
Abstract
Bacterial transporters mediate the exchanges between intracellular and extracellular environments. Modification of transport route could be applied to speed up the metabolic reactions and promote the production of aimed compounds. Herein, lysine 2-monooxygenase (DavB) and δ-aminovaleramidase (DavA) were co-expressed in Escherichia coli BL21(DE3) to produce nylon-5 monomer 5-aminovalerate from l-lysine. Then, PP2911 (4-aminobutyrate transporter in Pseudomonas putida) and LysP (the lysine specific permease in E. coli) were overexpressed to promote 5-aminovalerate production using whole cells of recombinant E. coli. The constructed E. coli strain overexpressing transport proteins exhibited good 5-aminovalerate production performance and might serve as a promising biocatalyst for 5-aminovalerate production from l-lysine. This strategy not only shows an efficient process for the production of nylon monomers but also might be used in production of other chemicals.
Similar content being viewed by others

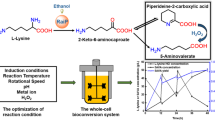

Introduction
Polyamides (also known as nylons), owing to their excellent material properties in mechanical strength, melting point, corrosion resistance and other superior qualities, have been applied extensively in industrial and medical fields1,2,3. The basic materials for polyamides production are generally acquired from petrochemicals. As concerns about environmental issues and petroleum resource availability continue to grow, it is critical that we produce viable alternatives from cost-effective renewable resources. Nylon-4,6 has been commercialized and outpaces nylon-6 and nylon-6,6 in melting and thermal stability4. Nylon-5, which is produced from the homopolymerization of 5-amniovalerate, exhibits similar general properties to nylon-4,64. What is significant is that 5-amniovalerate could be produced through biotechnological routes, which guarantees the green route to nylon-5 production. 5-Aminovalerate could also serve as a potential C5 platform chemical to produce 5-hydroxyvalerate, glutarate and 1,5-pentanediol5. In this regard, it is of high importance to develop methods for bio-synthesis of 5-aminovalerate.
5-Aminovalerate is an intermediate of l-lysine catabolism that is called aminovalerate pathway in Pseudomonas putida6. The transformation process is composed of two steps: oxidation of l-lysine into 5-aminovaleramide catalyzed by lysine 2-monooxygenase (DavB) and hydrolysis of 5-aminovaleramide into 5-aminovalerate catalyzed by δ-aminovaleramidase (DavA). Great efforts have been made for fermentative production of l-lysine7,8,9,10,11,12,13,14,15,16,17. The annual production of l-lysine is estimated at a large quantity of 2,200,000 tones and falling prices will result from the overcapacity of l-lysine synthesis8. In this context, it would be desirable and economically feasible to produce 5-aminovalerate using l-lysine as a starting material.
Significant researches have been devoted to explore methods for 5-aminovalerate production, for example, enzymatic synthesis18,19 and whole cell biotransformation20. It is universally believed that whole cells are more stable catalyst compared to isolated enzymes due to the protected environment provided by the cell membranes. However, the cell membranes could also be a main barrier for the reactions between substrates and the catalytic enzymes21. Meanwhile, the production process is limited by the accumulation of final products in the cytoplasm22. One general and efficient solution to these problems is to overexpress transport proteins. Bacterial transport proteins are kinds of transmembrane binding proteins of physiological importance in selective permeation of certain molecular. The metabolic process is accelerated with enhancement of substrate import and transportation of final metabolites from cytoplasmic enzymes. In this study, we constructed a novel recombinant Escherichia coli strain by overexpressing transport proteins (Fig. 1), which exhibited good 5-aminovalerate production performance.

Scheme for the production of 5-aminovalerate from l-lysine by engineered E. coli.
DavB, lysine 2-monooxygenase; DavA, δ-aminovaleramidase; LysP, lysine specific permease; PP2911, 4-aminobutyrate transporter.
