
Abstract
A new species of fossil conifer wood, Podocarpoxylon donghuaiense sp. nov., is described from the late Eocene of Nadu Formation in Baise Basin of the Guangxi Province, South China. This fossil wood is characterized by distinct growth rings, circular to oval tracheids in cross section, 1–2-seriate opposite pits on radial tracheid walls, uniseriate (rarely biseriate) rays, smooth end walls of ray parenchyma cells and the absence of resin ducts, suggesting its affinity to Podocarpaceae. The new species is distinctive from other Cenozoic woods ascribed to this family by the combination of distinctive growth rings, the absence of axial parenchyma, the occurrence of bordered pits on tangential tracheid walls and the occurrence of 3–4 cuppressoid or taxodioid pits on cross-fields. This represents the first record of podocarpoid fossil wood in South China and provides fossil evidence for the early dispersal and diversification of Podocarpaceae in eastern Asia as well as for mild temperate seasonal climate in this region during the late Eocene.
Similar content being viewed by others


Introduction
As the second largest family within the conifers, the modern Podocarpaceae largely comprises evergreen trees and shrubs belonging to 194 species within 19 genera1. This family is mainly distributed in tropical and subtropical regions from central to South America, Africa (include Madagascar), Indochina through Malesia to Australia and Oceania1,2, extending as far north as China and Japan as well as to Mexico and the Caribbean3,4,5. Podocarpaceae are most abundant in montane tropical rainforests, but occasionally occur in lowland forests6. In comparison with extensive investigations on extant phylogenetics and geography of the family, its early evolution and migration in deep time are still poorly known, mainly due to the lack of megafossils, especially of Cenozoic age.
Fossil records of podocarps from the Mesozoic were well documented in both the Northern and Southern hemispheres. Molecular and fossil evidence suggests that the podocarps originated during Triassic-Jurassic time in Gondwana and apparently spread to the Northern Hemisphere during the Jurassic7. Although the family appears to be of ancient origin, molecular dating analysis suggests that the majority of extant genera have arisen relatively recently in the Upper Cretaceous to Cenozoic8.
Although modern Podocarpaceae are widespread mostly in the Southern Hemisphere, there are numerous reports of Cenozoic megafossils and palynomorphs attributable to this family from the Northern Hemisphere. Podocarpus shoots were found in Eocene deposits of Tennessee9, whereas pollen seemingly belonging to this genus is known from the Miocene of Oregon and Idaho10 and late Eocene of Colorado11. In Europe, leaves of Prumnopitys have been reported from the Eocene of England12, while leaf fossils assigned to Podocarpus have been found in Eocene to Oligocene deposits of Ukraine13,14 and of the southeastern regions of European Russia15. A phylloclade of Phyllocladus has also been described by Krasnov13 from Eocene deposits of the Kharkov region (Ukraine). The assignment of these fossils to Podocarpus and Phyllocladus need special examination, however. In particular, several records of Podocarpus-like leaves from Eocene to Miocene localities of central and eastern Europe ascribed to the species Podocarpus eocenica Ung. have been reconsidered by Ferguson et al.16 as members of Amentotaxus, a genus of the Taxaceae. In addition, podocarpoid fossil wood has been reported from the Oligocene-Miocene of southern Ural17. Podocarpaceae palynomorphs occur widely in Europe from the Paleocene to the Miocene18,19,20, whereas the records of Podocarpus pollen from the Pliocene of Iceland21 and Pleistocene of Belgium21,22 possibly result from redeposition from older beds19.
In Asia, several fossil woods attributed to Podocarpoxylon are known from the early Tertiary to Pliocene of India23. Podocarpus leaves have been reported from the Oligocene of Assam24, while Nageia leaves have also been described from Eocene deposits of Hainan Island and Guangdong Province, South China25,26. Podocarpaceae fossil wood of Cenozoic age has not been recorded in China before but several apparent podocarpaceous woods, including Podocarpus (Nageia) nagi Pilger27 have been reported from Lower Cretaceous deposits28,29. As for microfossils, Paleocene palynological records of Podocarpaceae locally occur in India, but they are very poorly represented in Southeast Asia where the pollen of Podocarpus sensu lato appears in the late Eocene7. In northern Kazakhstan, Podocarpaceae pollen grains have also been reported from the Eocene, but they disappear there in the late Oligocene30.
Here we describe a new fossil wood of Podocarpoxylon from the late Eocene Nadu Formation of Baise Basin, Guangxi Province of South China and review the phytogeographic and ecological implications of our finding. This is the first occurrence of fossil Cenozoic Podocarpaceae wood from South China.
Results
Systematics
Order: Araucariales Gorozh.
