
Abstract
Qinghai Lake is a major migrating bird breeding site that has experienced several recent highly pathogenic avian influenza virus (HPAIV) epizootics. From 2006 to 2009 we studied Qinghai’s wild birds and pikas for evidence of AIV infections. We sampled 941 healthy wild animals and isolated seventeen H7N2 viruses (eight from pikas and nine from wild birds). The H7N2 viruses were phylogenetically closely related to each other and to viruses isolated in Hong Kong in the 1970s. We determined the pathogenicity of the H7N2 viruses by infecting chickens and mice. Our results suggest that pikas might play an important role in the ecology of AIVs, acting as intermediate hosts in which viruses become more adapted to mammals. Our findings of AI infection in pikas are consistent with previous observations and raise the possibility that pikas might play a previously unrecognized role in the ecology of AIVs peridomestic aquatic environments.
Similar content being viewed by others


Introduction
Influenza A viruses (IAVs) exhibit a wide host range that includes birds and mammals. Viruses exhibit varying degrees of host adaptation after crossing species barriers1,2. Influenza A viruses pose a continual threat to humans and animals because they frequently change via mutation and/or reassortment.
Wild aquatic birds are the main natural reservoir of IAVs, harboring 16 of 18 hemagglutinin (HA) and 9 of 11 recognized neuraminidase (NAs) glycoproteins3,4,5,6. IAVs replicate in the gastrointestinal track of aquatic birds and are most often spread through the fecal-orally route2. They generally cause little or no signs of disease in birds. Migratory birds have a the potential to spread these viruses over large geographical areas.
Qinghai Lake, the largest lake in China is located in the Qinghai-Tibet plateau and serves as a home for ~150,000 birds each year. It constitutes major stop area for various migration flyways. Notably, after highly pathogenic avian influenza (HPAI) H5N1 outbreaks occurred among wild birds at Qinghai Lake in 2005 and 20067,8, HPAI H5N1 strains spread across major flyways to North America, Eurasia, and Africa9 suggesting a geotemporal association.
The pika (Ochotona curzoniae), a small rabbit-like mammal, is a natural resident of the Qinghai-Tibet plateau that has recently been considered as a natural host for influenza viruses10,11. Sialic acids with affinity for human and avian IAVs are expressed in the respiratory tract of pikas12. Previously, HPAI H5N1 viruses have been isolated from pikas10 and pikas have been shown to be seropositive against H9N2 low pathogenic avian influenza (LPAI) viruses11. Additionally, experimental infections have shown that pikas are susceptible to HPAI H5N1, LPAI H9N2, human H1N1, and human H3N2 viruses12. These data suggest that pikas could play an important role in influenza ecology, and thus might deserve more attention in surveillance studies.
In the past, surveillance for influenza viruses in Qinghai Province focused mainly on aquatic birds and poultry7,8,13,14,15,16,17 and wild mammals have seldom been studied. With this knowledge gap in mind, we conducted this study of wild birds and pikas.
Materials and Methods
Sample collection
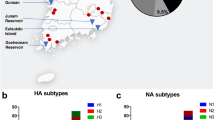
In an effort to evaluate wild animals in the Qinghai Province ecosystem, we captured wild birds and black-lipped pikas (Ochotona curzoniae) from August 2006 to October 2009 in Qinghai Province some captured site where are on the distributions of generalized migration routes of migratory birds18). Specific sites included most areas around the Qinghai lake: Qumalai, Zhiduo, Jiuzhi, Maduo, Golmud, Delingha, Wulan, Gangsha, Gonghe, Tongren, Qilian, Mengyuan and Yushu (Fig. 1). All captured animals were clinically healthy. Aquatic and passerine birds were swabbed and released. Pikas were euthanized and their tissues (heart, liver, spleen, lungs, kidneys, and brain) were harvested and preserved in liquid nitrogen.

The circles indicate dates and locations of animal-trapping sites. The picture is created by Photoshop CS5.
Virus isolation and identification
Virus isolation in 10-day-old specific pathogen free (SPF) embryonated chicken eggs was attempted from each tissue sample. Virus subtyping was performed by RT-PCR19.