Results
Feasibility of 5-aminovalerate production by recombinant E. coli
Firstly, the genes davA (gene ID: 1044092) and davB (gene ID: 1044093) of P. putida were inserted into the MCS1 of pETDuet-1 (Fig. S1A) to construct pETDuet-DavAB (Fig. S1B) which was then transferred into E. coli BL21(DE3). The 5-aminovalerate production performance of the resulting recombinant strain E. coli pDAB was verified accompany with E. coli pD harboring blank vector pETDuet-1 and E. coli pDB harboring recombinant plasmid pETDuet-DavB (Fig. S1D). The details for genetic manipulation and catalytic reaction process are presented in “Methods” section.
As shown in Fig. 2, E. coli pD was unable to assimilate l-lysine and no 5-aminovalerate was accumulated. The inability of E. coli pD to consume l-lysine implied the suitability of E. coli as a host for efficient l-lysine conversion. E. coli pDAB was capable of converting 14.6 g/L l-lysine into 10.1 g/L 5-aminovalerate in 24 h, at a yield of 0.86 mol/mol. Although l-lysine was consumed, no 5-aminovalerate was detected during the whole reaction process with E. coli pDB. These results suggested that both DavB and DavA should work efficiently in recombinant E. coli for the 5-aminovalerate production. On the other hand, the E. coli pDB exhibited a higher l-lysine consumption rate than that of E. coli pDAB (Fig. S2A), implying the intracellular 5-aminovalerate might constrain the conversion of l-lysine.

Verification of 5-aminovalerate production by different recombinant E. coli strains.
Results are means ± SD of three parallel replicates.
Effect of transport proteins on 5-aminovalerate production
To elevate 5-aminovalerate production, an improvement of l-lysine absorption was proposed to serve as an efficient approach. LysP is the specific lysine permease in E. coli with high affinity to l-lysine (apparent Km of 2.5 μM)23. The gene lysP (gene ID: 946667) in E. coli K12 was amplified and integrated into the MCS2 of pETDuet-DavAB (Fig. S1C) to construct strain E. coli pDABL. Protein PP2911 of P. putida KT2440 was annotated as 4-aminobutyrate transporter. 4-Aminobutyrate transporters from Arabidopsis thaliana24 and mammals25 were capable of transporting 5-aminovalerate in addition to 4-aminobutyrate. In order to increase the export rate of 5-aminovalerate, the gene pp2911 (gene ID: 1042844) from P. putida KT2440 was inserted into pACYCDuet-1 (Fig. S1E) to construct pACYCDuet-PP2911 (Fig. S1F). It was then transferred into E. coli pDAB to construct E. coli pDABP. The 5-aminovalerate production performance of E. coli pDABL and E. coli pDABP was then examined.
As shown in Table 1, the recombinant strain E. coli pDABP overexpressing DavB, DavA and PP2911 enabled to convert 22.4 g/L of l-lysine to 16.2 g/L 5-aminovalerate in 24 h. The introduction of PP2911 into E. coli pDAB leads to a production improvement by 60.4% and a yield increase to 0.90 mol/mol. However, E. coli pDABL yielded 6.9 g/L 5-aminovalerate from 11.5 g/L l-lysine in 24 h, which failed to show advantage compared with E. coli pDAB (Fig. S2). According to the results, the export of 5-aminovalerate rather than the absorption of l-lysine acted as the major factor limiting 5-aminovalrate production.
In order to explore whether the co-expression of LysP and PP2911 has positive effect on 5-aminovalerate production, the plasmid pACYCDuet-PP2911 was transferred into E. coli pDABL to construct E. coli pDABLP. As expected, the recombinant strain E. coli pDABLP successfully produced 16.9 g/L 5-aminovalerate from 22.5 g/L l-lysine in 24 h with a yield of 0.94 mol/mol (Table 1). Among all the constructed strains, the recombinant strain E. coli pDABLP overexpressing transport proteins LysP and PP2911 attained the best 5-aminovalerate production performance in titer, productivity and yield.