Family: Podocarpaceae Endl.
Genus: Podocarpoxylon Gothan, 1908.
Species: Podocarpoxylon donghuaiense Li, Jin, Quan et Oskolski, sp. nov.
Etymology
The specific name “Donghuai” is the locality name where the fossils were collected.
Holotype
DHW006.
Paratypes
DHW001 to DHW005, DHW007 to DHW0013.
Repository
Fossil wood samples and microscopic slides are deposited in the Museum of Biology, Sun Yat-sen University, Guangzhou, China.
Type locality and horizon
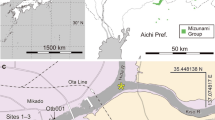
Nadu Fm., late Eocene. Specimens were collected in Donghuai coal-mine. Baise City, Guangxi Province (Fig. 1).

Location of the study area.
Location of Baise Basin, Guangxi Province, South China. (drawn by L.L., using Adobe Photoshop CS5).
Diagnosis
Growth rings with prominent latewood, transition from earlywood to latewood gradual; circular pits on radial tracheid walls uniseriate, sometimes biseriate opposite, circular pits rarely occur in tangential walls of tracheids; cross-field pits cupressoid and taxodioid, circular to oval, 1–4 pits (mean 2) in opposite 1–2 rows per cross-field; axial parenchyma absent; rays predominately uniseriate, rarely partially biseriate; ray cells with smooth horizontal and tangential walls; ray tracheids absent; resin ducts absent.
Description
Growth rings distinct, 5.2–7.9 mm wide, with prominent latewood, transition from earlywood to latewood gradual (Fig. 2A). Earlywood tracheids thin-walled, circular to oval in cross-sectional outline, 23–40 μm (mean 32 μm) in tangential diameter and 23–56 μm (mean 37 μm) in radial size (Fig. 2B). Latewood tracheids moderately thick-walled, circular to oblong in cross-sectional outline, 7–23 μm (mean 13 μm) in tangential diameter and 11–21 μm (mean 15 μm) in radial size.

Wood structure of Podocarpoxylon donghuaiense sp. nov. (DHW006).
(A) Transverse section showing distinct growth rings (arrow). Scale bar = 400 μm. (B) Transverse section of early wood showing circular to oval tracheids. Scale bar = 100 μm. (C) Tangential section showing the predominately uniseriate rays. Scale bar = 100 μm. (D) Tangential section of late wood showing spiral thickenings in tracheids (arrow). Scale bar = 100 μm. (E) Radial section showing 1–2 seriate opposite pits in radial tracheid walls (arrow). Scale bar = 20 μm. (F) Radial section showing spiral and branched thickenings on tracheid walls. Scale bar = 20 μm. (G) Tangential section showing partially 2-seriate rays (arrow). Scale bar = 50 μm. (H) Radial section showing cupressoid and taxodioid pits on cross-fields. Scale bar = 20 μm. (I) Radial section showing cupressoid pits on cross-fields. Scale bar = 10 μm. (J) Radial section showing taxodioidpits on cross-fields. Scale bar = 10 μm.
Pits in radial tracheid walls uniseriate, occsionally also biseriate opposite (Fig. 2E). Pits bordered, circular (11–17 μm in diameter with the average of 15.7 μm) to oval (14–20 μm in size) in outline. Crassulae absent. Bordered pits of 8.2–15.0 μm in diameter rarely occur also on the tangential walls of latewood tracheids. Well-developed tightly spaced spiral and branched thickenings present on radial and tangential walls of latewood tracheids (Fig. 2D,F), with angles to the tracheid axes ranging from 40° to 60°.
Axial parenchyma absent.
Rays 42–331 μm (mean 100 μm) in height, predominately 1-seriate (Fig. 2C,D) and rarely partially 2-seriate (Fig. 2G), completely composed of parenchyma cells, 1–16 cells (mean 5 cells). Ray cells oval or elliptical in tangential section, both vertical and horizontal walls of ray parenchyma cells smooth, indentures absent. Cross-field pits possibly cupressoid or taxodioid type, circular to oval of 6–12 μm in size, with 1–4 pits (mean 2) per cross-field (Fig. 2H–J). Crystals not found. Resin ducts absent.
Discussion
Comparison with modern materials
Within the conifer families, the fossil wood from Donghuai may not be placed into Pinaceae, Cupressaceae or Cephalotaxaceae, as it has no resin ducts, axial parenchyma and ray tracheids. This late Eocene wood is also distinctive both from Araucariaceae by the absence of two or more seriate alternate intertracheary pitting and by the lacking crowded araucarioid cross-field pits and from Sciadopityaceae by the absence of window-like cross-field pits. Therefore, it remains for us to consider the relationships of this fossil wood to Taxaceae or Podocarpaceae31,32,33,34.
