
Abstract
The principal aim of this study was to investigate the in vitro co-infection of Caco-2 cells with Candida albicans and enterohemorrhage Escherichia coli (EHEC). The ability of both species to colonize or invade the Caco-2 cells was evaluated by indirect immunofluorescence and inverted microscopy. The damage to Caco-2 cells was evaluated by measuring lactate dehydrogenase (LDH) activity. C. albicans virulence gene expression (HWP1, ALS3, PLB1, SAP4, and EFG1) was evaluated by quantitative real-time polymerase chain reaction (qRT-PCR). Compared to single infections with enterohemorrhage Escherichia coli or C. albicans, a co-infection colonized or invaded Caco-2 cells more quickly, and C. albicans tended to accumulate more easily, accompanied by the upregulation of related genes. In addition, the LDH activity in the co-infected group was higher than in cells infected with C. albicans or with enterohemorrhage Escherichia coli, accompanied by the upregulation of toxicity-related genes. Using Caco-2 cells as an infection model, this study demonstrated that co-infecting in vitro enterocytes with C. albicans and enterohemorrhage Escherichia coli enhanced the invasiveness and tissue damaging effects of C. albicans.
Similar content being viewed by others
Introduction
As opportunistic fungal pathogens, Candida species are normally harmless commensals in the gastrointestinal tract, genitourinary tract or oropharyngeal tract of most healthy individuals1. However, in specific conditions, such as long-term antibiotic treatment, immunocompromisation or critical illness, Candida species can also turn into pathogens and cause superficial infections, such as oral or vaginal candidiasis, deep-seated infections, and systemic infections2,3.
The main pathogenic Candida species in the human body is C. albicans4,5, notably the C. albicans in gastrointestinal tract. C. albicans can not only cause intestinal candidosis, but can also destroy the intestinal epithelial barrier and result in the occurrence of candidemia6,7. However, most common clinical situations involve a co-infection by various microorganisms, rather than a single infection by one microorganism. In recent years, there have been reports on the effects of co-infection leading to invasive candidosis8,9,10,11,12. Several studies have reported the enhanced adherence and invasiveness of C. albicans on epithelium cells, as well as the tissue damage caused by C. albicans during co-infection8,11,12,13,14,15. Most of these studies focused on co-infection by various Candida species; there are few studies focused on co-infection by Candida species and a non-Candida species13,16,17.
The aim of this study was to characterize a C. albicans and enterohemorrhage Escherichia coli co-infection on enterocytes in vitro using Caco-2 cells. Indirect immunofluorescence and quantitative real-time polymerase chain reaction (qRT-PCR) was used to evaluate results. The expression of virulence genes (HWP1, ALS3, PB1, SAP4 and EFG1) was also analyzed.
Results
Colonization or Invasion of Caco-2 cells by C. albicans
The positive rates of detection in the single infection group and the co-infection group across all time points are summarized in Table 1. Results indicated that the positive rates in the co-infection group were significantly higher than those in the single infection group within the first hour after infection. At later time points, no statistically significant differences were detected between the groups.
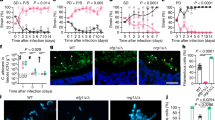
Under inverted microscopy, in the single infection group (group 1), C. albicans were detected at the yeast phase, pseudohyphal phase and hyphal phase from the time point of 1 hour onward (Fig. 1a), 2 hours onward (Fig. 1b) and 3 hours onward (Fig. 1c) after infection, respectively. In the co-infection group (group 3), C. albicans were detected at the yeast phase, pseudohyphal phase and hyphal phase from the time point of 30 minutes onward (Fig. 1d), 2 hours onward (Fig. 1e) and 3 hours onward (Fig. 1f) after infection, respectively. C. albicans tended to cluster more often in the co-infection group than in the single infection group. In addition, enterohemorrhage Escherichia coli and C. albicans were present in the same location in the co-infection group (Fig. 1g).

The morphogenic states of C.
albicans during infection in the single infection group (a–c) and the co-infection group (e–f) at different observation time points (A 1 hour, B 2 hours, C 3 hours, D 0.5 hours, E 2 hours, F 3 hours). Green fluorescent dots (blue arrows) indicate enterohemorrhage Escherichia coli and red arrows indicate C. albicans in the co-infection group (g). Magnification: 10 × 20.
