
Abstract
A DREB-type transcription factor gene GmDREBL has been characterized for its functions in oil accumulation in seeds. The gene is specifically expressed in soybean seeds. The GmDREBL is localized in nucleus and has transcriptional activation ability. Overexpression of GmDREBL increased the fatty acid content in the seeds of transgenic Arabidopsis plants. GmDREBL can bind to the promoter region of WRI1 to activate its expression. Several other genes in the fatty acid biosynthesis pathway were also enhanced in the GmDREBL-transgenic plants. The GmDREBL can be up-regulated by GmABI3 and GmABI5. Additionally, overexpression of GmDREBL significantly promoted seed size in transgenic plants compared to that of WT plants. Expression of the DREBL is at higher level on the average in cultivated soybeans than that in wild soybeans. The promoter of the DREBL may have been subjected to selection during soybean domestication. Our results demonstrate that GmDREBL participates in the regulation of fatty acid accumulation by controlling the expression of WRI1 and its downstream genes and manipulation of the gene may increase the oil contents in soybean plants. Our study provides novel insights into the function of DREB-type transcription factors in oil accumulation in addition to their roles in stress response.
Similar content being viewed by others

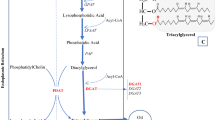
Introduction
Soybean is an important economic crop and provides oil and proteins for human and animals. Increasing the FA (fatty acid) contents and improving the oil quality are closely related to our daily life. So far, numerous efforts have been made to meet the needs of human food and industry production by changing the fatty acid content in seeds1. However, the extracted fatty acids from the existing oil plants are far from enough and hence traditional breeding methods and transgenic approaches manipulating fatty acid biosynthesis pathway are used to increase oil content in soybean. In plants, the pathways for lipid biosynthesis and oil accumulation had been studied and the genes related to fatty acid biosynthesis have been characterized. There are several key genes in the process of fatty acid biosynthesis. One is ACCase encoding acetyl CoA carboxylase in the first key step of fatty acid biosynthesis and malonyl-CoA is produced2. The second one is KASIII, which encodes 3-ketoacyl-ACP synthase III to catalyze the formation of a 4-carbon product3,4. The carbon number of fatty acid is increased by two in acyl chain and elongation of the acyl chain from six to 16 carbon molecules is catalyzed by an enzyme named KAS15. Without KAS1, FA contents would be sharply reduced and plant growth and development would be strongly affected6. The genes related to FA biosynthesis such as Pl-PKβ1 (pyruvate kinase), PDHE1α (pyruvate dehydrogenase E1 alpha subunit), BCCP2 (acetyl-CoA carboxylase), ACP1 (acyl carrier protein) and KAS1 have similar expression pattern with WRI1 (WRINKLED1) and the FA biosynthesis-related genes were up-regulated in the WRI1-overexpressing plants7. WRI1 is an AP2-type transcription factor (TF) with two AP2 DNA-binding domains8 and it appears to be a master regulator of FAS (fatty acid synthesis) genes in expression level. There is a specific sequence motif AW-box in the promoter regions of the FAS genes and WRI1 binds to this motif in Arabidopsis9. Overexpression of WRI1 enhanced the oil content in transgenic Arabidopsis6,8,9,10,11 and maize12,13. In Castor bean, there are WRI1 binding consensus sites in the promoter region of RcBCCP2 and RcKAS1 and RcWRI1 possibly binds to these sites to play a pivotal role in fatty acid biosynthesis14. Overexpression of a single transcription factor gene WRI can increase the seed oil contents while manipulating a single fatty acid biosynthesis gene had only very limited effect on the oil content15,16.
Transcription factors can regulate expression of genes involved in a wide range of plant processes and have a cascade amplification effect8,17. Therefore, transcription factors are the promising targets to improve oil contents in plants. Several candidate transcription factors involved in fatty acid biosynthesis and accumulation have been characterized, including WRI18,18,19 and LEC2 (leafy cotyledon2)20 in Arabidopsis. WRI1 is a target of LEC219. The transcription factors regulating fatty acid contents have been identified from soybean in our lab. Two Dof-type (DNA-binding one zinc finger) genes GmDof4 and GmDof11 were found to increase the content of total fatty acids in their transgenic Arabidopsis seeds by activating the ACCase and ACSL (long-chain-acyl CoA synthetase) genes respectively21. Through microarray analysis, a MYB-type gene GmMYB73 was identified and this gene can suppress expression of GL2 (GLABRA 2), a negative regulator of oil accumulations22. Overexpression of GmMYB73 enhanced lipid contents in seeds of transgenic Arabidopsis through release of GL2-inhibited PLDα1 (phospholipase D) expression22,23,24. Overexpression of GmbZIP123 also enhanced lipid content and oil accumulation by regulating two sucrose transporter genes SUC1 and SUC5 and three cell-wall invertase genes cwINV1, cwINV3 and cwINV625. Recently, through RNA-seq analysis, gene co-expression networks have been identified for soybean seed trait regulation and GmNFYA (nuclear transcription factor Y alpha) is found to enhance seed oil contents in transgenic Arabidopsis plants26.
In the present study, a DREB-type (dehydration-responsive element-binding) transcription factor gene GmDREBL, was cloned and found to increase the seed lipid content in the transgenic plants. GmDREBL directly activates the expression of WRI1 to promote fatty acid accumulation. Our study provides a novel viewpoint into the regulation of the fatty acid accumulation in seeds and should add more understanding of the function for the DREB-type transcription factor gene in soybean.
Results
Structural features of GmDREBL and its gene expression
Using high-throughput RNA-seq analysis, 87 transcription factor genes have been identified in developing seeds along with the fatty acid accumulation25. These genes were tested for lipid regulation through transgenic analysis. Among these, one gene encoding an AP2 domain protein of 211 amino acids was further studied. Because the encoded protein was clustered with the DREB subfamily of AP2 family (Fig. 1A), the gene was named as GmDREBL (Glyma12g11150).

Phylogentic analysis of GmDREBL and organic-specific gene expression.