The presence of spiral and branched thickenings on the walls of latewood tracheids in combination with smooth horizontal and tangential walls of ray cells suggests the fossil wood from Donghuai may be considered as a member of Taxaceae31,32,33,34. After careful examination of the thickenings on tracheid walls in this sample we suggest, however, that these structures are not of the same nature as the spiral tertiary thickenings that occur in all modern genera of this family, with the exception of Austrotaxus R.H. Compton31,34. Extant Taxaceae show finer spiral thickenings, more widely spaced and tilted at lower angles (usually not exceeding 45°) in respect to the tracheid axes than the thickenings observed in the late Eocene wood from Donghuai. Such features of the fossil wood as tracheids bearing spiral thickenings confined only to the latewood and their absence in the earlywood has also not been reported in any modern Taxaceae31,32,33,34. Therefore, the spiral and branched structures occurring in tracheids of the Donghuai wood are more likely artifacts (probably, effects of wood compression) than true tertiary thickenings. The occurrences of similar artifacts in fossil woods seemingly belonging to Taxaceae have been surveyed by Afonin & Philippe35. In fossil woods assigned to Podocarpaceae these structures have been reported by Chudajberdyev17 in tracheids of Podocarpoxylon uralense Chudajb.
As far the thickenings on tracheid walls are recognized as artifacts rather than diagnostic traits, the suite of other characters (smooth horizontal and tangential walls of ray cells without indentures, cupressoid and taxodioid pits on cross fields) indicates that the fossil wood reported here has greatest affinity to the Podocarpaceae31,32,33,34. This family, however, shows considerable diversity of wood structure that does not allow distinction between its genera using wood anatomical traits. The late Eocene wood from Donghuai is different from most modern Podocarpaceae in growth rings with conspicuous latewood, lacking axial parenchyma and the occurrence of 1–4 pits on the cross-field. However, the presence of prominent latewood portion has been reported in growth rings of Podocarpus acutifolius Kirk, Podocarpus macrophyllus (Thunb.) D. Don., Lagarostrobus (Dacrydium) franklinii (Hook. f.) Quinn, Halocarpus (Dacrydium) bidwillii (Hook. f. ex T. Kirk) Quinn, Phyllocladus glaucus Carr. and P. trichomanoides D. Don32,34,36,37,38. Axial parenchyma is also lacking in Dacrydium elatum (Roxb.) Wall. ex. Hook., D. colensoi Hook., D. biforme (Hook.) Pilg., D. kirkii F. Muell. ex Parl., D. intermedium Kirk, D. taxifolium Banks & Sol. ex D. Don, Halocarpus bidwillii Lepidothamnus intermedius (Kirk) Quinn., Manoao colensoi (Hook.) Molloy., Microcachrys tetragona (Hook.) Hook., Phyllocladus alpinus Hook.f., P. glaucus, P. trichomanoides, Podocarpus elongatus (Aiton) L’Hér. ex Pers., Prumnopitys harmsiana (Pilg.) de Laub. and P. taxifolia (Sol. ex D.Don) de Laub31,32,33,36,37,39,40. Although cross-fields with 1–2 pits are the most common condition in Podocarpaceae, the occurrence of cross-fields with up to 4 cupressoid or taxodioid pits has been reported for Dacrydium pierrei, D. intermedium, Podocarpus hallii, Microcachrys tetragona and Retrophyllum minor (Carrière) C. N. Page32,34,36,37,39. Therefore, the fossil wood from Donghuai shows most resemblance to some species of Prumnopytis (especially P. taxifolia) as well as to some members of Podocarpus, Dacrydium and Microcachrys, but it cannot be convincingly placed in any extant genus of Podocarpaceae on the basis of its anatomical traits.
Comparison with fossil materials
The fossil wood described here is characterized by an absence of axial parenchyma and by the cross-fields with cupressoid and/or taxodioid pits. Within fossil woods ascribed to Podocarpaceae, these traits are found in some species of the genus Podocarpoxylon Gothan31,32,41, as well as in the monospecific genus Prumnopityoxylon Franco & Brea that was recently described from the Upper Cenozoic of Argentina42. Moreover, this combination of wood traits has also been reported for Phyllocladoxylon annulatus Patton from the Oligocene of Australia43. The fossil sample from Donghuai exhibits similarities to Cenozoic species of Podocarpoxylon as well as with Prumnopityoxylon gnaedingerae Franco & Brea and Phyllocladoxylon annulatus Patton (Table 1).
