are supposedly linked to the Ginkgoales; however, there is no evidence of a direct and certain systematic relationship with Ginkgo. Such relationships are hypothesized for isolated secondary xylem pieces, mostly on the basis of the occurrence of tracheid bunches, with their tips bent alongside the rays (intrusive tracheids), according to Greguss8. However, assigning Palaeozoic, Triassic or Jurassic woods to Ginkgoales represents a challenging task because there is no consensus with regards to Palaeozoic Ginkgophytes systematics or wood anatomy16, and the use of a single xylological feature (intrusive tracheids) is risky17, whereas several other features (e.g., mostly araucarian radial pitting) strongly depart from modern Ginkgo wood anatomy. To sum up, Ginkgo wood anatomy before the Early Cretaceous is still a matter of hypothesis.
Here, we report a new Middle to Late Jurassic fossil wood from northeastern China. While the leaf and reproductive structures of northeastern China-based Ginkgo apodes filled a 100 Myr gap in the fossil record of Ginkgo leaf and sexual organs18, the present fossil wood, at 153–165 Myr ago, is only slightly younger than Ginkgo yimaensis but represents the oldest known Ginkgo wood species19. It thus documents the earliest Ginkgo wood anatomy and establishes what could be called the missing “ancestral form” or the dawn for Ginkgo wood evolution.
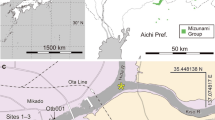
In northeastern China, one of the major fossil wood localities is located in Changgao Town of Beipiao City, western Liaoning, including Lamaying, Shebudaigou, Taizishan, Toudaogou and Renjiagou villages. The fossil wood specimen was collected near Toudaogou village (121°00′–121°09′ E, 41°43′–41°47′ N) in Changgao Town of Beipiao City. The fossil wood locality and geological sections of the Tiaojishan Formation in Beipiao, western Liaoning, northeastern China are described by Wang et al.20.
Stratigraphically, the fossil wood specimen was preserved in the Tiaojishan Formation (previous known as the Lanqi Formation). This formation is widely distributed in western Liaoning Province and the neighbouring northern part of Hebei Province. The Tiaojishan Formation is usually considered to be Middle Jurassic in age, based on palaeontological assemblages21,22. Recent isotopic dating of 40Ar/39Ar in volcanic rock revealed, however, a transition between the late Middle Jurassic and the early Late Jurassic ages (approximately 153 Ma to 165 Ma) for the Tiaojishan Formation23,24.
The Tiaojishan Formation is 2000 m thick and is lithologically composed of intermediate lava and pyroclastic rocks, intercalated with basic volcanic rocks and sedimentary deposits25,26, with plant-bearing beds made of fine-grained sandstones intercalated with shales. These beds contain abundant well-preserved fossil plants, including leaf foliages, seeds and fruits, permineralized rhizomes and fossil wood20. Many anatomically preserved plant specimens were recently reported from the Tiaojishan Formation, including fern rhizomes Ashicaulis27,28,29,30, cycad stem Lioxylon31 and conifers Araucariopitys, Pinoxylon, Sciadopityoxylon and Xenoxylon20,32 (Fig. 1).

Stratigraphic column of the Tiaojishan Formation in Beipiao, Liaoning Province, China (Drawing based on lithological descriptions of Wang et al.22).
1. Plant fossils; 2. Fossil wood; 3. Andesitic conglomerate; 4. Sandstone; 5. Siltstone; 6. Andesitic brecciated lava; 7. Andesitic lava breccias; 8. Andesite; 9. Andesite agglomerate; 10. Conglomeratic tuffaceous sandstone; 11. Shale; 12. Coal seam; 13. Andesitic brecciated tuff; 14. Tuffaceous conglomerate; 15. Quaternary sediments.
Results
Systematic palaeontology
Class: Ginkgopsida
Order: Ginkgoales
Family: incertae sedis
Genus: Ginkgoxylon Saporta
Type species: Ginkgoxylon gruetii Pons et Vozenin-Serra
Ginkgoxylon liaoningense sp. nov. Jiang, Wang, Philippe et Zhang

Ginkgoxylon liaoningense sp. nov. from western Liaoning, China.