(A) Phylogenetic tree analysis of GmDREBL protein and other DREB proteins. The phylogenetic tree was created with the high similarity sequences of AP2 domains from 6 G. max proteins and 11 Arabidopsis thaliana proteins using PHYLIP and the bootstrap analyses were conducted with 200 replicates. The bootstrap value was showed in the branch and the length of each branch represents the distance between the sequences. (B) The organ-specific expression of GmDREBL in G. max cv. HN44. The roots, shoots and leaves of two weeks seedling were sampled. The pods with 2 cm length and the opening flowers of adult plant were sampled. Two developmental stages of soybean seeds were selected. Weight of a H1 developing seed is only 4% of highest seed fresh weight. Weight of a H5 developing seed is 35% of highest seed fresh weight. (C) The organ-specific expression of AtDREB2A and AtDREB2B in Arabidopsis thaliana. The homologues of GmDREBL with high identity in Arabidopsis were AtDREB2A and AtDREB2B (e-value < 1.3E-30). Their expression was evaluated by publicly available data of microarray in AtGenExpress of TAIR (http://www.arabidopsis.org).
The expression of GmDREBL was examined in different organs of soybean plants. The gene was highly expressed in the H1 and H5 stages of the developing seeds in comparison with the other organs tested (Fig. 1B). The homologues of GmDREBL in Arabidopsis were AtDREB2A (AT5G05410) and AtDREB2B (AT3G11020) (e-vlaue < 1.3E-30). Both of them exhibited high expression in the developing seeds (heart embryo and early cotyledon stages), in addition that AtDREB2A also had high expression in root and stamen of Arabidopsis (Fig. 1C). These results suggest that GmDREBL may be involved in regulation of the seed-related process.
GmDREBL subcellular localization and transcriptional activation
The GmDREBL encoded a putative DREB/AP2-type transcription factor. We then examined the subcellular localization of GmDREBL by transiently expressing the gene in epidermal cells of tobacco leaves. The lower panel of Fig. 2A showed that GmDREBL was located in the nucleus, while the upper panel of Fig. 2A showed that GFP control was mainly observed in the cytoplasm.

Subcellular localization and transactivation activity of GmDREBL.
(A) Subcellular localization of GmDREBL in leaf epidermal cells of tobacco by transient expression assay. (B) Transactivation activity analysis of GmDREBL in Arabidopsis protoplasts. Bars indicate SD (n = 4). **Indicate significant difference compared to BD (P < 0.01).
The transcription activation ability of GmDREBL was further examined by using a dual-luciferase reporter (DLR) assay system in Arabidopsis protoplasts. The coding sequence of GmDREBL and the DNA sequence encoding the GAL4 DNA-binding domain (GAL4DBD) were combined to generate pBD-GmDREBL effector plasmid. The vector containing only the GAL4DBD sequence was used as a negative control and the vector fused with VP16 was used as a positive control. As shown in Fig. 2B, GmDREBL activated the reporter LUC gene, compared to the negative control BD, indicating that GmDREBL has the transcriptional activation activity. Given that the GmDREBL is located in the nucleus and has transcriptional activation activity, this protein is likely a transcription factor.
GmDREBL increased the fatty acid content in seeds of transgenic plants
To investigate the function of GmDREBL during the plant growth and development, the GmDREBL was fused with a GFP gene, driven by 35S promoter and transformed into Arabidopsis plants and the GmDREBL-overexpressing lines were identified by qRT-PCR. We totally screened 22 transgenic lines and most of these lines showed relatively higher levels of GmDREBL expression compared to the Col-0 (Fig. 3A). However, two lines (DL-3 and DL-18) exhibited extremely low expression level (Fig. 3A–C) and may be regarded as vector control for comparison.

The effect of GmDREBL on seed-related traits.
(A) Expression of GmDREBL in transgenic plants. The relative expression level of GmDREBL was normalized using AtACTIN7 as an internal control. Bars indicate SD (n = 4). (B) Examination of GmDREBL presence by PCR in transgenic plant using genomic DNA as a template. An actin 2 gene was also amplified as the control. (C) Detection of GmDREBL-GFP in transgenic plants by Western blot analysis using mouse anti-GFP antibody. And the detection of β actin was used as the control. (D) Overexpression of GmDREBL increased total fatty acid content of transgenic Arabidopsis seeds. Bars indicate SD (n = 4). (E) The profile of fatty acids in dry seeds of Col-0 and three lines DL-2, DL-4 and DL-25. Bars indicate SD (n = 4). (F) The profile of fatty acids in fresh leaves of 12-day-old seedlings from Col-0 and three lines DL-2, DL-4 and DL-25. Bars indicate SD (n = 4). (G) The rosette phenotypes of Col-0 and three homozygous lines DL-2, DL-4 and DL-25 in two and three weeks. (H) The scanning electron micrograph of the seed coat of Col-0 and transgenic plants with GmDREBL. The white line stands for 200 μm. (G) The statistical result of seed length and seed width of wild type Col-0 plant and transgenic plants with GmDREBL. Bars indicate SD (n = 12). (I) Overexpression of GmDREBL increased 1000-seed weight of transgenic Arabidopsis seeds. Bars indicate SD (n = 4). (J) The seed yield per plant of Col-0 and three homozygous lines DL-2, DL-4 and DL-25. Bars indicate SD (n = 10). All the asterisks indicate significant difference compared to the corresponding controls (p-value < 0.05).
Because the RNA-seq-identified transcription factors are roughly correlated with fatty acid accumulation during soybean seed development25, we examined whether the GmDREBL alters the fatty acid accumulation in the overexpressing transgenic lines. Among these lines, 15 overexpression lines exhibited significantly higher total fatty acid content in seeds than vector control (DL-3 and DL-18) and Col-0 (Fig. 3D). The left lines also showed increases in seed total fatty acid contents on average. This result suggests that GmDREBL has a positive role on total fatty acid accumulation in seeds.
To further decipher which composition of fatty acid could be regulated by GmDREBL, we selected three homozygous T3 transgenic lines DL-2, DL-4 and DL-25 for further analysis. The profile and content of fatty acids in the seeds of Col-0 and the three lines were compared. The result showed that overexpression of GmDREBL significantly enhanced the contents of C18:1, C18:2, C18:3 in all the three transgenic lines and the C20:1 content in DL-4 overexpression lines (Fig. 3E). The profile and content of fatty acid were also measured in leaves of the transgenic plants and Col-0 plants and no significant difference was observed between transgenic lines and Col-0 (Fig. 3F). These results indicate that GmDREBL promotes accumulation of fatty acids specifically in seeds, but not in leaves of the transgenic plants.