Within seven species that have no axial parenchyma (Prumnopityoxylon gnaedingerae Franco & Brea, Podocarpoxylon aparenchymatosum Gothan, P. dusenii Kräusel, P. latrobensis Greenwood, P. sahnii (Ramanujam) Trivedi & Srivastava, P. tiruvakkaraianum (Ramanujam) Trivedi & Srivastava and Phyllocladoxylon annulatus Patton), cross-fields with more than two pits occur only in P. aparenchymatosum and P. gnaedingerae. Podocarpoxylon aparenchymatosum from the Eocene deposits of Antarctica44,45 differs, however, from the Donghuai wood in possessing 3-seriate pitting on radial tracheid walls. Unlike Prumnopityoxylon gnaedingerae, the Donghuai wood sample shows distinct growth rings and by higher rays height with the occasional occurrence of bi-seriate portions.
It follows from its unique character combinations that the late Eocene wood from Donghuai can be recognized as a new species named here as Podocarpoxylon donghuaiense. Although this fossil wood shows certain similarity to Prumnopityoxylon, Franco & Brea’s42 diagnosis of this genus lacks sufficient detail to separate it from Podocarpoxylon. Any of the wood traits, that have been considered by these authors as diagnostic for Prumnopityoxylon (i.e. “slightly distinct or indistinct growth rings; absence of axial parenchyma; uniseriate and homocellular rays; uniseriate or biseriate, opposite or sub-opposite, separate or contiguous tracheid pitting; taxodioid or cuppressoid cross-field pitting, with 1–5 bordered pits per field”)42, can be also found elsewhere in Podocarpoxylon and their occurrences are consistent with Gothan’s44 diagnosis of this genus. For this reason, we ascribe the new species to Podocarpoxylon rather than to Prumnopityoxylon.
Biogeographic implications
Podocarpoxylon donghuaiense sp. nov. is the first record of podocarpoid fossil wood in China. Coeval macrofossils assigned to Podocarpaceae have already been reported from the South China: well-preserved leaves of Nageia have recently been described from the Eocene Changchang Formation of Hainan Island and the Eocene Youganwo and Huangniuling formations of Guangdong Province25,26. Podocarpoxylon donghuaiense is distinct, however, from the wood of extant Nageia species by the absence of axial parenchyma32,33,34. Thus it is very unlikely that the fossil wood from Guangxi belonged to the same plant taxon as the fossil leaves from Hainan and Guangdong, despite the geographical proximity and nearly the same age of these three findings.
Jin et al.’s25 and Liu et al.’s26, as well as our data confirm that the common occurrence of the Podocarpaceae species in the late Eocene vegetation of South China, that is consistent with the age of initial appearance of this family in Southeast Asia estimated by palynological records7. As the results of molecular dating suggest8, diversification of Podocarpaceae during Eocene gave rise to such modern genera distributed now in this region, as Dacrydium, Podocarpus and Nageia. Although Podocarpoxylon donghuaiense shares some wood traits with some extant species of Podocarpus and Dacrydium, this fossil wood can be convincingly ascribed to neither of them, probably because these taxa had not yet been emerged as distinct entities in the late Eocene.
Most modern species of Podocarpaceae have growth rings with indistinct to very narrow latewood, or growth rings lacking. Contrastingly, the growth rings of Podocarpoxylon donghuaiense show relatively high proportion of latewood with gradual transition from earlywood to latewood. These traits are indicative for plants with long growing period without rapid shift to seasonal dormancy that are encountered mainly in the middle latitudes of both hemispheres46,47,48. Within modern Podocarpaceae, this type of growth rings has been reported only in six species (Podocarpus macrophyllus ranged from Myanmar through mainland China and Taiwan to Japan, Podocarpus acutifolius, Halocarpus bidwillii, Phyllocladus glaucus and P. trichomanoides from New Zealand and Lagarostrobus franklinii from Tasmania49) which are restricted to the temperate regions without dry season, with hot or warm summer (Cfa and Cfb climate types of Köppen’s classification)50. The occurrence of Podocarpaceae wood with prominent latewood in distinctive growth rings may therefore be an indicator that South China had mild temperate seasonal climate during the late Eocene.
Conclusion
A new species of fossil conifer wood, Podocarpoxylon donghuaiense sp. nov., is described from the late Eocene of Baise Basin of Southern China. This is the first Cenozoic record of podocarpoid fossil wood in Southern China. The new species is distinguished from other Cenozoic woods ascribed to Podocarpaceae by the unique combination of distinctive growth rings, the absence of axial parenchyma, the lack of bordered pits on tangential tracheid walls and the occurrence of 3–4 cuppressoid or taxodioid pits on cross-fields. The tightly spaced thickenings on latewood tracheid walls of the fossil wood are recognized as artifacts (probably, effects of wood compression) rather than true tertiary thickenings. Therefore, Podocarpoxylon donghuaiense sp. nov. provides the first robust physical evidence for the early steps of dispersal and diversification of Podocarpaceae in eastern Asia, as well as for warm and wet climate in this region and the presence of clear growth rings also suggest this was a seasonal climate during the late Eocene.