(a) Transverse section with a well-marked growth ring. Scale bar = 100 μm. (b) Transverse section with intercellular spaces (red arrows) and locally destroyed walls (black arrows), possibly resulting from a fungal attack. Scale bar = 40 μm. (c) Tangential section; note the tracheid tip contiguous to the ray margin. Scale bar = 100 μm. (d) Tangential section with inflated axial parenchyma associated to a ray (red arrow). Scale bar = 60 μm. (e) Tangential section with tracheid tips associated to inflated axial parenchyma or to ray parenchyma. Scale bar = 100 μm. (f) Tangential section with inflated axial parenchyma associated to a three-cell ray (red arrow). Scale bar = 40 μm.

Ginkgoxylon liaoningense sp. nov. from western Liaoning, China.
(a) A tracheid bunch (red arrow) with tips bent alongside wood rays. Scale bar = 80 μm. (b) A tracheid bunch (red arrow) with storied tips overlapping one another. Scale bar = 40 μm. (c) Tracheid radial pits (red arrow), mostly uniseriate, round and distant. Scale bar = 80 μm. (d) Tracheid radial pits contiguous and biseriate, either alternate (red arrow) or subopposite (black arrow). Scale bar = 40 μm. (e) Pairs of opposite tracheid radial pits within a uniseriate contiguous pit chain (red arrow). Scale bar = 40 μm. (f) Sanio’s rims (black arrows). Scale bar = 40 μm. (g) Ray cells, possible resiniferous content (red arrow) cells, and ray cell transverse walls (black arrows). Scale bar = 40 μm. (h) Cross-fields. Note that the cross-fields are (1) 2–4 cupressoid oculipores, with an oblique aperture (red arrow). Scale bar = 40 μm. (i) Axial parenchyma, with a possible crystalliferous chamber (black arrow). Scale bar = 40 μm. (j) A trabecula (red arrow). Scale bar = 40 μm.
Type specimen: PB22285 and slides. (housed at the palaeobotanical collection of Nanjing Institute of Geology and Palaeontology, CAS).
Type locality: Toudaogou village, Beipiao City, Liaoning Province, China.
Type horizon: Tiaojishan Formation, Middle to Late Jurassic.
Etymology: after the fossil location in Liaoning Province.
Diagnosis: Tracheidoxyl with Ginkgoxylon features, including strongly inflated axial parenchyma chains, idioblasts, intrusive tracheid tips, ordered cupressoid oculipores in cross-fields, opposite pairs of radial pits separated by Sanio’s rims and an irregular aspect of the cross-section; similar to G. chinense Zhang, Zheng et Shang, but with approximately half the radial pits being contiguous and some imbricate biseriate alternate radial pits.
Description
The specimen PB22285 is preserved as a tracheidoxyl, with well-marked growth rings. The transition from early wood to late wood is gradual, with an intermediate type occupying most of the ring (Fig. 2a). The axial cells in cross-section are irregular in size and shape (Fig. 2b), and the tracheids are mostly quadrate to elliptical in the early wood, and more regularly narrowly rectangular in the late wood. Intercellular spaces are often distinct (Fig. 2b, red arrows), and the tracheid wall is thick, even in the early wood, suggesting a compression wood type. The tracheid walls are destroyed in some fungi infected areas (Fig. 2b, black arrows).
In a tangential section (Fig. 2c–f), xylem rays are homogeneous, relatively low, (1) 2–4 (15) cells high and are often associated with inflated axial parenchyma (Fig. 2d,f, red arrows). The tracheid tips are often contiguous to the ray margin or associated to inflated axial parenchyma (Fig. 2c,e).