Overexpression of GmDREBL enhances seed size in transgenic Arabidopsis
We further examined the effects of GmDREBL overexpression on seedling growth and other seed-related traits. We found that there was no difference in rosette size between overexpression lines and Col-0 at different stages of seedling growth (Fig. 3G). The seed size and the 1000 seed-weight were substantially greater in the overexpressing lines than that in Col-0 (Fig. 3H–J). The GmDREBL-GFP protein can be detected in the seed integument cells (Figure S1A). However, the seed yield per plant did not change (Fig. 3K). The length of silique in the three transgenic lines was not significantly different from that in Col-0 either (Figure S1B,C). These results suggest that the GmDREBL increases the seed size/weight but not the total seed yield per plant in transgenic Arabidopsis.
GmDREBL activates expression of WRI1 and other fatty acid biosynthesis-related genes
As the total fatty acids (FA) showed a substantial increase in the seeds of the GmDREBL-overexpressing transgenic plants, it is possible that the GmDREBL regulates the expression of genes involved in FA biosynthesis. We then examined the expression levels of 13 FA-biosynthesis related genes including NAD(P)-binding Rossmann-fold superfamily (At01g24360, At3g03980, At4g13180, At5g18210), KASIII (At01g62640), FAD1 (At01g74960), Beta-ketoacyl synthase (At02g04540), Enoyl-ACP reductase 1 (At02g05990), Thioesterase superfamily (At02g22230, At5g10160), EMB3147 (At02g30200), aldehyde reductase (At3g04000) and KASI (At5g46290). Among these, the gene encoding KASI and the gene (At5g10160) encoding a thioesterase superfamily protein were upregulated in the GmDREBL-overexpressing lines (Fig. 4A). Because these two genes can be directly regulated by WRI1, a key regulator in the biosynthesis of fatty acids8,9,10,19, we further tested the WRI1 expression by qRT-PCR. We find that overexpression of GmDREBL significantly up-regulated WRI1 expression (Fig. 4A). The WRI1 downstream genes FATA, LPD1 and At5g16240 (plant stearoyl-acyl-carrier-protein desaturase family protein) were also enhanced in transgenic plants (Fig. 4A). These results indicate that the GmDREBL promotes expression of WRI1 and other fatty acid biosynthesis-related genes.

GmDREBL can regulate the expression of WRI1.
(A) The expression level of WRI1 and its downstream genes in Col-0 plants and GmDREBL-overexpressing plants. *Indicate significant difference (p-value < 0.05). (B) The intensity of ChIP signal of Col-0 plants and GmDREBL-overexpressing plants with a GFP-tag for GmDREBL. The WRI1 gene promoter is enriched in GmDREBL-overexpressing plants. Bars indicate SD (n = 4). *Indicates significant difference (P < 0.05). (C) The LUC image of tobacco leaves co-infiltrated with A. tumefaciens GV3101 containing different combination of constructs. (D) Quantitative analysis of LUC luminescence intensity in (C). Bars indicate SD (n = 7). *Indicates significant difference (p-value < 0.05). (E) The DRE cis-element present in WRI1 promoter sequence. (F) Gel-shift assay showing that GmDREBL can bind the promoter of WRI1. 100-fold unlabeled probe was used in the third lane. CCGAC motif in Labelled mutant probe was changed to TTATT in fourth lane. Arrows indicate the position of the protein–DNA complex. (G) Illustration of CCGAC motif distribution in the promoters of Glyma15g34770 (WRI1 homologue, e-value = 3.4E-101). (H) GmDREBL enhanced the expression of Glyma15g34770 in GmDREBL-transgenic hairy root of soybean, but not the expression of Glyma08g24420 (WRI1 homologue, e-value = 3.4E-101). *Indicate significant difference (p-value < 0.05).
We further detected whether GmDREBL can directly bind to the promoter of WRI1 by ChIP-qPCR and gel shift assay. Figure 4B showed that, GmDREBL protein expressed in the siliques of the GmDREBL-GFP transgenic plants significantly enriched the promoter region of WRI1, as revealed by anti-GFP antibody. In contrast, the sample of Col-0 plants without GmDREBL protein rarely enriched the WRI1 promoter region. The promoters of WRI1 contain one DRE cis-element of CCGAC (Fig. 4C). We next asked whether GmDREBL could directly bind to the promoters of WRI1. The fragment covering CCGAC motifs and flanking sequence was identified as candidate binding sites in WRI1 promoter. GmDREBL was found to specifically bind to the fragment (Fig. 4D, second and third lane from left). However, GmDREBL could not bind to the mutated version of the fragments (Fig. 4D, fourth lane from left). These results demonstrate that GmDREBL can directly bind to the promoter of WRI1 in plants.
We then examined whether the WRI1 promoter activity was enhanced by GmDREBL using the luciferase expression system in vivo by transiently expressing the constructs in tobacco leaf. The WRI1 promoter: LUC reporter construct (pWRI1:LUC) in pGWB435 or pGWB405-GmDREBL construct (35S:GmDREBL) was transfected into agrobacterium. After culture in LB liquid medium at 28°C overnight, the agrobacteria were collected and suspended in the infiltration buffer. The two transfected agrobacteria were injected into the tobacco leaves solely or together and cultured for 3 days. After that, the leaves were detected by a low-light cooled charge-coupled device imaging apparatus and we find that the GmDREBL enhanced the LUC activity driven by WRI1 promoter (Fig. 4E,F). These results imply that the GmDREBL can stimulate WRI1 promoter activity.
To further confirm that GmDREBL could enhance the expression of WRI1 homologue in soybean. We identified two homologues of WRI1 in soybean, namely Glyma15g34770 (e-value = 3.4E-101) and Glyma08g24420 (e-value = 3.4E-101). We found that two CCGAC cis-elements are present in the promoter of Glyma15g34770 (Fig. 4G). By overexpression of GmDREBL in soybean transgenic hairy roots, we found that GmDREBL could enhance the expression of Glyma15g34770 but not the Glyma08g24420 (Fig. 4H).