Methods
The fossil woods described here were collected from the Nadu Formation outcropping at Donghuai Coal-mine in the west part of the Baise Basin (transliterated also as the Bose Basin by some authors), Guangxi, South China (23° 52′ 14.84″N, 106° 34′ 49.27″E, Fig. 1). The geological age of the formation is believed to be late Eocene based on the well-studied Nadu Mammalian Fauna51.
The well-preserved wood specimens examined represent portions of a main trunk (DHW001-013). The holotype DHW006 is about 10 cm diameter by 15 cm in length. Thin-sections were prepared according to standard methods of cutting, grinding and polishing using different grades of carborundum powder52. Wood slides were examined using a Carl Zeiss Axio Scope A 1 light microscope. Microphotographs were taken using a Cool Snap digital camera fitted with QCapture Pro 6.0 photographic software. Wood anatomical characters were measured and described according to the IAWA list of microscopic features for softwood identification53. The systematic position of the fossil wood was determined by consulting the key to morphogenera of fossil conifer woods54 and by carefully comparing with similar modern and fossil woods. Fossil wood generic nomenclature and circumscription followed the criteria of Philippe & Bamford54. Fossil wood specimens and thin sections are housed in the Museum of Biology, Sun Yat-sen University, Guangzhou, China.
Additional Information
How to cite this article: Li, L. et al. First record of Podocarpoid fossil wood in South China. Sci. Rep. 6, 32294; doi: 10.1038/srep32294 (2016).
References
Farjon, A. World checklist and bibliography of conifers. Richmond: Royal Botanic Gardens, Kew, 2 edition (2001).
Conran, J. G. et al. Generic relationships within and between the gymnosperm families and Phyllocladaceae based on an analysis of the chloroplast gene rbcL. Aust. J. Bot. 48, 715–724 (2000).
Dalling, J. W. et al. “Ecology and Distribution of Neotropical Podocarpaceae” In Ecology of the Podocarpaceae in tropical forests, eds Turner, B. L. & Cernusak, L. A. (Washington, DC: : Smithsonian Insitiution Scholarly Press,) 95, 43–56 (2011).
Enright, N. J. & Jaffré, T. “Ecology and Distribution of the Malesian Podocarps” In Ecology of the Podocarpaceae in tropical forests. eds Turner, B. L. & Cernusak, L. A. (Washington, DC: : Smithsonian Insitiution Scholarly Press,) 95, 57–77 (2011).
Adie, H. & Lawes, M. J. “Podocarps in Africa: Temperate Zone Relicts or Rainforest Survivors?” In Ecology of the Podocarpaceae in Tropical Forests. eds Turner, B. L. & Cernusak, L. A. (Washington, DC: : Smithsonian Insitiution Scholarly Press,) 95, 79–100 (2011).
Cernusak, L. A. et al. “Podocarpaceae in Tropical Forests: A Synthesis” In Ecology of the Podocarpaceae in Tropical Forests. eds Turner, B. L. & Cernusak, L. A. (Washington, D.C.: : Smithsonian Insitiution Scholarly Press,), 189–196 (2011).
Morley, R. J. “Dispersal and Paleoecology of Tropical Podocarps” In Ecology of the Podocarpaceae in Tropical Forests, eds Turner, B. L. & Cernusak, L. A. (Washington, D.C.: : Smithsonian Insitiution Scholarly Press,) 21–42 (2011).
Biffin, E., Conran, J. G. & Lowe, A. J. “Podocarp Evolution: a Molecular Phylogenetic Perspective” In Ecology of the Podocarpaceae in tropical forests. eds Turner, B. L. & Cernusak, L. A. (Washington, D.C.: : Smithsonian Insitiution Scholarly Press,) 1–20 (2011).
Dilcher, D. Podocarpus from the Eocene of North America. Science 164, 299–301 (1969).
Taggart, R. E. Additions to the Miocene Sucker Creek flora of Oregon and Idaho. Amer. J. Bot. 60, 923–928 (1973).
Leopold, E. B., Manchester, S. R. & Meyer, H. W. “Phytogeography of the Late Eocene Florissant flora reconsidered” In Paleontology of the Upper Eocene Florissant Formation, Colorado. Geological Society of America Special Paper 435. eds Meyer, H. W. & Smith, D. M. (Boulder, Colorado: Geological Society of America) 53–70 (2008).