LDH activity as an indicator of tissue damage
In order to determine the extent of Caco-2 cell damage induced by Candida, the levels of LDH activity were measured every 2 hours for 24 hours after infection. It is important to highlight that half of the number of Candida cells and enterohemorrhage Escherichia coli cells were present in the starting inoculum of the co-infection group. Repeated measures analysis of variance was used to test if the measurements were equal between each time point (the measurement at the first time point was taken as the control). The results suggested that LDH values in group 1, 2, and 3 increased with time. LDH values in group 1, 2 and 3 significantly increased at 20 hours, 12 hours and 10 hours, respectively (Fig. 2). When comparing the total trends among the three groups, it was found that LDH values in group 2 were greater than those in group 1, taking all measurements as a whole (P < 0.001), group 3 greater than in group 1 (P = 0.0004), and group 3 greater than in group 2 (P = 0.0028). These results indicate that the tissue damage in the co-infection group was greater than the damage of a single infection.

LDH activity at different time points (black asterisk: 20 hours; red asterisk: 12 hours; blue asterisk: 10 hours).
*Significantly higher than the corresponding preoperative level (P < 0.05).
Gene expression
qRT-PCR revealed a wide range of expression of toxicity-related genes. The results were expressed as the mean percentage expression, relative to the expression of ACT1 during Caco-2 cell infection. ACT1 gene expression levels were constant in all assays.
HWP1 and ALS3 expression by C. albicans was upregulated during co-infection with enterohemorrhage Escherichia coli 6 hours post-infection (P = 0.0005, P = 0.0491) (Fig. 3). Expression of both PLB1 and SAP4 genes was detected 6 and 24 hours after infection in both groups. PLB1 gene expression was upregulated in the co-infection group at 24 hours after infection (P = 0.0193) (Fig. 3), but SAP4 gene expression was upregulated in the co-infection group at both 6 and 24 hours after infection (P = 0.0019, P = 0.0062) (Fig. 3). EFG1 gene expression demonstrated similar results to SAP4 expression: an upregulation in the co-infection group at both 6 and 24 hours post-infection (P = 0.0473, P = 0.0227) (Fig. 3).

mRNA relative expression at 6 hours (a) and 24 hours (b) after infection. *Significantly different between groups (P < 0.05).
Discussion
The intestinal mucosal barrier is highly important in host defense, as it is the first cell layer that prevents an invasion of pathogens, such as Candida species and enterohemorrhage Escherichia coli18. In recent years, a number of studies have demonstrated that the gastrointestinal tract is the main reservoir for C. albicans and a source of gastrointestinal candidiasis and candidemia6,7. It was found in our study that enterohemorrhage Escherichia coli promotes the colonization and invasiveness and tissue damaging effects of C. albicans to enterocytes in vitro.
The invasion of epithelial cells by C. albicans is a key element in the physiopathology of candidiasis. It is generally accepted that yeast are the noninvasive, commensal morphology19,20. However, in our study, we discovered that yeast-phase C. albicans could colonize or invade Caco-2 cells in the early stages of both single and co-infections. However, the colonization or invasion of Caco-2 cells by C. albicans occurred earlier when combined with enterohemorrhage Escherichia coli than it did during a single infection. Two other observations were that enterohemorrhage Escherichia coli and C. albicans were present in the same location in the co-infection group and that the co-infection group tended to cluster more often than the single infection group. This suggests that C. albicans colonized or invaded enterocytes more efficiently in the co-infection group. It is interesting to note that there were no differences between the single infection and co-infection groups when it came to the presence of the pseudohyphal and hyphal phases. Although the underling mechanism remains unclear, one possible explanation is that enterohemorrhage Escherichia coli promotes the superficial infection of Caco-2 cells with C. albicans by inducing damage to the epithelium. This would facilitate the access of C. albicans to the epithelial layers.
The invasion of host cells by C. albicans is associated with expression of the hyphal-associated genes. ALS3 is a hyphal-specific gene expressed by C. albicans hyphae and pseudohyphae, but not yeast21,22. Both ALS321,22,23 and HWP123,24 were directly associated with hyphae formation and invasiveness in C. albicans. In this experiment, qRT-PCR showed that during co-infection, C. albicans strains exhibited an upregulation of HWP1 and ALS3 compared to a single infection at 6 hours post-infection. Based on the qRT-PCR results and the functions of ALS3 and HWP1, it is tempting to speculate that invasiveness of C. albicans in the co-infection group is enhanced by the presence of C. albicans hyphae following HWP1 and ALS3 upregulation.