In the radial section (Fig. 3a–j), some tracheid bunches are present, with storied tips bent alongside wood rays, and the tips sometimes overlap one another (Fig. 3a,b, red arrows). The tracheid radial pits are mostly uniseriate, round and distant (Fig. 3c, red arrow), sometimes contiguous and somewhat flattened, and locally biseriate. The pattern of biseriate pits is variable, from alternate crowded (Fig. 3d, red arrow) to sub-opposite crowded (Fig. 3d, black arrow) or opposite distant (Fig. 3e, red arrow), sometimes with Sanio’s rims (Fig. 3f, black arrows). Ray cells in each crossing field yield 4–6 tracheids in the early wood, and ray cell transverse walls are thin and unpitted (Fig. 3g, black arrows), often resiniferous (Fig. 3g, red arrow); cross-fields with (1) 2–4 cupressoid oculipores, with an oblique aperture (Fig. 3h, red arrow). Oculipores are ordered in columns and lines. The axial parenchyma are abundant and well distributed within the ring, with possible crystalliferous chambers (Fig. 3i, black arrow). Trabeculae are locally preserved (Fig. 3j, red arrow), whereas resin canals are not observed.
Comparison
Except for Protoginkgoxylon, most fossil wood genera with some Ginkgo type features are used for specimens with a strongly araucarian radial pitting. Baieroxylon Greguss has idioblasts but no inflated axial parenchyma chain. Palaeoginkgoxylon Feng, Wang et Roessler, based on a Permian type, has mixed pitting, but with opposite pairs only locally5. This is also the case for Ginkgophytoxylon Vozenin-Serra, Broutin et Toutin-Morin, which was erected based on a type from the Permian of France33. Ginkgomyeloxylon Giraud et Hankel, although diagnosed as having a “Ginkgo type of wood”, is based on a type with strongly araucarian radial pitting15,34. Similarly, Ginkgoxylpropinquus Savidge, despite its diagnosis, is based on a type with Steinkerne-preserved araucarian radial pitting35. This could probably also be the case for Primoginkgoxylon Süß, which has thick-walled idioblasts and uninflated axial parenchyma12.
Protoginkgoxylon was validly recognized by Zheng and Zhang36 based on Permian material (see supplementary note). The type material of Protoginkgoxylon Zheng et Zhang has idioblasts and axial parenchyma, but this is only slightly inflated, never reaching 3–4 tracheids in width as observed in our material. Its radial pitting is described as of the “protopinoid-type”, but the protologue illustrates a pitting much more araucarian than our material. It never shows opposite pit pairs separated by Sanio’s rim. Although they display some Ginkgo-like features, the other species included in Protoginkgoxylon Zheng et Zhang similarly differ from the material studied here by their mixed type of radial pitting being much more araucarian, with usually more than half of the pits being contiguous and/or alternate.
Anatomically, our new fossil wood material combines strongly inflated axial parenchyma chains (in addition to idioblasts), intrusive tracheid tips, cupressoid oculipores in its cross-fields, opposite pairs separated by Sanio’s rims and an irregular aspect of the cross-section. This combination of features is typical for Ginkgo37 and for Ginkgoxylon Saporta11. The diagnosis of Ginkgoxylon as emended by Khudaiberdyev9 does not preclude the inclusion of our material in this genus. Thus, although radial pitting includes a significant share of the araucarian type of radial pitting (approximately 40% to 50%), we assign the present fossil wood material to Ginkgoxylon. Within this genus, our material is most similar to the species of Ginkgoxylon chinense Zhang, Zheng et Shang from the late Early Cretaceous of Liaoning, but it is rather unique because of its partly araucarian radial pitting. Therefore, a new species is proposed for the present fossil wood material from the Jurassic of Beipiao, western Liaoning, China. A comparison of the xyological characters of fossil wood genera linked to Ginkopsida taxa is shown in Supplementary Table S1.
Discussion
Ginkgoales has only one living species, Ginkgo biloba; however, this plant group has an extensive and diverse fossil record. This is evidenced by the numerous fossil leaf taxa, e.g., Baiera Braun, Eretmophyllum Thomas, Ginkgoidium Yokoyama, Ginkgoites Seward, Ginkgoitocladus Krassilov, Glossophyllum Kraeusel, Pseudotorellia Florin, and Sphenobaiera Florin38. Compared with these diverse leaf taxa, we still have a very limited knowledge of the history of Ginkgoales wood diversity. In contrast to most other gymnosperms and angiosperms, the wood of Ginkgo is particularly prone to degradation and hence, is less likely to get fossilized39. Although some fossil wood taxa have been documented based on Paleozoic material, which may be related to Ginkgoales, the fossil wood record of Ginkgoales will probably remain a matter of discussion for a long time.