Greenwood, D. R., Hill, C. R. & Conran, J. G. Prumnopitys anglica sp. nov. (Podocarpaceae) from the Eocene of England. Taxon 62, 565–580 (2013).
Krasnov, A. N. Primitiae florae tertiariae Rossiae meridionalis. Pechatnik, Kharkov. 109 p. (in Russian) (1910).
Pimenova, N. V. Flora of the Tertiary sandstones of the western bank of Dnepier river in the Ukrainian S.S.R. Acad. Sci. Ukr. S.S.R., Kiev. 176 pp. (in Ukrainian) (1937).
Makulbekov, N. M. Paleogene floras of the Western Kazakhstan and the lower reaches of the Volga river. Alma-Ata: Nauka (Kazakhstan SSR). 234 p. (in Russian) (1977).
Ferguson, D. K., Jähnichen, H. & Alvin, K. L. Amentotaxus Pilger from the European Tertiary. Feddes Repert. 89, 379–410 (1978).
Chudajberdyev, R. Fossil woods from the outskirts of Smolino lake. Botanicheskij Zhurnal 18(5), 704–710 (in Russian) (1958).
Chiguriaeva, A. A. Atlas of microspores from the Tertiary deposits of the USSR. Kharkov: A.M. Gorky Kharkov State University. 118 pp. (in Russian) (1956).
Ferguson, D. K. On the phytogeography of Coniferales in the European Cenozoic. Palaeogeogr. Palaeoclimatol. Palaeoecol. 3, 73–110 (1967).
Cavagnetto, C. & Anadón, P. Preliminary palynological data on floristic and climatic changes during the Middle Eocene–Early Oligocene of the eastern Ebro Basin, northeast Spain. Rev. Palaeobot. Palynol. 92, 281–305 (1996).
Pflug, H. Sporenbilder aus Island und ihre stratigraphische Deutung. Neues Jahrb. Geol. Paläontol. 107(2), 141–172 (1959).
Greguss, P. & Vanhoorne, R. L'Âge des couches de sable situées entres les argiles de la Campine et le Poederlien en Belgique. Bull. Inst. Roy. Sci. Nat. Belgique 40(3), 1–6 (1964).
Srivastava, R. & Guleria, J. S. A Catalogue of Cenozoic (Tertiary) plant megafossils from India (1989–1005). Birbal Sahni Insititute of Palaeobotany: Diamond Jubilee Special Publication. Lucknow. P. 1–76 (2006).
Awasthi, N., Mehrotra, R. C. & Lakhanpal, R. N. Occurrence of Podocarpus and Mesua in the Oligocene sediments of Makum Coalfield, Assam, India. Geophytology 22, 193–198 (1992).
Jin, J. H., Qiu, J., Zhu, Y. A. & Kodrul, T. M. First fossil record of the genus Nageia (Podocarpaceae) in south China and its phytogeographic implications. Pl. Syst. Evol. 285, 159–163 (2010).
Liu, X. Y., Gao, Q. & Jin, J. H. Late Eocene leaves of Nageia Gaertner (section Demmaroideae Mill.) from Maoming Basin, South China and their implications on phytogeography. Journal of Systematics and Evolution 53(4), 297–307 (2015).
Yang, J. J., Qi, G. F. & Xu, R. H. Studies on fossil woods excavated from the Dabie mountains. Sci. Silvae Sin. 26(4), 379–386 (1990).
Cui, J. Studies on the Fusinized-Wood Fossils of Podocarpaceae From Huolinhe Coalfield, Inner Mongolia, China. Acta Botanica Sinica 37(8), 636–640 (1995).
He, D. C. The coal forming plants of Late Mesozoic in Da Hinggan Mountains. China Coal Industry Publishing House, Beijing (in Chinese with English abstract) (1995).
Zaklinskaia, E. O. Stratigraphic significance of the gymnosperm pollen from Cenozoic deposits of the Pavlodar region of Irtysh basin and of the northern Aral Sea region. Moscow: Academy of Sciences of the USSR. 221 p. (in Russian) (1957).
Phillips, E. W. J. The identification of coniferous woods by their microscopic structure. Journal of the Linnean Society of London, Botany 52, 259–320 (1941).
Greguss, P. Identification of gymnosperms on the basis of xylotomy. Budapest, Akadémiai Kiadó. 263 p (1955).
Chavchavadze, E. S. Wood of conifers. Leningrad: Nauka Publishing House. 192 p. (in Russian) (1979).
Jiang, X. M., Cheng, Y. M. & Yin, Y. F. Atlas of Gymnosperms Woods of China. Beijing: Science Press, 1–490 (2010).