Optimization of biocatalysis conditions of E. coli pDABLP
To elevate 5-aminovalerate productivity of E. coli pDABLP, the biocatalysis conditions were optimized. The effect of reaction temperature on production rate was determined. The influence of l-lysine concentration on the yield of 5-aminovalerate was examined. The conversion rate and catalytic activity which was defined as the 5-aminovalerate produced by per unit biocatalyst per hour were determined to optimize the biocatalyst concentrations. The specific optimization process is addressed in “Methods” section.
As shown in Fig. 3A, the optimal temperature was determined to be 30 °C. The 5-aminovalerate concentration increased as reaction temperature increased up to 30 °C and decreased thereafter. The yield of 5-aminovalerate maintained over 0.80 mol/mol when the l-lysine concentration below 40 g/L and decreased sharply with higher l-lysine concentration (Fig. 3B). Thus, 40 g/L of l-lysine were chosen for further study. As expected, the conversion rate increased with the increase of biocatalyst concentration (Fig. 3C), while the catalytic activity decreased (Fig. 3D). Taken all associated factors into consideration, an OD600nm of 60 was used for further research. Finally, the optimum catalytic conditions were determined to be reaction temperature, 30 °C; substrate concentration, 40 g/L; and biocatalyst concentration, OD600nm = 60.

Optimization of biocatalysis conditions using strain E. coli pDABLP.
(A) Temperature. (B) l-Lysine concentration. (C,D) Biocatalyst concentration. Results are means ± SD of three parallel replicates.
Production of 5-aminovalerate under optimal conditions
According to the results above, the bioconversion of l-lysine into 5-aminovalerate was conducted under optimized conditions. As shown in Fig. 4A, 29.6 g/L 5-aminovalerate was produced from 38.6 g/L l-lysine in 48 h. The yield of 5-aminovalerate was 0.96 mol/mol. Then the fed-batch process was performed to evaluate the production capacity of E. coli pDABLP. The time-course of 5-aminovalerate production from l-lysine was shown in Fig. 4B. The 5-aminovalerate concentration increased continuously in 156 h to a final titer of 63.2 g/L. In total, 102.3 g/L l-lysine was consumed. The yield of 5-aminovalerate from l-lysine was 0.77 mol/mol. The lower yield compared with batch biotransformation process might be caused by the instability of DavA, as mentioned in the previous research18.

Batch (A) and fed-batch (B) process of 5-aminovalerate production from l-lysine by the recombinant strain E. coli pDABLP under optimized conditions. The reactions were carried out in a 300-ml conical flask containing 50 ml reaction mixture: 1/15 M PBS (pH 7.0), l-lysine and cell suspension (OD600nm = 60). The reaction broth was incubated in a water baths shaker at 30 °C with shaking at 120 rpm. For the batch biotransformation process, results are means ± SD of three parallel replicates.
Discussion
Bio-based and biodegradable polymers have drawn worldwide attention as a result of environmental concerns and stress of limitation of petroleum resources. Polylactides stands at the forefront of practical use. Besides lactate, the biomonomers used to form polymers have been widely extended to succinic acid, 1,4-butanediol, 1,3-propanediol, terephthalic acid et al.26,27. 5-Aminovalerate, the precursor of nylon-5, has attained much attention recently5,18,19,20,28. l-Lysine, the original material of 5-aminovalerate, is generally manufactured by direct microbial fermentation of molasses or starch. Since the overcapacity of l-lysine synthesis has resulted in its falling price, the biotransformation of l-lysine, such as 5-aminovalerate production, might extend the scope of application and be beneficial to l-lysine industry.