LDH activity was used to evaluate the degree of tissue damage in the single infection and co-infection groups. Of the single species infection groups, enterohemorrhage Escherichia coli caused more damage than C. albicans. Compared to group 1, in group 2, tissue damage was more severe and occurred earlier. This result was expected, given the fact that enterohemorrhage Escherichia coli is deemed more pathogenic. In the co-infection group, the degree of damage was more severe than both groups 1 and 2. These results suggest that tissue damage in the co-infection group was greater than the damage of a single infection. According to these findings, it is obvious that potential synergism may exist between microorganisms in a co-infection that enhances damage caused to epithelial cells.
In order to better elucidate pathogenic differences, PLB1 and SAP4 genes expressions were measured. PLB1 is considered an important virulent factor in intestinal C. albicans bacteremia25. Expression of this enzyme can lead to the disruption of membrane structure and function26. The SAP gene family was also confirmed to play a crucial rule in the virulence of C. albicans27. The current study showed that the expression of PLB1 and SAP4 genes was always detected at 24 hours after infection, confirming a potential role of these genes in enterocyte cell damage. In addition, PLB1 and SAP4 were expressed at a higher level in the co-infection group than in the single infection groups, indicating that enterohemorrhage Escherichia coli promotes the tissue damaging effects of C. albicans, with the presence of extracellular hydrolytic enzymes following PLB1 and SAP4 upregulation.
It is well established that EFG1 and components of its upstream regulatory pathway are essential for all stages of C. albicans-epithelial interaction23. In the present study, the EFG expression of C. albicans by qRT-PCR was measured both in the single infection and co-infection groups. The results showed that EFG expressions at both 6 and 24 hours after infection were greater in the co-infection group than in the single infection group, indicating that enterohemorrhage Escherichia coli promoted the ability of C. albicans to invade and cause tissue damage to Caco-2 cells.
It should be noted that the inverted microscope and the immunofluorescence methodology used in the present study cannot discriminate between C. albicans that are adherent to Caco-2 cells versus ones that have invaded Caco-2 cells.
In summary, the present study confirms the effectiveness of Caco-2 cells as an in vitro model to study the attributes of Candida virulence. Usage of indirect immunofluorescence, LDH activity evaluation, and the developed methodology of RT-PCR for Candida cell quantification in tissue, has conclusively shown that co-infecting in vitro enterocytes with C. albicans and enterohemorrhage Escherichia coli enhanced the invasiveness and tissue damaging effects of C. albicans.
Material and Methods
Microorganisms
Enterohemorrhage Escherichia coli O157:H7 and C. albicans ATCC 10231 (obtained from the American Type Culture Collection) were used for all assays in this study. The identity of the C. albicans strains were confirmed by PCR-based sequencing using specific primers ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′)28.
C. albicans strains were routinely cultured in liquid YPD (1% yeast extract, 2% bacto-peptone, 2% D-glucose) medium in a shaking incubator at 120 rev/min for 48 hours at 37 °C, and maintained in solid YPD (2% agar was added to liquid YPD). Enterohemorrhage Escherichia coli strains were cultured in LB (0.5% yeast extract, 1% tryptone, 1% NaCL) broth at 37 °C for 10 hours.
Both microorganisms were measured with the viable plate count method. After this, the cellular densities of C. albicans and E. coli were adjusted to 2 × 106 cells/ml before the experiment. C. albicans cells were grown to log phase for another 2 hours at 37 °C prior to usage.
Culture of intestinal epithelial cells
The colon adenocarcinoma-derived cell line Caco-2 (obtained from the Insitute of Biochemistry and Cell Biology, Shanghai, China) was used as an in vitro model for intestinal cells in this study. Caco-2 cells are capable of forming monolayer cells that are similar, in many ways, to normal human enterocytes. These cells were routinely cultured in Dulbecco’s modified eagle medium (DMEM), supplemented with 10% fetal calf serum (FCS), without antibiotics or antifungal agents. The cells were maintained in a humidified incubator at 37 °C in 5% CO2 environment in saturated humidity. For every experiment, 1 × 105 Caco-2 cells were seeded onto each well of a 12-well plate and cultured up to a high cell density before the experiment.