Afonin, M. & Philippe, M. Fossil woods from the Lower Cretaceous (Albian) of Kamchatka Peninsula. Russian Far East. Cretaceous Research 50, 110–119 (2014).
Patel, N. R. Wood anatomy of podocarpaceae indigenous to New Zealand. 2. Podocarpus. New Zealand Journal of Botany 5, 307–321 (1967b).
Patel, N. R. Wood anatomy of Podocarpaceae indigenous to New Zealand. 1. Dacrydium New Zealand Journal of Botany 5, 171–184 (1967a).
Patel, N. R. Wood anatomy of Podocarpaceae indigenous to New Zealand. 3. Phyllocladus. New Zealand Journal of Botany 6, 3–8 (1968).
Esteban, L. G., de Palacios, P. D. P., Casasús, A. J. & Fernandez, F. G. Characterisation of the xylem of 352 conifers. Investigación Agraria, Sistemas y Recursos Forestales 13, 452–478 (2004).
Vásquez Correa, M. A., Alcántara Vara, E. & Herrera Machuca, M. A. Wood anatomy of Colombian Podocarpaceae (Podocarpus, Prumnopitys and Retrophyllum). Botanical Journal of the Linnean Society 164, 293–302 (2010).
Süss, H. & Velitzelos, E. Zwei neue fossile Hölzer der Formgattung Podocarpoxylon Gothan aus tertiären Schichten der Insel Lesbos, Griechenland. Feddes Repertorium 111(3–4), 135–149 (2000).
Franco, M. J. & Brea, M. First extra-Patagonian record of Podocarpaceae fossil wood in the Upper Cenozoic (Ituzaingó Formation) of Argentina. New Zealand Journal of Botany 53(2), 103–116 (2015).
Patton, R. T. Fossil wood from Victorian brown coal. Proceedings of the Royal Society of Victoria 70, 129–143 (1958).
Gothan, W. Die fossilen Hölzer von der Seymour und Snow Hill Insel. Wissenschaftliche Ergebnisse der Schwedischen Südpolar Expedition 1901–1903. 3, Stockholm, Lithographisches Institut des Generalstabs, 1–33 (1908).
Pujana, R. R., Santillana, S. N. & Marenssi, S. A. Conifer fossil woods from the La Meseta Formation (Eocene of Western Antarctica): Evidence of Podocarpaceae-dominated forests. Rev. Palaeobot. Palynol. 200, 122–137 (2014).
Creber, G. T. & Chaloner, W. G. Influence of environmental factors on the wood structure of living and fossil trees. Botanical Review 50, 357–448 (1984).
Brison, A., Philippe, M. & Thévenard, F. Are Mesozoic growth rings climate induced? Paleobiology 27, 531–538 (2001).
Taylor, E. L. & Ryberg, P. E. Tree growth at polar latitudes based on fossil growth ring analysis. Palaeogeography, Palaeoclimatology, Palaeoecology. 255, 246–264 (2007).
Farjon, A. & Filer, D. An Atlas of the World’s Conifers. Leiden: Brill. 512 p. (2013).
Peel, M. C., Finlayson, B. L. & McMahon, T. A. Updated world map of the Köppen-Geiger climate classification. Hydrology and Earth System Sciences 11, 1633–1644 (2007).
Tong, Y. S. A review of middle to late Eocene Mammalian faunas from China. Acta Palaeontologica Sinica 28, 663–682 (1989).
Lacey, W. S. “Palaeobotany technique, 2” In Viewpoint in Biology, eds Carthey, J. D. & Duddington, C. L. (Butterworths, London) 202–243 (1963).
IAWA Committee IAWA List of microscopic features for softwood identification. IAWA Journal 25, 1–70 (2004).
Philippe, M. & Bamford, M. K. A key to morphogenera used for Mesozoic conifer-like woods. Rev. Palaeobot. Palynol. 148, 184–207 (2008).
Kräusel, R. Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wusten Ägyptens. IV. Die fossilen Floren Ägyptens. Abhandlungen der Bayerischen. Akademie Wissenschaften. Mathematisch-naturwissenschaftliche Abteilung. Neue Folge.Heft 47, 1–140 (1939).
Schönfeld, E. Die Kieselhölzer aus der Braunkohle von Böhlen bei Leipzig. Palaeontographica Abteilung B 99, 1–83 (1955).
Kräusel, R. Die fossilen Konifern-Hölzer (unter Ausschluss von Araucarioxylon Kraus) II. Kritische Untersuchungen zur Diagnostik lebender und fossiler Koniferen-Hölzer. Palaeontogr. Bohem. 89, 83–203 (1949).