Genome sequencing and phylogenetic tree analysis
Viruses were sequenced as previously described10. Briefly, total RNA was extracted from allantoic fluid with Trizol® LS (Invitrogen, Carlsbad, CA), followed by RT-PCR using a One-Step RT-PCR kit (Qiagen, Hilden, Germany) with universal influenza primers20. PCR products were cloned into the pMD18-T vector, and sequenced. Viral sequences were aligned using Clustal W within the BioEdit software package (version 5.0.9). Maximum likelihood phylogenetic trees were inferred with 1000 bootstraps using MEGA (Version 6).
Animal infections
We characterized the in vivo pathogenicity of two AIVs isolated from pikas: A/Pika/QH-Maduo/01/2006 (Maduo06) and A/Pika/Maduo/01/2009(Maduo09). Ten 5-week-old SPF white leghorn chickens (obtained from the Merial Vital Laboratory Animal Technology Co. Ltd., Beijing, China) were housed in negative-pressure isolator cages with HEPA-filtered air. They were inoculated intravenously (i.v.) with 0.2 ml/chicken of a 1:10 dilution of bacteria-free fresh allantoic fluid containing a virus isolate. The process was performed according to OIE methods21 in evaluating the intravenous pathogenicity index (IVPI).
In addition, ten chickens were inoculated intranasally with 106.0EID50/animal of each the viruses under study in a 0.2 ml volume, and 5 chickens were inoculated intranasally with 0.2 ml PBS as non-infection control. To evaluate bird to bird transmission, two additional chickens were housed with the inoculated chickens at 12 hours post infection. Clinical symptoms were recorded every day for 14 days. Oropharyngeal and cloacal swabs were collected from animals on days three, five, and seven post-inoculation (pi.) to evaluate viral shedding. Three chickens from each group were euthanized on day three pi to measure virus replication in lungs, intestine, pancreas, heart, spleen, kidney, thymus, and brain. Viral titers (EID50) were calculated using the method of Reed and Muench22.
Six-week-old SPF BALB/c mice obtained from the Experiment Animal Center of Zhejiang University, Hangzhou, China, were randomly divided into four groups (20/group). Mice were anesthetized with dry ice, and inoculated intranasally (i.n.) with 106.0EID50 of Maduo06 and Maduo09 viruses in 50 μl PBS, respectively. As non-infection control, the other group (20 mice) were inoculated i.n. with 50 μl PBS. Mice were monitored daily for weight loss and mortality. Three mice in each group were euthanized on days three, five and seven pi to evaluate virus replication and histopathological changes in lung, intestine, pancreas, heart, spleen, kidney, thymus, and brain. The viral titers (EID50) were calculated using the method of Reed and Muench22.
Ethics statement
Animal experiments were executed in accordance with the Regulations for the Administration of Affairs Concerning Experimental Animals approved by the State Council of the People’s Republic of China. All the animal experiments and samples collected were approved by the Institutional Animal Care and Use Committee (IACUC) of Zhejiang University, permission number: SYXK 2012-0178.
Results
Influenza isolation from the wild animals in Qinghai
From August 2006 to October 2009, 360 pikas, 74 resident wild birds, 14 migrating birds and 493 water birds were captured from thirteen sites of the Qinghai-Tibet plateau and a total of 22 viruses (2.3%) were isolated. HA and NA sequencing revealed that seventeen viruses were of the H7N2 subtype and five viruses were of the H5N1 type (H5N1 viruses were published before). For the H7N2 viruses (Table 1), two viruses were isolated from migrating birds (Acanthis), and seven viruses were isolated from resident birds (Passer montanus, Melanocorypha mongolica, Podoces and Montifringilla), and eight viruses were isolated from pikas.
Molecular characterization and phylogenetic analysis of the H7N2 influenza viruses
Phylogenetic analysis showed that the HA genes of all H7 subtype isolates were divided into two geographical distinct lineages (Eurasian and American) (Fig. 2). These individual groups were classified by temporal and geographic relationships rather than by host species. All H7N2 isolates were closely related to 1970s-era Hong Kong AI H7 isolates and clustered in a group, which belongs to the Eurasian branch (Fig. 2). These seventeen H7N2 viruses were very different by HA gene analyses from H7N9 viruses that have infected humans in China since 2013 (Fig. 2). Phylogenetic analysis of the NA and internal genes also showed clustering with H7 AIVs isolated in Hong Kong (Figure S1), suggesting that the H7N2 viruses isolated from pikas were of avian origin. We did not detect reassortment in any of H7N2 viruses.