Modern Ginkgo sometimes has a mixed type of radial pitting, especially in an area with disturbed growth. The amount of mixed radial pitting observed here is consequently not considered to preclude a relationship with Ginkgo. Because there is only one representative of the genus today, the past variability in Ginkgo wood anatomy was probably greater.
Wood with Ginkgo biloba-like anatomy is rare in the fossil record6,40. It is striking that the present fossil wood material in the Tiaojishan Formation, western Liaoning is found in association with several Ginkgo foliage species, including G. huttonii, G. lepida and G. sibirica41,42. In the same strata, however, Ginkgoites (with G. tasiakouensis) and Sphenobaiera (with S. colchica and S. paucipartita) also occur, and thus a link between the studied wood material and a particular foliage species cannot safely be hypothesized. Geologically, our fossil material is only slightly younger than Ginkgo yimaensis. Ginkgo yimaensis is probably the oldest Ginkgo with modern ovulate organs18 at ca. 170 Myr.
There are some other Ginkgoxylon species that have been described and putatively dated as Jurassic. Ginkgoxylon quangnamense Serra was reported based on material found ex-situ in Vietnam43. It might be Early Jurassic in age44; however, this generic identification must be confirmed because the type material was poorly preserved43. The case is similar with Ginkgoxylon dixitii Biradar et Mahabale from Andhra Pradesh of India45. This wood originates from the Kota Formation, which was long thought to be Early Jurassic in age; however, a recent palynological study demonstrated an Early Cretaceous age46. A Ginkgoxylon sp. was described from the Tendaguru strata in Tanzania47. This deposit is only partly Jurassic in age, and the fossil wood material originates from strata that possibly belong to the Aptian (Early Cretaceous)47.
With Ginkgoxylon liaoningense (Middle to Late Jurassic transition), we now have an ancestral form for a species series, continuing with G. chinense Zhang, Zheng et Shang (Aptian, Early Cretaceous), G. gruettii Pons et Vozenin-Serra (Cenomanian, Late Cretaceous), and Ginkgo beckii Scott, Barghoorn et Prakash (Miocene). Anatomical changes in this series are gradual and mostly limited to the radial pitting becoming more and more abietinean.
In conclusion, the fossil material described here from the Tiaojishan Formation in western Liaoning, China, dated as the Middle to Late Jurassic transition in age, is the oldest well-dated occurrence of the genus Ginkgoxylon. Although its anatomy departs slightly from that of the modern Ginkgo, it displays all its characteristic features. It differs only in having a more mixed type of radial pitting, which also occurs, albeit locally, in modern Ginkgo wood. The xylem structures of Ginkgoxylon liaoningense illustrates the basal state of Ginkgo wood anatomy and will contribute to the understanding of Ginkgo evolution.
Methods
The fossil specimen used in this study was preserved as silicified wood. The techniques used for the investigation are the classical thin section method for silicified wood described in Jones and Rowe48. Nomenclatural and taxonomical positions follow those of Philippe10, Bamford and Philippe49 and Philippe and Bamford11. Photographs were taken with ACT-1C DXM1200C software adapted to a Nikon E600 transmitted light microscope. All fossil wood specimens and slides to which this study refers are housed in the Palaeobotany collection of the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, in Nanjing (China), with registration numbers PB22285.
Additional Information
How to cite this article: Jiang, Z. et al. A Jurassic wood providing insights into the earliest step in Ginkgo wood evolution. Sci. Rep. 6, 38191; doi: 10.1038/srep38191 (2016).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Zhou, Z. Y. & Zheng, S. L. The missing link in Ginkgo evolution. Nature 423, 821–822 (2003).