GmDREBL is upregulated by GmABI3 and GmABI5
Arabidopsis ABI3 and ABI5 are key regulators for fatty acid accumulation21,27 and ABI3 contains AP2 domain whereas ABI5 belongs to bZIP family. There are putative cis-elements in the promoter region of GmDREBL for possible AP2-domain protein and bZIP protein binding. We then determined if GmDREBL has any co-expressions with GmABI3 (Glyma08g47240) and GmABI5 (Glyma10g08370) from soybean, whose identity with ABI3 and ABI5 is 71.7% and 81.3%, respectively. The expression pattern of GmABI3, GmABI5 and GmDREBL was determined during soybean seed development by real-time PCR and these genes have similar expression pattern (Fig. 5A–C), consistent with the accumulation trend of fatty acids in our previous report25. These results imply that GmDREBL may be regulated by GmABI3 and GmABI5.

GmDREBL is up-regulated by GmABI3 and GmABI5.
(A) The expression level of GmDREBL in different stages of seed development. Six developmental stages of soybean seeds were selected according to the ratio of seed weight of the stage to highest seed fresh weight, namely 4, 8, 12, 20, 35 and 80% (H1–H6). (B) The expression level of GmABI3 in different stages of seed development. (C) The expression level of GmABI5 in different stages of seed development. (D) The expression of GmABI3 in soybean transgenic hairy roots harboring GmABI3. (E) The expression of GmABI5 in soybean transgenic hairy roots harboring GmABI5. (F) The expression of GmDREBL in soybean transgenic hairy roots expressing GmABI3 or GmABI5. Bars indicate SD (n = 4). All the asterisks indicate significant difference compared to the corresponding controls (p-value < 0.05).
We further constructed pBI121-GmABI3 and pBI121-GmABI5 vectors and transfected them into Agrobacterium rhizogenes K599. These agrobacteria were used to infect hypocotyls of soybean seedlings for generation of transgenic hairy roots with overexpression of GmABI3 or GmABI5 (Fig. 5D,E). The expression of GmDREBL was increased in the transgenic hairy roots overexpressing GmABI3 or GmABI5 (Fig. 5F). These results suggest that GmABI3 and GmABI5 promote GmDREBL expression.
GmDREBL is subjected to selection during domestication
Our above study indicates that overexpression of GmDREBL promotes accumulation of fatty acid content in seeds of transgenic plants (Fig. 3). Considering that cultivated soybeans usually had much higher oil contents compared to the wild soybeans, we investigated whether there is any difference in the expression levels of GmDREBL from cultivated soybean and GsDREBL from wild type soybean. It can be seen that the average expression level of GmDREBL in cultivated soybeans is significantly higher than that of GsDREBL in wild soybeans (Fig. 6A), suggesting that the DREBL expression may have been subjected to selection during domestication of soybean. We further determined the relationship between expression levels of DREBL and seed oil contents in 76 wild and cultivated soybeans and found that the seed oil contents were positively correlated with the DREBL gene expressions with a coefficiency of 0.4378 (Fig. 6B).

DREBL expression in wild and cultivated soybeans.
(A) The expression level of DREBL in different wild type and cultivated soybeans. Asterisks indicate significant differences compared with the Col-0 plants (P < 0.05). (B) Correlation between expression of DREBL and lipid content in soybean seeds. r = 0.4278.
Since the DREBL gene expression is relatively higher in the cultivated soybeans and the oil contents are roughly correlated with the DREBL gene expression levels among all the soybeans tested, we determined the sequences of DREBL promoter regions to see if there is any natural variations. The promoters of GmDREBL from 33 cultivated varieties and GsDREBL from 43 wild type soybeans were cloned and sequenced. The phylogenetic tree analysis was performed and we found that most of the wild soybeans were clustered together (Fig. 7, names in black), however, some of the wild soybeans clustered with cultivated soybeans (Fig. 7, names in yellow), possibly suggesting a close relationship of the cultivated soybeans with the few wild soybeans in the same group.

Phylogenetic tree analysis of DREBL promoter in different wild type and cultivated soybeans.
Phylogenetic tree of DREBL promoter sequences from 33 cultivated soybeans and 43 wild soybeans. The yellow names indicated the cultivated soybean accessions and the black names indicated the wild soybean accessions. The tree was constructed by using PHYLIP and the bootstrap analyses were conducted with 1000 replicates.
We further compared the nucleotide diversity among the promoter sequences and found 7 major insertion/deletion (indel) regions (Table 1). Based on the combination of these indels, seven major haplotypes were identified (Table 1). Type 1 had only one cultivar with a specific 15 bp-insertion at −944 position in the DREBL promoter. Type 2 had 19 wild soybeans and 32 cultivated soybeans and Williams 82 belonged to this type 2. Type 1 and 2 had higher oil contents than the Type 3 to 7 and Type 3 to 7 all had wild soybeans (Table 1, Table S1). Type 2 haplotype is very similar to the Type 7 and Type 7 had two more ‘T’ than Type 2. These results indicate that specific indel combinations in cultivated and/or wild soybeans may contribute to the increase of oil contents. It should be noted that the coding sequences of the DREBL gene in the wild and cultivated soybeans did not show much variations28.
We calculated the π value representing the genetic diversity of these promoter sequences and found that wild soybeans had a π value of 0.012299, whereas the cultivated soybeans had a much lower π value of 0.002058, indicating that the wild soybeans have a relatively higher level of genetic diversity in the DREBL gene. These results suggest that the cultivated soybeans may have been selected in the DREBL gene promoters during domestication.
Discussion
Our previous studies have demonstrated that transcription factors GmDof4, GmDof11, GmMYB73, GmbZIP123 and GmNFYA play important roles in regulation of seed-related traits and fatty acid biosynthesis22,25,26,27. Presently, we further found that an AP2-domain protein GmDREBL promotes fatty acid accumulation in seeds of the transgenic Arabidopsis through up-regulation of the master regulatory gene WRI1 and other genes related to fatty acid biosynthesis. The GmDREBL also enhances seed size and seed weight. The expression of the DREBL gene is relatively higher in developing seeds of the cultivated soybeans compared to that in wild soybeans and the expression level is roughly positively correlated with the oil contents in all the soybean accessions. The promoter sequences were further analyzed and the genetic variations in these sequences showed correlations with fatty acid accumulation in soybean seeds.