Viruses isolated in this study are indicated in red. Maximum likelihood phylogenetic trees were inferred and with 1000 times bootstrap using MEGA (Version 7).
We further examined the isolates for the presence of putative mammalian adaptive mutations and antiviral resistance (Table 2). The HA cleavage site of all H7N2 isolates is PEIPKGR, and the receptor binding sites in HA exhibited Q and G in positions 222 and 224, respectively (H3 numbering). No NA deletion was observed in any of the H7N2 isolates. Positions 627 and 701 in PB2 displayed E and D, respectively. Mutations in NA (Oseltamivir resistant amino acid sites: 119E, 274H and 292R) and M2 (Amantadine resistant amino acid sites: 26L, 27V, 30A and 31S) associated with antiviral resistance were not detected.
Animal infections
Maduo06 and Maduo09 viruses were used to assess the pathogenicity of H7N2 viruses moving from pikas to chickens and mice. Results from pathogenicity studies indicated that the IVPI values in chickens were 0.17 (Maduo06) and 0.19 (Maduo09). Contact chickens showed no clinical symptoms, and the infected chickens showed unapparent clinical symptoms except for transient depression. High viral replication was only detected in lung tissues with a mean viral load titer of 2.65 ± 0.15 (log10EID50/g) for Maduo06 and 2.15 ± 0.08 (log10EID50/g) for Maduo09 (Fig. 3A). Low viral replication was detected in the intestines, with mean titers of 0.25 ± 0.00 (log10EID50/g) for Maduo06 and 0.75 ± 0.00 (log10EID50/g) for Maduo09 (Fig. 3A). Virus replication was not detected in other organs. Viruses were detected from the oropharynx and cloaca of infected chickens with maximum viral shedding occurring at seven dpi. Virus titers were between 1.25 ± 0.00 to 3.44 ± 0.58 (log10EID50/ml) (Table 3).

Viral replication in tissues of inoculated chickens (A) and the body weight of inoculation mice (B). Maduo06: A/Pika/QH-Maduo/01/2006(H7N2); Maduo09: A/Pika/Maduo/01/2009(H7N2).
Mice survived the experimental infections with H7N2 viruses. Body weight increased over time and there were no differences when compared to the control group (Fig. 3B). No viral replication was detected in any of the organs of the euthanized mice. Histological examination revealed no pathological changes in animals infected with either virus. These data confirmed that the H7N2 viruses isolated from pikas were of low pathogenicity in chickens and mice.
Discussion
Previous reports have suggested that plateau pikas can be asymptomatically infected with HPAI H5N110 and LPAI H9N211. The findings raise the possibility that pikas might play a role in the transmission of AIVs at Qinghai-Tibet Plateau and provide opportunities for the adaptation of AIVs to mammals. Our results provide evidence that H7N2 AIV has become enzootic in wild animals around Qinghai Lake. We isolated five H5N110 and eight H7N2 viruses from pikas. Additionally, nine H7N2 viruses were isolated from wild birds. Overall, our findings suggest that both H5N1 and H7N2 viruses continue to circulate in the Qinghai-Tibet Plateau area ecosystem.
Our study validates a previous report suggesting that the wild pikas could serve as a previously unrecognized natural host of IAVs. Like pigs, pikas possess both avian and mammalian receptors and could potentially serve as “mixing vessels” for the generation of novel IAVs12. Of note, pikas have been experimentally infected with both human H1N1 and human H3N2 influenza viruses12. Their susceptibility to various influenza A viruses adds to the complex ecosystem around the Qinghai Lake in Qinghai-Tibet Plateau area. As species of wild birds may live in close contact with pikas in a den, we speculate that complex viral transmission patterns involving multiple species might commonly occur in this ecosystem.
From the animal experiments, we found that H7N2 isolates were low pathogenic to chickens and mice. In our field investigations, all samples were obtained from healthy-appearing wild animals. Under experimental conditions, the pika-associated H5N1viruses (2.2 clade and 2.3.2 clade) resulted in sub-clinical infection in rabbits (a species closely related to pikas)10. Hence, it seems possible that pikas may serve as healthy mammalian reservoir for IAVs. What is not clear is how to implement interventions that would reduce mammal contact with migrating aquatic birds or humans. The H5N1/H7N2/H9N2 AIVs infections in pikas detected here and past experiments suggests that they should be considered in future AIV surveillance programs at least for the Qinghai-Tibet Plateau area.