Kvaček, J., Falcon-Lang, H. & Dašková, J. A new Late Cretaceous ginkgoalean reproductive strtucture Nehvizdyella gen. nov. from the Czech Republic and its whole-plant reconstruction. American Journal of Botany 92, 1958–1969 (2005).
Pittermann, J., Stuart, S. A., Dawson, T. E. & Moreau, A. Cenozoic climate changes shaped the evolutionary ecophysiology of the Cupressaceae conifers. PNAS 109, 9647–9652 (2013).
Süß, H. Zwei neue fossile Hölzer der Morphogattung Ginkgoxylon Saporta emend. Süß aus tertiären Schichten der Insel Lesbos, Griechenland, mit einer Übersicht über Fossilien mit ginkgoaler Holzstruktur. Feddes Repertorium 114, 301–319 (2003).
Feng, Z., Wang, J. & Röϐler, R. Palaeoginkgoxylon zhoui, a new ginkgophyte wood from the Guadalupian (Permian) of China and its evolutionary implications. Review of Palaeobotany and Palynology 162, 146–158 (2010).
Pons, D. & Vozenin-Serra, C. Un nouveau bois de Ginkgoales du Cénomanien de l’Anjou, France. Courier Forschungsinstitut Senckenberg 147, 199–213 (1992).
Zhang, W., Zheng, S. L. & Shang, P. A new species of Ginkgoalean wood (Ginkgoxylon chinense Zhang et Zheng sp. nov.) from the Lower Cretaceous of Liaoning, China. Acta Palaeontologica Sinica 39 (sup.), 220–225 (2000).
Greguss, P. Permische fossile Hölzer aus Ungarn. Palaeontogr. Abt. B 109, 131–146 (1961).
Khudaiberdyev, R. Wood of Ginkgo from the Upper Cretaceous of South-West Kyzylkum. Doklady. Akademia Nauk SSSR 145, 422–424 (1962).
Philippe, M. Nomenclature générique des trachéidoxyles mésozoïques à champs araucarioïdes. Taxon 42, 74–80 (1993).
Philippe, M. & Bamford, M. A key to morphogenera used for Mesozoic conifer-like woods. Review of Palaeobotany and Palynology 148, 184–207 (2008).
Süß, H., Rößler, R., Boppré, M. & Fischer, O. W. Drei neue fossile Hölzer der Morphogattung Primoginkgoxylon gen. nov. aus der Trias von Kenia. Feddes Repertorium 120(5/6), 273–292 (2009).
Zheng, S. L. et al. Fossil woods of China (ed. Zheng, S. L. et al.) 1–356 (China Forestry Publishing House, 2008).
Pastro, B. T., Degani, S. I., Guerra, S. M. & Schultz, C.-L. Lenhos de Ginkgophyta em florestas petrificadas no Triassico superior Sul-Rio-Grandense, Brasil. Revista Brasileira Paleontologia 12(2), 139–148 (2009).
Gnaedinger, S. Ginkgoalean woods from the Jurassic of Argentina: taxonomic considerations and palaeogeographical distribution. Geobios 45, 187–198 (2012).
Bauer, K., Kustatscher, E. & Krings, M. The ginkgophytes from the German Kupferschiefer (Permian), with considerations on the taxonomic history and use of Baiera and Sphenobaiera. Bulletin of Geosciences 88(3), 539–556 (2013).
Süß, H. & Kelber, K.-P. Eine neue Art der Morphogattung Baieroxylon Greguss aus dem Keuper von Franken, Deutschland. Feddes Repertorium 122, 257–267 (2011).
Zheng, S. L. & Zhou Z. Y. A new Mesozoic Ginkgo from western Liaoning, China and its evolutionary significance. Review of Palaeobotany and Palynology 131, 91–103 (2004).
Zhou, Z. Y. & Zhang, B. L. A Middle Jurassic Ginkgo with ovule-bearing organs from Henan, China. Palaeontographica (B) 211, 113–133 (1989).
Wang, Y. D., Saiki, K., Zhang, W. & Zheng, S. L. Biodiversity and palaeoclimate of the Middle Jurassic floras from the Tiaojishan Formation in western Liaoning, China. Progress in Natural Sciences 16, 222–230 (2006).