Membrane-associated transporter proteins play an irreplaceable role in signal transduction, osmotic regulation and substance transportation. Metabolic engineering of transport route has long been applied to speed up the reaction and promote the production of aimed compounds. The lysine specific permease, LysP, is able to accelerate l-lysine uptake by 10- to 20-fold29. The expression of LysP was regulated by extracellular concentration of lysine and was shut off when l-lysine concentration was higher than 25 μM30. In a previously study, overexpression of LysP in E. coli BL21(DE3) caused the improvement of l-pipecolic acid production by 5-fold31. In this context, it is beneficial for LysP expression in l-lysine based biotransformation process. 4-Aminobutyrate transporters from different kingdoms showed high affinity for 5-aminovalerate in addition to 4-aminobutyric acid. In this work, LysP was overexpressed for lysine import while PP2911 was overexpressed for 5-aminovalerate export. PP2911 alone caused 5-aminovalerate production improvement by 60.4% and a yield increased to 0.90 mol/mol. Co-expression of LysP and PP2911 further improved the 5-aminovalerate production performance of the recombinant strain.
Several biotechnological routes have been used to produce 5-aminovalerate from l-lysine. Enzymatic synthesis of 5-aminovalerate from l-lysine was achieved by using lysine α-oxidase from Trichoderma viride19 or by coupling DavA and DavB from P. putida18. The maximum production of 5-aminovalerate catalysed by purified enzymes was 20.8 g/L18. Owing to the complexity of implementation and high operating cost, enzymatic production of 5-aminovalerate is still impractical in industrial field. Park et al. converted 7 g/L l-lysine into 3.6 g/L 5-aminovalerate through co-expression of davAB gene of P. putida in E. coli W31105. Considering the low yield of 5-aminovalerate, they developed a new fed-batch fermentation process through which the mixture of l-lysine, glucose and MgSO4 was gradually added to reactor. At last, 5-aminovalerate was accumulated to a titer of 90.6 g/L20. In the present study, a new recombinant strain E. coli pDABLP based on E. coli BL21(DE3) was constructed to produce 5-aminovalerate from l-lysine. By overexpressing transport proteins LysP and PP2911, the result recombinant strain was able to transform more l-lysine than the strain only expressing DavA and DavB during batch biotransformation and the yield of 5-aminovalerate increased from 0.86 to 0.94 mol/mol. Although 63.2 g/L 5-aminovalerate was produced through a fed-batch transformation process, a higher concentration of 5-aminovalerate is expected to be achieved once the fermentative system developed by Park et al. is introduced.
5-Aminovaleramide, the sole intermediate of 5-aminovalerate production, belongs to primary amides with potential application value. The rapid l-lysine consumption rate of E. coli pDB implied the feasibility of 5-aminovaleramide production while E. coli lacks the enzyme to transform 5-aminovalerate as far as we know. The content of 5-aminovaleramide was not quantified because of the lack of commercially available 5-aminovaleramide. In the fed-batch biotransformation process (Fig. 4B), the continually l-lysine consumption in 156 h suggested the stability of DavB. In this regard, the recombinant E. coli pDB could serve for the efficient production of 5-aminovaleramide from l-lysine.
Production of C3, C4 platform chemicals such as glycerol, lactate, 3-hydroxypropinate, succinate, 2,3-butanediol, 1,4-butanediol and butanediamine32,33 has been extensively studied in recent years. However, only a few researches have focused on C5 platform chemicals production34,35. l-Lysine might serve as a suitable raw material for production of C5 difunctional alkanes. The l-lysine decarboxylase catalyzed the decarboxylation of l-lysine to produce cadaverine which could transform into 5-aminovaleraldehyde through subsequent transamination36. 5-Aminovaleraldehyde could be oxidized by 4-aminobutyraldehyde dehydrogenase to produce 5-aminovalerate37. Then, 5-aminovalerate could convert into glutarate by using δ-aminovalerate aminotransferase (DavT) and glutaric semialdehyde dehydrogenase (DavD)6. Glutarate semialdehyde, the only intermediate metabolite of glutarate production, could also be reduced into 5-hydroxyvalerate by NADH at the presence of 5-hydroxyvalerate dehydrogenase38. Accordingly, it is desirable and feasible to explore methods for biochemical production of C5 platform chemicals from directed biotransformation of l-lysine. In this study, the transformation of l-lysine into 5-aminovalerate was realized and accelerated with the co-expression of transport proteins. Further conversion of l-lysine into glutarate via 5-aminovalerate is under way based on the achieving results.