As mentioned above, Qinghai Lake is a major breeding site for migratory birds that overwinter from Southeast Asia, India, and Tibet and the migratory range of these birds is significant. In 2005, a HPAIV H5N1 outbreak occurred among migratory wild birds at Qinghai Lake7 threatening nearby poultry and mammalian species17. This H5N1 lineage subsequently spread throughout North America, Eurasia, and Africa9. This threat is likely to continue as multiple HPAI H5N1 lineages have been persistently circulating in Qinghai8,14,15,16 and H9N2 AIVs have also infected wild birds at Qinghai Lake13,23.
Asymptomatic infection of plateau pikas with H5, H7 and H9 subtype AIVs could lead to mammalian adaptation. Fortunately, in our study, the HA and NA genes of the H7N2 isolates had no evidence of recent reassortment, suggesting that these viruses have become enzootic and stable in these wild animals. It is unclear why the Qinghai H7N2 viruses which have the ability to infect both wild birds and mammals have not yet been detected in domestic poultry in the Qinghai-Tibet plateau area. Perhaps this is due to limited AIV surveillance among poultry in this area. It seems logical that if AIVs are enzootic among the many wild birds in the Qinghai-Tibet plateau area, especially around the Qinghai Lake area, infections in other wild and domestic animals are likely occurring but unrecognized. More extensive AIV surveillance among Qinghai Lakes migrating birds, wild mammals, and domestic animals seems imperative.
Our study highlights once again the importance of Qinghai-Tibet Plateau area ecosystem in influenza virus circulation. The finding of H7N2 subtype influenza viruses from wild birds and pikas expands the complexity of the influenza virus gene pool in Qinghai Province.
Additional Information
How to cite this article: Su, S. et al. Characterization of H7N2 Avian Influenza Virus in Wild Birds and Pikas in Qinghai-Tibet Plateau Area. Sci. Rep. 6, 30974; doi: 10.1038/srep30974 (2016).
References
Webster, R. G., Bean, W. J., Gorman, O. T., Chambers, T. M. & Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiological reviews 56, 152–179 (1992).
Yoon, S. W., Webby, R. J. & Webster, R. G. Evolution and ecology of influenza A viruses. Current topics in microbiology and immunology 385, 359–375, doi: 10.1007/82_2014_396 (2014).
Easterday, B. C., Trainer, D. O., Tumova, B. & Pereira, H. G. Evidence of infection with influenza viruses in migratory waterfowl. Nature 219, 523–524 (1968).
Munster, V. J. et al. Towards improved influenza A virus surveillance in migrating birds. Vaccine 24, 6729–6733, doi: 10.1016/j.vaccine.2006.05.060 (2006).
Olsen, B. et al. Global patterns of influenza a virus in wild birds. Science 312, 384–388, doi: 10.1126/science.1122438 (2006).
Webby, R. J., Webster, R. G. & Richt, J. A. Influenza viruses in animal wildlife populations. Current topics in microbiology and immunology 315, 67–83 (2007).
Liu, J. et al. Highly pathogenic H5N1 influenza virus infection in migratory birds. Science 309, 1206, doi: 10.1126/science.1115273 (2005).
Wang, G. et al. H5N1 avian influenza re-emergence of Lake Qinghai: phylogenetic and antigenic analyses of the newly isolated viruses and roles of migratory birds in virus circulation. The Journal of general virology 89, 697–702, doi: 10.1099/vir.0.83419-0 (2008).
Cui, P. et al. Bird migration and risk for H5N1 transmission into Qinghai Lake, China. Vector borne and zoonotic diseases 11, 567–576, doi: 10.1089/vbz.2009.0240 (2011).
Zhou, J. et al. Characterization of the H5N1 highly pathogenic avian influenza virus derived from wild pikas in China. Journal of virology 83, 8957–8964, doi: 10.1128/JVI.00793-09 (2009).