Zhang, W. & Zheng, S. L. Early Mesozoic fossil plants in western Liaoning, northeast China (in Chinese). Mesozoic Stratigraphy and Palaeontology of Western Liaoning (3) (ed. Yu, X. H. et al.) 239–338 (in Chinese) (Geological Publishing House, 1987).
Wang, W. L. et al. Mesozoic stratigraphy and palaeontology of Western Liaoning, vol 1. (in Chinese with English abstract). (Geological Publishing House, 1989).
Chang, S., Zhang, H. C., Renne, P. R. & Fang Y. High-precision 40Ar/39Ar age constraints on the basal Lanqi Formation and its implications for the origin of angiosperm plants. Earth and Planetary Science Letters 279(3–4), 1–10 (2009).
Zhang, H., Wang, M. X. & Liu, X. M. Constraints on the upper boundary age of the Tiaojishan Formation volcanic rocks in West Liaoning-North Hebei by LA-LCP-MS dating. Science Bulletin 58(22), 3574–3584 (2008).
Xu, K. et al. Jurassic in Northern China VII. Northeast Stratigraphic Division. 1–261 (Petroleum Industry Press, 2003).
Jiang B. Y., Yao X. G., Niu Y. Z., Rao X. & Li Q. J. Outline of the Jurassic and Cretaceous systems in western Liaoning. 1–84 (in English, with Chinese translation) (University of Science and Technology of China Press, 2010).
Tian, N., Wang, Y. D., Zhang, W., Jiang, Z. K. & Dilcher, D. L. Ashicaulis beipiaoensis sp. nov., a new Osmundaceous fern species from the Middle Jurassic of Liaoning Province, Northeastern China. International Journal of Plant Sciences 174(3), 328–339 (2013).
Tian, N., Wang, Y. D., Zhang, W. & Jiang, Z. K. A new structurally preserved fern rhizome of Osmundaceae (Filicales) Ashicaulis wangii sp. nov. from the Jurassic of Western Liaoning and its significances for palaeobiogeography and evolution. Science China, Earth Sciences 57(4), 671–681 (2014).
Tian, N. et al. A specialized new species of Ashicaulis (Osmundaceae, Filicales) from the Jurassic of Liaoning, NE China. Journal of Plant Research 127, 209–219 (2014).
Tian, N., Wang, Y. D., Dong, M., Li, L. Q. & Jiang, Z. K. A systematic overview of fossil osmundalean ferns in China: Diversity variation, distribution pattern, and evolutionary implications. Palaeoworld 25(2), 149–169 (2016).
Zhang, W., Wang, Y. D., Saiki, K., Li, N. & Zheng, S. L. A structurally preserved Cycad-like stem, Lioxylon gen. et sp. nov., from the Jurassic in western Liaoning, China. Progress in Natural Science 26 (Special issue), 236–248 (2006).
Jiang, Z. K., Wang, Y. D., Zheng, S. L., Zhang, W. & Tian, N. Occurrence of Sciadopitys-like fossil wood (conifer) in the Jurassic of western Liaoning and its evolutionary implications. Chinese Science Bulletin 57(6), 569–572 (2012).
Vozenin-Serra, C., Broutin, J. & Toutin-Morin, N. Bois permiens du sud-ouest de l’Espagne et du sud-est de la France. Implications pour la taxonomie des gymnospermes paléozoïques et la phylogénie des Ginkgophytes. Palaeontographica Abteilung B 221, 1–26 (1991).
Giraud, B. & Haenkel, O. Nouveaux bois fossiles des dépôts du Karoo du Bassin de Luwegu (Tanzanie méridionale). Annales de Paléontologie 72(1), 1–27 (1986).
Savidge, R. A. Xylotomic evidence for two new conifers and a Ginkgo within the Late Triassic Chinle Formation of Petrified Forest National Park, Arizona, USA. In Parker, W. G., Ash, S. R., Irmis, R. B. (Eds.), A Century of Research at Petrified Forest National Park: Geology and Paleontology. Museum of Northern Arizona special publications 62, 147–149 (2006).