GmDREBL is located in nucleus and has obvious transcriptional activation ability (Fig. 2). These two features, together with the finding that GmDREBL binds to WRI1 gene promoter and activates this promoter activity (Fig. 4), demonstrate that GmDREBL is a transcription factor directly regulating WRI1 gene expression. In the gene co-expression network study, Glyma15g34770 (WRI1 homologue) was found to be co-expressed with the GmDREBL (Glyma12g11150)29. We found that GmDREBL could enhance the expression of Glyma15g34770 in soybean transgenic hairy root. This suggests that they may participate in the regulatory network for TAG synthesis. Considering that the GmDREBL is abundantly expressed in developing seeds (Fig. 1) and WRIl is a master regulator of lipid biosynthesis, it is most likely that the GmDREBL would activate WRI1 for lipid accumulation especially in seeds. Consistent with this, the lipid contents in leaves of the GmDREBL-overexpressing transgenic plants was not enhanced, probably due to that the gene is barely expressed in leaves (Fig. 1). The GmDREBL localization was also examined in integument cells of GmDREBL-GFP transgenic Arabidopsis seeds and the GmDREBL-GFP seemed to be localized in nucleus and some other regions of the cells (Figure S1A). The localization of the GmDREBL in regions other than the nucleus is probably due to the constitutive expression of the gene driven by the 35S promoter.
Genes in de novo synthesis of FAs was further examined and the expression of KAS1 (b-Ketoacyl-[acylcarrier protein] synthase I) was significantly increased in GmDREBL-overexpressing transgenic lines compared to WT plants (Fig. 4A). KAS1 is responsible for the elongation of fatty acid (FA) synthesis from C4 to C166. Lacking of KAS1 leads to an obvious change in the polar lipid component, abnormal embryo development before the globular stage and sharp decrease in FA levels in seeds. Genes such as Pl-PKb1, PDHE1a, FAD2, FAD3, BCCP2, ACP1, oleosin and KAS1 in fatty acid synthesis have similar expression patterns with WRI1 during the seed developmental process7. These gene expressions increased in WRI1-overexpressing plants but decreased in mutants with down-regulated WRI1 expression. WRI1 is a key regulator in FA biosynthetic pathway. Two BnWRI1 genes, BnWRI1-1 and BnWRI1-2, promoted seed oil increase by 10–40% in transgenic plants overexpressing BnWRI1-1 or BnWRI1-2 and the seed size was also enlarged11. Overexpression of BnSTM resulted in the induction of genes relevant to FA synthesis including BnLEC1, BnLEC2 and BnWRI1 and seed oil content was also increased in the transgenic plants30. In Arabidopsis, WRI1 regulated the expression of BCCP219. BCCP2 is involved in fatty acid biosynthetic process and has acetyl-CoA carboxylase activity. ZmWRI1a, the homologue of AtWRI1 in maize, complemented the reduced fatty acid content of Atwri1-4 mutant and overexpression of ZmWRI1a dramatically increased the fatty acid content in maize grain13. Studies also reported that overexpression of ZmWRI1 resulted in an oil increase12. In the present study, we proved that AtWRI1 or its soybean homologue was upregulated in transgenic plants/transgenic hairy roots overexpressing GmDREBL (Fig. 4A,H). ChIP-qPCR, EMSA and transient transcriptional activation experiments in tobacco leaves (Fig. 4B–F) further demonstrate that GmDREBL can directly bind to the promoter and activate WRI1 expressions, finally leading to FA synthesis and oil accumulation.
It should be noted that there is an ABA-responsive elements (ABRE) in the promoter of GmDREBL and ABI3 and ABI5 are also master regulatory factors for fatty acid biosynthesis21,31. The expression of GmABI3, GmABI5, GmDREBL and GmWRI1 shared the same expression pattern and was in accordance with the fatty acid accumulation trend (Fig. 5). Through soybean-transgenic hairy root experiments, we proved that GmDREBL can be upregulated by GmABI3 and GmABI5. These results indicate that GmABI3 and GmABI5 may be the upstream regulator of GmDREBL.
Because the average DREBL expression level in cultivated soybeans is significantly higher than that in wild soybean plants (Fig. 6A), we compared the promoter regions of the DREBL in the wild and cultivated plants. We identified at least seven haplotypes of DREBL promoters and compared to type 2 containing the reference Williams 82 soybean, the type 1 has two major insertions at −404 and −944 bp positions (Table 1). The type I contains only one cultivar SN30 with high oil content (Table S1), which is clustered with many wild soybeans (Fig. 7). This DREBL allele from SN30 may be further tested in breeding for potential improvement or alteration of oil content. Alternatively, the type 2 allele, which is prevalent in many cultivars, may be introduced into SN30 for further improvement of oil accumulation.
DREB-like transcription factors are generally stress-related and could be induced by abiotic stresses such as drought and high salt32,33,34. We also found that GmDREBL could be sharply induced by drought and high salt (Figure S2A,B) and overexpressing GmDREBL increased the survival rate of transgenic plants under 200 mM NaCl treatment (Figure S2C,D). The GmDREBL could directly bind to the promoter region of stress-related gene LEA and activate LEA expression for stress tolerance (Figure S3). The GmDREBL also promoted expression of the AtDREB2B (AT3G11020) (Figure S1D), which may further contribute to stress tolerance of the transgenic plants.
In conclusion, overexpressing GmDREBL increased fatty acid accumulation in seeds of transgenic plants, likely through the control of a key regulator WRI1 for FA synthesis. Besides, GmDREBL can be regulated by GmABI3 and GmABI5. Our study provides novel insight to the function of DREB-type genes in both oil accumulation and seed size control in addition to their roles in stress response. Further manipulation of the GmDREBL in soybean and evaluation in field test should shed light on its roles in regulation of oil contents in seeds.