Conclusion
We constructed a series of recombinant E. coli strains based on the aminovalerate pathway in P. putida to produce 5-aminovalerate from l-lysine. Acceleration of substrate import and product export was carried out by overexpressing transport proteins LysP and PP2911. The final constructed strain E. coli pDABLP exhibited good 5-aminovalerate production performance and might serve as an important biocatalyst for 5-aminovalerate production from renewable biomass.
Methods
Materials
l-Lysine, 5-aminovalerate and isopropyl-β-d-1-thiogalactopyranoside (IPTG) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Polymerase chain reaction (PCR) primers were obtained from Sangon (Shanghai, China). Ampicillin and chloramphenicol were purchased from Amresco (USA). Restriction enzymes were obtained from Fermentas (Lithuania). T4 DNA ligase and fastPfu DNA polymerase were purchased from MBI (USA) and Transgen Biotech (China), respectively. All other chemicals were of analytical grade and commercially available.
Strains, plasmids, genetic methods and culture conditions
E. coli DH5α was used as the host strain for gene manipulation. E. coli BL21(DE3) was used for protein expression. The genes coding DavAB, DavB and PP2911 were amplified by PCR from genomic DNA of P. putida KT2440 using primers DavAB.f/DavAB.r, DavAB.f/DavB.r and PP2911.f/PP2911.r (Table 2) respectively. The lysP gene was cloned from E. coli K12 chromosomal DNA with primers LysP.f/LysP.r (Table 2). The resulting DNA fragments were digested with corresponding restriction endonuclease and inserted into expression vector pETDuet-1 (Fig. S1A) or pACYCDuet-1 (Fig. S1E).
All of the E. coli strains were cultivated in Luria-Bertani (LB) medium at 37 °C and 180 rpm. When necessary, ampicillin (100 μg/ml) and chloramphenicol (40 μg/ml) were added.
Biotransformation by recombinant E. coli strains
For whole cell biocatalyst preparation, the recombinant E. coli strains harboring different plasmids were cultivated to the early exponential phase (OD600nm = 0.4–0.6) at 37 °C. Then, 1 mM IPTG was added to the culture medium to induce protein expression. The culture was incubated at optimized temperature of 25 °C for additional 12 h. Cells were harvested by centrifugation (6000 rpm, 10 min), washed twice with 1/15 M phosphate buffer solution (PBS, pH 7.0).
To verify the 5-aminovalerate production by recombinant E. coli strains and the effect of transport proteins on 5-aminovalerate production, reactions were carried out in a 50-ml conical flask containing 10 ml reaction mixture: 1/15 M PBS (pH 7.0), about 20 g/L l-lysine and cell suspension (OD600nm = 30). The reaction broth was incubated in a water baths shaker at 30 °C with shaking at 120 rpm. Samples (0.2 ml) were withdrawn at timed intervals and centrifuged (12,000 rpm, 15 min) before analysis.
Optimization of biocatalysis conditions
The effects of catalytic conditions on biotransformation were determined in a 50-ml flask containing 10 ml reaction mixture: 1/15 M PBS (pH 7.0), l-lysine and cell suspension at 120 rpm. Only one parameter changed once while the others remained constant. To determine the effect of temperature on 5-aminovalerate production, the initial concentration of l-lysine was set as approximately 20 g/L. The OD600nm of the cell suspension was set as 30. Samples were taken after incubation of 3 h at various temperatures (20, 25, 30, 37, 42, 50 °C). The effect of l-lysine concentration on the yield of 5-aminovalerate was determined at 30 °C and the OD600nm was set as 30. Samples were taken after incubation of 3 h with different l-lysine concentrations (10, 20, 30, 40, 50, 60 g/L). To determine the effect of the whole cell concentration, the reactions were conducted with approximately 40 g/L l-lysine at 30 °C. Samples were taken after incubation of 12 h with various concentrations of whole cells (OD600nm = 15, 30, 45, 60, 75). The samples (0.2 ml) were centrifuged (12,000 rpm, 15min) before derivatization and analyzation.