Groups and infection
Four groups were assigned to study in vitro single infection and co-infection of Caco-2 cells by C. albicans and enterohemorrhage Escherichia coli. In group 1, Caco-2 cells were infected with 1 ml 2 × 106 C. albicans cells. In group 2, Caco-2 cells were infected with 1 ml 2 × 106 enterohemorrhage Escherichia coli cells. These two groups, 1 and 2, were single species infections. In group 3, Caco-2 cells were infected with both 0.5 ml 2 × 106 C. albicans cells and 0.5 ml 2 × 106 enterohemorrhage Escherichia coli cells. Group 4 was set as a negative control and neither infected with C. albicans nor enterohemorrhage Escherichia coli. All groups were incubated at 37 °C in a 5% CO2 environment in saturated humidity.
The surface colonization or invasion of Caco-2 cells by C. albicans
The in vitro adherence assay was performed according to a previously described protocol29. The surface colonization or invasion of Caco-2 cells by C. albicans in single and co-infections was determined using inverted microscopy and indirect immunofluorescence. Briefly, the Caco-2 cells were infected with C. albicans and enterohemorrhage Escherichia coli. Every 30 minutes after infection, the medium above the cells was aspirated and the monolayers were rinsed three times with phosphate buffer solution (PBS) to remove fungal cells not associated with epithelial cells, followed by fixation in 80% alcohol for 40 minutes at 4 °C. All fungal cells remaining that had colonized or invaded the surface of the Caco-2 cells were determined directly by inverted microscopy. Once the presence of C. albicans was observed, the colonization or invasion of Caco-2 cells by C. albicans was considered positive. The experiments were repeated twelve times at each time point. The positive rate at each time point was calculated as the number of positive detections divided by 12 experiments. Fisher’s exact test was used to examine the differences in the positive rate between the single infection and co-infection groups. Enterohemorrhage Escherichia coli was stained by Anti-E. coli O157 antibody (FITC) (abcam). After rinsing with PBS three times, each well was fully observed under indirect immunofluorescence in sequences to ensure all fields were observed.
Damage assay: Lactase dehydrogenase (LDH) activity
The extracellular activity of LDH released from infected enterocytes into the medium was monitored as a measure of tissue damage. A Cytotoxicity Detection Kit (LDH) (Roche Diagnostics, Indianapolis, IN) and Modular P800 (Roche) were used for the damage assay. Arguments of Modular P800 were set as follows: Assay-Rate A, Wavelength-492/690 nm, time-10 mins, point- 22~32.
The extracellular LDH activity of all groups was continuously measured at 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 and 24 hours after infection. The degree of tissue damage was defined as the LDH activity of experimental groups (groups 1, 2, and 3) subtracting the LDH activity of the control group, which was incubated with growth medium alone (group 4). The experiments were performed three times.
Analysis of fungal virulence gene expression
RNA extraction
For RNA extraction, samples were collected in a 1.5 ml microcentrifuge tube and frozen by dipping in liquid nitrogen, followed by adding 500 μl Buffer RB/ ß-mercaptoethanol. Water-saturated phenol, 2 M NaAc (pH 4.0), and chloroform were added to the mixture in order to lyse cells. Fungal RNA extraction kit (E.Z.N.A. ® Fungal RNA Kit, Omega) was then used to complete total RNA extraction from the tissue, according to the manufacturer’s recommendations. The procedure was performed twice.
Synthesis of complementary DNA (cDNA)
For synthesis of cDNA, PrimeScript®RT Master Mix (Takara) was used to synthesize cDNA, following the manufacturer’s recommendations with a modification. Briefly, 8 μl RNA was incubated with 2 μl 5X PrimeScript RT Master Mix at 37 °C for 15 minutes. Then, the mixture was placed at 80 °C for 10 seconds to stop the reaction of synthetase.
Primer design
The following pairs of primers were used in this assay: ACT130, EFG130, HWP131, ALS332, SAP433, and PLB113 (Table 2). The ACT1 had been previously used as a housekeeping gene and was also used in this study30. To verify the specificity of each primer pair for its corresponding target gene, PCR using the various primer pairs was applied to genomic DNA extracted from each of the Candida strains. The mRNA expression levels of these selected genes were measured to evaluate the impacts of co-infection.