Methods
Plant materials and growth conditions
Soybean (G. max) seeds of cultivar Heinong44 were planted in the experimental farm of IGDB, CAS, in Beijing. Arabidopsis plants were grown at 22 °C with a photoperiod of 16 h/8 h (light/dark) per day. All plant materials were harvested and stored at −70 °C for RNA isolation.
Cloning of the GmDREBL gene
The full-length opening reading frame of GmDREBL was amplified from Heinong44 leaf RNA and cloned into the Gateway-T vector (invitrogen, pCR8/GW/TOPO TA Cloning Kit). This construct was confirmed by sequencing. PCR primers were listed in Table S2.
Plasmid construction and protein subcellular localization in Tobacco leaves
35S-GmDREBL-GFP was generated using vector pGWB405 from invitrogen driven by 35S promoter. As GmDREBL has already been cloned into T-vector, the GmDREBL gene was further cloned into the pGWB405 vector through gene recombination, with GFP fusion at the downstream. The pGWB405 vector containing the GFP gene driven by the 35S promoter was used as a control. Each plasmid was transformed into Agrobacterium tumefaciens GV3101. The Agrobacterium strain was incubated in LB medium overnight and re-suspended in infiltration buffer (100 μM acetosyringone, 10 mM MES and 10 mM MgCl2) to an ultimate concentration of 1.0OD=600. And then the strain was infiltrated into tobacco leaves. The infiltrated plant leaves were incubated in greenhouse for 3 days and the subcellular localization of the GmDREBL protein was visualized under a confocal microscope (Leica TCS SP5).
Transcriptional activation analysis in Arabidopsis protoplasts
The transcriptional activation activity was examined in the Arabidopsis protoplast system performed as described previously35. The reporter was a plasmid generated from pUC19 containing the firefly LUC reporter gene and was driven by a modified 35S promoter plus 5 × UAS (upstream activating sequence)36. The GmDREBL gene was fused into the GAL4 DNA BD-coding sequence and constructed into pRT107 to generate an effector plasmid pRT-BD-GmDREBL under the control of 35S promoter. pRT107 vector containing the BD sequence was used as a negative control and the vector containing the BD-VP16 fusion sequence as a positive control. A pPTRL plasmid containing a CaMV 35S promoter and Renilla LUC was used as an internal control. The constructs above were transformed into Arabidopsis protoplasts by PEG-mediated transfection. Using a GloMaxTM 20/20 Luminometer (Promega), luciferase activity was measured for each co-transfection sample after culturing for 16–20 h.
Generation of GmDREBL transgenic Arabidopsis plants
The pGWB405-GmDREBL-GFP vector was transfected into A. tumefaciens GV3101 by electroporation, then introduced into Arabidopsis Col-0 plants by the vacuum infiltration method37. Homozygous transgenic lines were obtained and the lines DL-2, DL-4 and DL-25 with different expression levels were selected for further analysis. The subcellular localization of the GmDREBL protein in integument cells of seed was visualized under a confocal microscope (Leica TCS SP5). Positiveness of transgenic plant was examined by PCR using plant DNA as a template and an AtActin-2 gene was amplified as a control. Western blot was performed using GFP-Tag mouse antibody (Abmart) and plant β actin mouse monoclonal antibody (Cwbiotech).
Measurement of seed size and weight per thousand seeds
Seeds from different transgenic lines were coated by gold dust and pictures were taken under a scanning electron microscope. The length and width of 20 seeds were measured using ImageJ program. Weight per thousand seeds was obtained through weighing 1000 seeds.
Quantitative analysis of Fatty Acid content
Seeds (10 mg) from Col-0 and transgenic lines, with four biological replicates, were used for extraction of fatty acids as previously described38. After extraction, the fatty acids were subjected to gas chromatography (GC2014, Shimadzu). Peaks corresponding to each FA species can be identified by FAME analytical standard (Cat. no. 18920-1AMP, Supelco).
Quantitative reverse transcriptase-PCR (qRT-PCR)
According to the instructions, total RNA was isolated from siliques of Col-0 and the transgenic lines using TRIzol reagent (Tiangen). The reverse transcription was carried out using a first-strand cDNA synthesis kit (TransGen Biotech). The cDNA was then used as templates for qRT-PCR using SYBR qPCR mix (Toyobo). After reaction on a LightCycler480 System (Roche), the relative expression level of each sample was quantified using an internal control. The UKN1 (Glyma12g02310) gene and AtACTIN7 gene were chosen as an internal control for soybean (Glycine max) and Arabidopsis system, respectively. All the qRT-PCR primers are listed in Table S2.
Transactivation of target promoter by GmDREBL in tobacco leaves
Using Gateway® technology (Invitrogen), the 3 kb sequences upstream from the ATG codons of WRI1 were inserted into pGWB435 to generate promoter: LUC reporter constructs. The 35S:GmDREBL plasmid and the reporter plasmid were transformed into A. tumefaciens GV3101. The Agrobacterium strains were cultured in LB medium with spectinomycin and rifampicin at 28 °C. After culture, the harvested Agrobacterium were re-suspended in infiltration buffer containing 10 mM MES, 0.2 mM acetosyringone and 10 mM MgCl2 with pH5.7 until the OD600 concentration reached 1.0. Using a syringe without needle, equal amounts of various combined bacterial suspensions were injected into the young leaves of the five-week-old tobacco plants and the plants were cultured at 24 °C for 3 days. Then the injected leaves were cut off, spread with 100 mM luciferin (Promega) and placed in darkness for 5 min. Then the LUC activity was detected with a low-light cooled charge-coupled device imaging apparatus (iXon; Andor Technology). At least six independent biological replicates were performed for each experiment.
ChIP-PCR assay
ChIP assays were carried out according to a published protocol39 with modifications. Siliques (3 g) of the GmDREBL-GFP-transgenic lines and Col-0 were cross-linked in 1% formaldehyde and the chromatin was isolated based on the previous procedures39. Using GFP antibody (EarthOX), the DNA-protein complex was immunoprecipitated. The precipitated DNA was further subjected to quantitative PCR analysis.