Zheng, S. L. & Zhang, W. Late Paleozoic ginkgoalean woods from northern China. Acta Palaeontologica Sinica 39, 119–126 Supl (2000).
Süß, H. Zur Problematik des Nachweises fossiler Ginkgo-Holzreste. Zeitschrifft für geologischen Wissenchaften 16, 335–336 (1988).
Zhou, Z. Y. Mesozoic ginkgoalean megafossils: a systematic review A. Ginkgo biloba-A global treasure from biology to medicine (ed. Hori, T. et al.) 183–206 (Springer Verlag, 1997).
Scott, R. A., Barghoorn, E. S. & Prakash, U., Wood of Ginkgo in the Tertiary of Western North America. American Journal of Botany 49(10), 1095–1101 (1962).
Taylor, T. N., Taylor, E. L. & Krings, M. Paleobotany. The biology and evolution of fossil plants. 1–1230 (Academic Press, Elsevier, 2009).
Van Konijnenburg-van Cittert, J. H. A. In situ gymnosperm fructifications from the Jurassic flora of Yorkshire. Acta Bot. Neerl. 20, 1–96 (1971).
Heer, O. Beiträge zur Jura-Flora Ostsibiriens and Amurlandes. Mémoires de l’Académie Impériale des Sciences de St.-Pétersbourg 25(6), 1–122 (1876).
Serra, C. Sur un nouveau bois de Ginkgoales récolté dans le Ho-Gia (Province du Quang-nam). Archives Géologiques du. Vietnam 10, 93–103 (1967).
Philippe, M. Radiation précoce des conifères Taxodiaceae et bois affines du Jurassique de France. Lethaia 27, 67–75 (1994).
Philippe, M. et al. Biogeography of Gondwanan terrestrial biota during the Jurassic - Early Cretaceous as seen from fossil wood evidence. Review of Palaeobotany and Palynology 129, 141–173 (2004).
Rajanikanth, A. & Chinnappa, C. Continental Jurassic of peninsular India – a floristic and stratigraphic riddle. 9th International Congress on the Jurassic System, 6th to 9th January 2014 in Jaipur, India (2014).
Kahlert, E., Schultka, S. & Süß, H. Die mesophytische Flora der Saurierlagerstätte am Tendaguru (Tansania). Erste Ergebnisse. Mitteilungen Museum Naturkunde Berlin, Geowissenschaftliche Reihe 2, 185–199 (1999).
Jones, T. P. & Rowe, N. P. Fossil plants and spores: modern techniques. 1–396 (Geological Society, 1999).
Bamford, M. & Philippe, M. Gondwanan Jurassic – Early Cretaceous homoxylous woods: a nomenclatural revision of the genera with taxonomical notes. Review of Palaeobotany and Palynology 113, 287–289 (2001).
Acknowledgements
This work was financially supported by the Strategic Priority Research Program (B) of the Chinese Academy of Sciences (XDB18030502), the State Key Program of Research and Development of Ministry of Science and Technology, China (2016YFC0600406); the National Natural Sciences Foundation of China (NSFC 41402004, 41272010, 41572014, and 41302004), the Team Program of Scientific Innovation and Interdisciplinary Cooperation of CAS. This is a contribution to IGCP 632.
Author information
Authors and Affiliations
Contributions
Z.J., Y.W., and M.P. designed and performed research; Z.J., Y.W., M.P. and W.Z. wrote the manuscript. Z.J., N.T. and S.Z. prepared the figures and table; All authors commented on and approved this version of the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Jiang, Z., Wang, Y., Philippe, M. et al. A Jurassic wood providing insights into the earliest step in Ginkgo wood evolution. Sci Rep 6, 38191 (2016). https://doi.org/10.1038/srep38191
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep38191
This article is cited by
-
Occurrence of Brachyoxylon Hollick et Jeffrey from the Lower Cretaceous of Zhejiang Province, southeastern China
Journal of Palaeogeography (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