Quantitative real-time PCR (qRT-PCR) assay
qRT-PCR was performed using SYBR Premix Ex Taq (Takara). A 20 μl mixture solution was composed of 10 μl SYBR Premix Ex Taq, 2 μl primers, 1 μl cDNA and 7 μl deionized water. qRT-PCR was performed with an initial denaturation step at 94 °C for 30 seconds, followed by 40 cycles of denaturation at 95 °C for 5 seconds, and primer annealing at 60 °C for 34 seconds. A melting curve was generated at 95 °C for 15 seconds, 60 °C for 1 minute and 95 °C for 15 seconds at the end of each PCR cycle to verify that a specific product was amplified. Control samples were included on each plate to ensure that multiple plates could be compared. All samples were run in triplicate. The Ct value of each sample was determined, and the relative gene expression levels were calculated using the ΔΔCt method, which was normalized to the housekeeping genes described above, as a control.
Statistical analysis
Data were analyzed using the SAS 9.4 statistical software (SAS Institure, Inc., Cary, NC). Continuous variables were expressed as the mean ± standard deviation while categorical variables were expressed as frequency and percentage. The independent Student’s t test was used to compare mRNA relative expression. Fisher’s exact test was performed to compare the positive detection rate of C. albicans between group 1 and group 3. Repeated measures analysis of variance was used to compare the LDH values within and between groups by taking different times for measurement as the repeated factor. Statistical significance level was set at 0.05.
Additional Information
How to cite this article: Yang, W. et al. Enterohemorrhagic Escherichia coli promotes the invasion and tissue damage of enterocytes infected with Candida albicans in vitro. Sci. Rep. 6, 37485; doi: 10.1038/srep37485 (2016).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Lim, C. S., Rosli, R., Seow, H. F. & Chong, P. P. Candida and invasive candidiasis: back to basics. Eur J Clin Microbiol Infect Dis. 31, 21–31 (2012).
Pfaller, M. A. & Diekema, D. J. Epidemiology of invasive mycoses in North America. Crit Rev Microbiol. 36, 1–53 (2010).
Yang, W., Yan, L., Wu, C., Zhao, X. & Tang, J. Fungal invasion of epithelial cells. Microbiol Res. 169, 803–810 (2014).
Pfaller, M. A. & Diekema, D. J. Epidemiology of invasive candidiasis: a persistent public health problem. Clin Microbiol Rev. 20, 133–163 (2007).
Bassetti, M. et al. Epidemiology, species distribution, antifungal susceptibility and outcome of nosocomial candidemia in a tertiary care hospital in Italy. PLoS One. 6, e24198 (2011).
Nucci, M. & Anaissie, E. Revisiting the source of candidemia: skin or gut? Clin Infect Dis. 33, 1959–1967 (2001).
Miranda, L. N. et al. Candida colonisation as a source for candidaemia. J Hosp Infect. 72, 9–16 (2009).
Diaz, P. I. et al. Synergistic interaction between Candida albicans and commensal oral streptococci in a novel in vitro mucosal model. Infect Immun. 80, 620–632 (2012).
Morales, D. K. & Hogan, D. A. Candida albicans interactions with bacteria in the context of human health and disease. PLoS Pathog. 6, e1000886 (2010).
Kali, A. et al. Prevalence of Candida co-infection in patients with pulmonary tuberculosis. Australas Med J. 6, 387–391 (2013).
Shirtliff, M. E., Peters, B. M. & Jabra-Rizk, M. A. Cross-kingdom interactions: Candida albicans and bacteria. FEMS Microbiol Lett. 299, 1–8 (2009).
Bamford, C. V. et al. Streptococcus gordonii modulates Candida albicans biofilm formation through intergeneric communication. Infect Immun. 77, 3696–3704 (2009).
Alves, C. T. et al. Candida albicans promotes invasion and colonisation of Candida glabrata in a reconstituted human vaginal epithelium. J Infect. 69, 396–407 (2014).
Silva, S. et al. Candida glabrata and Candida albicans co-infection of an in vitro oral epithelium. J Oral Pathol Med. 40, 421–427 (2011).
Cavalcanti, Y. W. et al. Virulence and pathogenicity of Candida albicans is enhanced in biofilms containing oral bacteria. Biofouling. 31, 27–38 (2015).
Tati, S. et al. Candida glabrata Binding to Candida albicans Hyphae Enables Its Development in Oropharyngeal Candidiasis. PLoS Pathog. 12, e1005522 (2016).