Gel-shift assay
The recombinant protein of maltose binding protein–GmDREBL was expressed in Escherichia coli BL21 using pMAL-c5x vector and purified from cells using Amylose Resin (NEB). The examined fragments in promoters of target genes were annealed by using synthesized oligonucleotides. The gel-shift assay was performed using a LightShift Chemiluminescent EMSA Kit (Thermo) according to the manufacturer’s instructions. The sequences of oligonucleotides are listed in Table S2.
The expression of GmWRI1 homologues, GmDREBL, GmABI3 and GmABI5 in soybean transgenic hairy roots expressing the GmDREBL, GmABI3 and GmABI5
pBI121-GmDREBL, pBI121-GmABI3 and pBI121-GmABI5 vectors were constructed and were transfected into A. rhizogenes K599 respectively. One-week-old seedlings of soybean Kefeng 1 were infected with K599 or K599 harboring the construct mentioned above following previous description40,41. Hairy roots were generated at the infection sites after 14 d and the seedlings were immersed in water for 3 d and then the original main roots were removed by cutting. Hairy roots of ~1 cm in length were collected and RNAs were extracted for further determination of GmWRI1 homolgoues, GmDREBL, GmABI3 and GmABI5 expression level. All PCR primers were listed in Table S2.
Phylogenetic tree analysis of DREBL gene promoters from wild type and cultivated soybean
DNA was extracted from wild type and cultivated soybeans and promoters of GmDREBL from cultivated soybean and promoters of GsDREBL from wild type soybean were cloned, sequenced and compared. Phylogenetic tree analysis was performed using PHYLIP (V. 3.69).
Comparison of DREBL expression in cultivated soybean and wild type soybean
RNA was extracted from developing seeds of 90 cultivated and 69 wild type soybeans and cDNA was made through reverse transcription. The DREBL expression was examined through real-time PCR.
Additional Information
How to cite this article: Zhang, Y.-Q. et al. Soybean GmDREBL Increases Lipid Content in Seeds of Transgenic Arabidopsis. Sci. Rep. 6, 34307; doi: 10.1038/srep34307 (2016).
References
MacKenzie, S. L. Current developments in the modification of oilseed composition. Outlook Agr. 24, 213–218 (1995).
Ohlrogge, J. B. & Jaworski, J. G. Regulation of fatty acid synthesis. Annu. Rev. Plant Phys. 48, 109–136 (1997).
Jackowski, S. & Rock, C. O. Acetoacetyl-Acyl carrier protein synthase, a potential regulator of fatty-acid biosynthesis in Bacteria. J. Biol. Chem. 262, 7927–7931 (1987).
Clough, R. C., Matthis, A. L., Barnum, S. R. & Jaworski, J. G. Purification and characterization of 3-Ketoacyl-Acyl carrier protein synthase-iii from Spinach - a condensing enzyme utilizing Acetyl-Coenzyme-a to initiate fatty-acid synthesis. J. Biol. Chem. 267, 20992–20998 (1992).
Shimakata, T. & Stumpf, P. K. Isolation and function of Spinach leaf beta-Ketoacyl-[Acyl-Carrier-Protein] synthases. Proc. Natl. Acad. Sci. USA 79, 5808–5812 (1982).
Wu, G. Z. & Xue, H. W. Arabidopsis beta-Ketoacyl-[Acyl Carrier Protein] synthase i is crucial for fatty acid synthesis and plays a role in chloroplast division and embryo development. Plant Cell 22, 3726–3744 (2010).
Ruuska, S. A., Girke, T., Benning, C. & Ohlrogge, J. B. Contrapuntal networks of gene expression during Arabidopsis seed filling. Plant Cell 14, 1191–1206 (2002).
Cernac, A. & Benning, C. WRINKLED1 encodes an AP2/EREB domain protein involved in the control of storage compound biosynthesis in Arabidopsis. Plant J. 40, 575–585 (2004).
Maeo, K. et al. An AP2-type transcription factor, WRINKLED1, of Arabidopsis thaliana binds to the AW-box sequence conserved among proximal upstream regions of genes involved in fatty acid synthesis. Plant J. 60, 476–487 (2009).
Baud, S., Wuilleme, S., To, A., Rochat, C. & Lepiniec, L. Role of WRINKLED1 in the transcriptional regulation of glycolytic and fatty acid biosynthetic genes in Arabidopsis. Plant J. 60, 933–947 (2009).
Liu, J. et al. Increasing seed mass and oil content in transgenic Arabidopsis by the overexpression of WRI1-like gene from Brassica napus. Plant Physiol. Bioch. 48, 9–15 (2010).
Shen, B. et al. Expression of ZmLEC1 and ZmWRI1 increases seed oil production in maize. Plant Physiol. 153, 980–987 (2010).
Pouvreau, B. et al. Duplicate maize wrinkled1 transcription factors activate target genes involved in seed oil biosynthesis. Plant Physiol. 156, 674–686 (2011).
Tajima, D. et al. Wrinkled 1 (WRI1) homologs, AP2-type transcription factors involving master regulation of seed storage oil synthesis in castor bean (Ricinus communis L.). Am. J. Plant Sci. 04, 7 (2013).
Shintani, D., Roesler, K., Shorrosh, B., Savage, L. & Ohlrogge, J. Antisense expression and overexpression of biotin carboxylase in tobacco leaves. Plant Physiol. 114, 881–886 (1997).
Dehesh, K., Tai, H., Edwards, P., Byrne, J. & Jaworski, J. G. Overexpression of 3-ketoacyl-acyl-carrier protein synthase IIIs in plants reduces the rate of lipid synthesis. Plant Physiol. 125, 1103–1114 (2001).
Riechmann, J. L. & Meyerowitz, E. M. The AP2/EREBP family of plant transcription factors. Biol. Chem. 379, 633–646 (1998).
Masaki, T. et al. ACTIVATOR of Spo(min):: LUC1/WRINKLED1 of a Arabidopsis thaliana transactivates sugar-inducible promoters. Plant Cell Physiol. 46, 547–556 (2005).
Baud, S. et al. WRINKLED1 specifies the regulatory action of LEAFY COTYLEDON2 towards fatty acid metabolism during seed maturation in Arabidopsis. Plant J. 50, 825–838 (2007).