Stockmans, F. Végétaux éocènes des environs de Bruxelles. Mtm. Mus. Roy. Hist. natur. Belg. 76, 3–56 (1936).
Zalewska, Z. Tertiary remains of fossil wood from Turów on the Lusatian Neisse I. Acta geol. Polon. 3, 481–543 (in Polish) (1953)
Kräusel, R. Beitrage zur Kenntnis der fossilen Flora von Südamerika. I. Fossile Hölzer aus Patagonien und benachbarten Gebieten. Arkiv för Botanik 19(9), 1–36 (1924)
Blokhina, N. I. “Fossil woods” In Cretaceous-Palaeogene of the Lesser Kuril Islands. New data on papaeontology and geological history. eds Krasilov, V. A., Blokhina, N. I., Markevich, V. S. & Ja, M. Serova (DVO AN SSSR: Vladivostok), 26–40 (in Russian) (1988).
Shilkina, I. A. The fossil woods of the Goderdzy Pass. Proceedings of the Komarov Botanical Institute. Ser. 8. Paleobotany 3, 127–179 (in Russian) (1958).
Rössler, W. Pliozäne Koniferenhölzer der Umgebung von Gleichenberg in Steiermark. Mitt. naturwiss. Ver. Steiermark 74, 64–97 (1937).
Lakhanpal, R. N., Guleria, J. S. & Awasthi, N. A podocarpaceous wood from the Pliocene of Kutch. Geophytology 5, 172–177 (1975).
Greenwood, D. R. Miocene wood from La Trobe Valley coal measures, Victoria, Australia. Alcheringa 29, 351–363 (2005).
Trivedi, B. S. & Srivastava, R. Gymnospermous woods from Early Tertiary of Chhindwara District of Madhya Pradesh. Phytomorphology 39(1), 61–68 (1989).
Müller-Stoll, W. R. & Schultze-Motel, I. Gymnospermen-Hölzer des Deutschen Jura. Teil 3: Abietoid (modem) getüpfelte Hölzer. Z. deutsch. geol. Ges. 141, 61–77 (1990).
Sahni, B. Revisions of Indian fossil plants. Part II. Coniferales (b. Petrifications). Mem. Geol. Surv. India (Palaeont. Ind.), N. S. 11, 1–124 (1931).
Bande, M. & Prakash, U. A podocarpaceaous fossil wood from the Deccan Intertrappean Beds of Malabar Hills, Bombay. Geophytology 14, 171–177 (1984).
Lemoigne, Y. & Beauchamp, J. P. Paléoflores tertiaires de la région de Welkite (Ethiopie, province du Shoa). Bull. Soc. Géol. Fr. 14, 336–346 (1972).
Adamson, S. & Currin, M. M. Buried trees in the Cape flats. Trans. Roy. Soc. South Africa 33, 13–23 (1951).
Wijninga, V. M. A Pliocene Podocarpus forest mire from the area of the high plain of Bogotá (Cordillera Oriental, Colombia). Rev. Palaeobot. Palynol. 92, 157–205 (1996).
Acknowledgements
This study was supported by the National Natural Science Foundation of China (Grant nos 41572011, 41528201, 41210001, 41372002), the joint Project of the National Natural Science Foundation of China and the Russian Foundation for Basic Research (Grant nos 41611130044, 16-55-53007), the Fundamental Research Funds for the Central Universities (Grant no. 16lgjc28) and the Scientific Research Fund, Hongda Zhang, Sun Yat-sen University. We thank the University of Johannesburg and the Komarov Botanical Institute (institutional research project no. 01201456545) for financial support for A.A.O. We thank graduate students majoring in Plant Science at Sun Yat-sen University for participating in the field collection of the fossils. We also thank Dr. Xin-Xin Feng for his helpful suggestions on this study and Yang Zeng, librarian of Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences for references. We also offer our sincere gratitude to Prof. R.A. Spicer for editing.
Author information
Authors and Affiliations
Contributions
C.Q., J.-H.J. and L.L. collected the fossil wood from Donghuai. J.-H.J., C.Q. and L.L. participated in the design of the study. L.L. and A.A.O. photographed specimens and arranged figures. L.L. and A.A.O. carried out the data analyses. J.-H.J., L.L. and A.A.O. conducted taxonomic treatments, phytogeographic and ecological interpretation. L.L. and A.A.O. wrote the manuscript and formatted the text. All authors read and approved the final manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Li, L., Jin, JH., Quan, C. et al. First record of Podocarpoid fossil wood in South China. Sci Rep 6, 32294 (2016). https://doi.org/10.1038/srep32294
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep32294
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