Yu, Z. et al. Lowly pathogenic avian influenza (H9N2) infection in Plateau pika (Ochotona curzoniae), Qinghai Lake, China. Veterinary microbiology 173, 132–135, doi: 10.1016/j.vetmic.2014.07.002 (2014).
Li, Y. et al. Distribution of sialic acid receptors and experimental infections with different subtypes of influenza A viruses in Qinghai-Tibet plateau wild pika. Virology journal 12, 63, doi: 10.1186/s12985-015-0290-8 (2015).
Liu, L. Y. et al. [Complete genome phylogenetic analysis of five H9N2 avian influenza viruses isolated from poultry flocks in Qinghai lake region]. Bing du xue bao = Chinese journal of virology/[bian ji, Bing du xue bao bian ji wei yuan hui] 30, 109–118 (2014).
Li, Y. et al. New avian influenza virus (H5N1) in wild birds, Qinghai, China. Emerging infectious diseases 17, 265–267, doi: 10.3201/eid1702.100732 (2011).
Li, Y. et al. Persistent circulation of highly pathogenic influenza H5N1 virus in Lake Qinghai area of China. Avian diseases 54, 821–829, doi: 10.1637/8657-021309-Reg.1 (2010).
Hu, X. et al. Clade 2.3.2 avian influenza virus (H5N1), Qinghai Lake region, China, 2009-2010. Emerging infectious diseases 17, 560–562, doi: 10.3201/eid1703.100948 (2011).
Zhou, J. Y. et al. Characterization of a highly pathogenic H5N1 influenza virus derived from bar-headed geese in China. The Journal of general virology 87, 1823–1833, doi: 10.1099/vir.0.81800-0 (2006).
Fang, L. Q. et al. Environmental factors contributing to the spread of H5N1 avian influenza in mainland China. PloS one 3, e2268, doi: 10.1371/journal.pone.0002268 (2008).
Lee, M. S., Chang, P. C., Shien, J. H., Cheng, M. C. & Shieh, H. K. Identification and subtyping of avian influenza viruses by reverse transcription-PCR. Journal of virological methods 97, 13–22 (2001).
Gall, A., Hoffmann, B., Harder, T., Grund, C. & Beer, M. Universal primer set for amplification and sequencing of HA(0) cleavage sites of all influenza A viruses. J Clin Microbiol 46, 2561–2567, doi: 10.1128/Jcm.00466-08 (2008).
Shankar, B. et al. Assessment of pathogenic potential of two Indian H5N1 highly pathogenic avian influenza virus isolates by intravenous pathogenicity index test. International Journal of Poultry Science 8, 283–290 (2009).
Reed, L. J. & Muench, H. A simple method of estimating fifty per cent endpoints. American journal of epidemiology 27, 493–497 (1938).
Zhang, H., Li, Y., Chen, J., Chen, Q. & Chen, Z. Perpetuation of H5N1 and H9N2 avian influenza viruses in natural water bodies. The Journal of general virology 95, 1430–1435, doi: 10.1099/vir.0.063438-0 (2014).
Acknowledgements
This paper was supported in part by National Program on key research project of China (2016YFC1201604), National Key Technology R & D Program of China (Grant No. 2015BAD12B00), the Fundamental Research Funds for the Central Universities (Y0201600147), China Agriculture Research System (Grant No. CARS-41-K11), the Wildlife Department of the State Forestry Administration and the Priority Academic Program Development of Jiangsu Higher Education Institutions. We thank Dr. Pablo Murcia critically reading the manuscript.
Author information
Authors and Affiliations
Contributions
S.S. conceived the study and wrote the paper. J.W., Y.Y., Z.L., J.G., L.Y., B.H., G.X., J.L. and S.J. performed and analyzed all the experiments. S.S., G.C.G. and J.Z. designed the study and revised the manuscript. All authors reviewed the results and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Su, S., Xing, G., Wang, J. et al. Characterization of H7N2 Avian Influenza Virus in Wild Birds and Pikas in Qinghai-Tibet Plateau Area. Sci Rep 6, 30974 (2016). https://doi.org/10.1038/srep30974
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep30974
This article is cited by
-
TLR7 and TLR8 evolution in lagomorphs: different patterns in the different lineages
Immunogenetics (2022)
-
Cottontail rabbits shed clade 2.3.4.4 H5 highly pathogenic avian influenza A viruses
Archives of Virology (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