Barros, P. P., Ribeiro, F. C., Rossoni, R. D., Junqueira, J. C. & Jorge, A. O. Influence of Candida krusei and Candida glabrata on Candida albicans gene expression in in vitro biofilms. Arch Oral Biol. 64, 92–101 (2016).
Yan, L., Yang, C. & Tang, J. Disruption of the intestinal mucosal barrier in Candida albicans infections. Microbiol Res. 168, 389–395 (2013).
Berman, J. & Sudbery, P. E. Candida Albicans: a molecular revolution built on lessons from budding yeast. Nat Rev Genet. 3, 918–930 (2002).
Saville, S. P., Lazzell, A. L., Monteagudo, C. & Lopez-Ribot, J. L. Engineered control of cell morphology in vivo reveals distinct roles for yeast and filamentous forms of Candida albicans during infection. Eukaryot Cell. 2, 1053–1060 (2003).
Hoyer, L. L., Payne, T. L., Bell, M., Myers, A. M. & Scherer, S. Candida albicans ALS3 and insights into the nature of the ALS gene family. Curr Genet. 33, 451–459 (1998).
Argimon, S. et al. Developmental regulation of an adhesin gene during cellular morphogenesis in the fungal pathogen Candida albicans. Eukaryot Cell. 6, 682–692 (2007).
Wachtler, B., Wilson, D., Haedicke, K., Dalle, F. & Hube, B. From attachment to damage: defined genes of Candida albicans mediate adhesion, invasion and damage during interaction with oral epithelial cells. PLoS One. 6, e17046 (2011).
Sundstrom, P., Balish, E. & Allen, C. M. Essential role of the Candida albicans transglutaminase substrate, hyphal wall protein 1, in lethal oroesophageal candidiasis in immunodeficient mice. J Infect Dis. 185, 521–530 (2002).
Mukherjee, P. K. et al. Reintroduction of the PLB1 gene into Candida albicans restores virulence in vivo. Microbiology. 147, 2585–2597 (2001).
Niewerth, M. & Korting, H. C. Phospholipases of Candida albicans. Mycoses. 44, 361–367 (2001).
Moran, G. P., Coleman, D. C. & Sullivan, D. J. Candida albicans versus Candida dubliniensis: Why Is C. albicans More Pathogenic? Int J Microbiol. 2012, 205921 (2012).
Williams, D. W., Wilson, M. J., Lewis, M. A. & Potts, A. J. Identification of Candida species by PCR and restriction fragment length polymorphism analysis of intergenic spacer regions of ribosomal DNA. J Clin Microbiol. 33, 2476–2479 (1995).
Dalle, F. et al. Cellular interactions of Candida albicans with human oral epithelial cells and enterocytes. Cell Microbiol. 12, 248–271 (2010).
Toyoda, M., Cho, T., Kaminishi, H., Sudoh, M. & Chibana, H. Transcriptional profiling of the early stages of germination in Candida albicans by real-time RT-PCR. FEMS Yeast Res. 5, 287–296 (2004).
Sellam, A. et al. Role of transcription factor CaNdt80p in cell separation, hyphal growth, and virulence in Candida albicans. Eukaryot Cell. 9, 634–644 (2010).
Cheng, G. et al. Comparison between Candida albicans agglutinin-like sequence gene expression patterns in human clinical specimens and models of vaginal candidiasis. Infect Immun. 73, 1656–1663 (2005).
Naglik, J. R. et al. Quantitative expression of the Candida albicans secreted aspartyl proteinase gene family in human oral and vaginal candidiasis. Microbiology. 154, 3266–3280 (2008).
Acknowledgements
Supported by Shanghai Municipal Commission of Health and Family Planning Research Project (Grant No. 201540136) and Minhang Municipal Commission of Health and Family Planning Research Project (Grant No. 2013WM12).
Author information
Authors and Affiliations
Contributions
Conception and design (J.T., W.Y. and Y.Z.); data collection (W.Y. and C.W.); analysis and interpretation of data (J.T., W.Y., Y.Z. and C.W.); writing the manuscript (W.Y. and C.W.); critical revision of the manuscript (J.T. and Y.Z.); administrative, technical, or material support (J.T.); supervision (J.T.).
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Yang, W., Zhou, Y., Wu, C. et al. Enterohemorrhagic Escherichia coli promotes the invasion and tissue damage of enterocytes infected with Candida albicans in vitro. Sci Rep 6, 37485 (2016). https://doi.org/10.1038/srep37485
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep37485
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