Santos-Mendoza, M. et al. Deciphering gene regulatory networks that control seed development and maturation in Arabidopsis. Plant J. 54, 608–620 (2008).
Wang, H. Y., Guo, J. H., Lambert, K. N. & Lin, Y. Developmental control of Arabidopsis seed oil biosynthesis. Planta 226, 773–783 (2007).
Liu, Y. F. et al. Soybean GmMYB73 promotes lipid accumulation in transgenic plants. BMC Plant Biol. 14, 73 (2014).
Shen, B., Sinkevicius, K. W., Selinger, D. A. & Tarczynski, M. C. The homeobox gene GLABRA2 affects seed oil content in Arabidopsis. Plant Mol. Biol. 60, 377–387 (2006).
Shi, L., Katavic, V., Yu, Y. Y., Kunst, L. & Haughn, G. Arabidopsis glabra2 mutant seeds deficient in mucilage biosynthesis produce more oil. Plant J. 69, 37–46 (2012).
Song, Q. X. et al. Soybean GmbZIP123 gene enhances lipid content in the seeds of transgenic Arabidopsis plants. J. Exp. Bot. 64, 4329–4341 (2013).
Lu, X. et al. The transcriptomic signature of developing soybean seeds reveals genetic basis of seed trait adaptation during domestication. Plant J. 86, 530–544 (2016).
Wang, H. W. et al. The soybean Dof-type transcription factor genes, GmDof4 and GmDof11, enhance lipid content in the seeds of transgenic Arabidopsis plants. Plant J. 52, 716–729 (2007).
Zhou, Z. K. et al. Resequencing 302 wild and cultivated accessions identifies genes related to domestication and improvement in soybean. Nat. Biotechnol. 33, 408–U125 (2015).
Yu, J. et al. SFGD: a comprehensive platform for mining functional information from soybean transcriptome data and its use in identifying acyl-lipid metabolism pathways. BMC Genomics 15, 271 (2014).
Elhiti, M. et al. Altered seed oil and glucosinolate levels in transgenic plants overexpressing the Brassica napus SHOOTMERISTEMLESS gene. J. Exp. Bot. 63, 4447–4461 (2012).
Penfield, S., Li, Y., Gilday, A. D., Graham, S. & Graham, I. A. Arabidopsis ABA INSENSITIVE4 regulates lipid mobilization in the embryo and reveals repression of seed germination by the endosperm. Plant Cell 18, 1887–1899 (2006).
Liu, Q. et al. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 10, 1391–1406 (1998).
Liu, Q., Zhao, N. M., Yamaguch-Shinozaki, K. & Shinozaki, K. Regulatory role of DREB transcription factors in plant drought, salt and cold tolerance. Chinese Sci. Bull. 45, 970–975 (2000).
Yamaguchi-Shinozaki, K. & Shinozaki, K. DREB regulons in abiotic-stress-responsive gene expression in plants. Mol. Breeding of Forage and Turf. 15–27 (2009).
Wei, W. et al. Soybean GmPHD-type transcription regulators improve stress tolerance in transgenic Arabidopsis plants. Plos One 4, e7209 (2009).
Ohta, M., Matsui, K., Hiratsu, K., Shinshi, H. & Ohme-Takagi, M. Repression domains of class II ERF transcriptional repressors share an essential motif for active repression. Plant Cell 13, 1959–1968 (2001).
Bechtold, N. & Pelletier, G. In planta Agrobacterium-mediated transformation of adult Arabidopsis thaliana plants by vacuum infiltration. Methods Mol. Biol. 82, 259–266 (1998).
Poirier, Y., Ventre, G. & Caldelari, D. Increased flow of fatty acids toward beta-oxidation in developing seeds of Arabidopsis deficient in diacylglycerol acyltransferase activity or synthesizing medium-chain-length fatty acids. Plant Physiol. 121, 1359–1366 (1999).
Saleh, A., Alvarez-Venegas, R. & Avramova, Z. An efficient chromatin immunoprecipitation (ChIP) protocol for studying histone modifications in Arabidopsis plants. Nat. Protoc. 3, 1018–1025 (2008).
Kereszt, A. et al. Agrobacterium rhizogenes - mediated transformation of soybean to study root biology. Nat. Protoc. 2, 948–952 (2007).
Hao, Y. J. et al. Soybean NAC transcription factors promote abiotic stress tolerance and lateral root formation in transgenic plants. Plant J. 68, 302–313 (2011).
Acknowledgements
This work is supported by the Transgenic Research Projects (2014ZX0800926B), the 973 projects (2013CB835205, 2015CB755702), the Transgenic Research Projects (2016ZX08009-003-004), the CAS leading projects (XDA08020106), the National Natural Science Foundation of China (31530004 and 31260568 [to FYZ]) and the State Key Lab of Plant Genomics.
Author information
Authors and Affiliations
Contributions
Y.-Q.Z. and X.L. performed the experiments; F.-Y.Z., Q.-T.L., W.W., S.-L.N., W.-K.Z. and B.M. contributed to some material preparation and analysis; Y.-Q.Z., X.L. and J.-S.Z. wrote the article; all authors reviewed the manuscript. J.-S.Z. and S.-Y.C. conceived the project and obtained funding.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zhang, YQ., Lu, X., Zhao, FY. et al. Soybean GmDREBL Increases Lipid Content in Seeds of Transgenic Arabidopsis. Sci Rep 6, 34307 (2016). https://doi.org/10.1038/srep34307
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep34307
This article is cited by
-
Metabolic Perspective on Soybean and Its Potential Impacts on Digital Breeding: An Updated Overview
Journal of Plant Biology (2024)
-
WGCNA and transcriptome profiling reveal hub genes for key development stage seed size/oil content between wild and cultivated soybean
BMC Genomics (2023)
-
Regulation of seed traits in soybean
aBIOTECH (2023)
-
4D genetic networks reveal the genetic basis of metabolites and seed oil-related traits in 398 soybean RILs
Biotechnology for Biofuels and Bioproducts (2022)
-
Ectopic expression of MYB repressor GmMYB3a improves drought tolerance and productivity of transgenic peanuts (Arachis hypogaea L.) under conditions of water deficit
Transgenic Research (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