Analytical methods
Cell density was measured at 600 nm (OD600nm) using an Ultrospec™ 2100 pro UV/visible spectrophotometer. Cells were diluted to suitable multiples in 1/15 M PBS (pH 7.0) to ensure the accurate measurement. 5-Aminovalerate and l-lysine concentration were quantified using a high-performance liquid chromatography (HPLC) system (Agilent 1100 series, Hewlett-Packard) equipped with a UV-Vis detector18. The samples were diluted to suitable multiples and 400 μl of diluted samples was derivatized with phenylisothiocyanate (PITC)-acetonitrile (100 mM, 200 μl) and triethylamine-acetonitrile (1 M, 200 μl) for 1 h at room temperature before being extracted by n-hexane (800 μl). The lower aqueous phase was injected, separated on an Eclipse XDB-C18 (5 μm, 4.6 mm × 150 mm) maintained at 38 °C39 and detected at 254 nm. The mobile phase (at a flow rate of 0.6 ml/min) was composed of a solution of (A) pH 6.5, 100 mM ammonium acetate-acetonitrile (97:3) and (B) acetonitrile. The percentage of A declined from 82% to 70% in the first 20 minutes and maintained at 70% in the following 20 minutes.
Additional Information
How to cite this article: Li, Z. et al. Overexpression of transport proteins improves the production of 5-aminovalerate from l-lysine in Escherichia coli. Sci. Rep. 6, 30884; doi: 10.1038/srep30884 (2016).
References
Kawasaki, N. et al. Synthesis, thermal and mechanical properties and biodegradation of branched polyamide 4. Polymer 46, 9987–9993 (2005).
Carlson, E. & Nelson, K. Nylon under the hood: a history of innovation. Automotive engineering 104, 84–89 (1996).
Kudo, K., Mochizuki, M., Kiriyama, S., Watanabe, M. & Hirami, M. Studies on the structure and properties of nylon 46 fiber. I. Dimensional stability. J. Appl. Polym. Sci. 52, 861–867 (1994).
Bermúdez, M., León, S., Alemán, C. & Muñoz-Guerra, S. Comparison of lamellar crystal structure and morphology of nylon 46 and nylon 5. Polymer 41, 8961–8973 (2000).
Park, S. J. et al. Metabolic engineering of Escherichia coli for the production of 5-aminovalerate and glutarate as C5 platform chemicals. Metab. Eng. 16, 42–47 (2013).
Revelles, O., Espinosa-Urgel, M., Fuhrer, T., Sauer, U. & Ramos, J. L. Multiple and interconnected pathways for l-lysine catabolism in Pseudomonas putida KT2440. J. Bacteriol. 187, 7500–7510 (2005).
Zhou, L.-B. & Zeng, A.-P. Engineering a lysine-ON riboswitch for metabolic control of lysine production in Corynebacterium glutamicum. ACS Synth. Biol. 4, 1335–1440 (2015).
Eggeling, L. & Bott, M. A giant market and a powerful metabolism: l-lysine provided by Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 99, 3387–3394 (2015).
Chen, Z., Bommareddy, R. R., Frank, D., Rappert, S. & Zeng, A.-P. Deregulation of feedback inhibition of phosphoenolpyruvate carboxylase for improved lysine production in Corynebacterium glutamicum. Appl. Environ. Microbiol. 80, 1388–1393 (2014).
Bommareddy, R. R., Chen, Z., Rappert, S. & Zeng, A.-P. A de novo NADPH generation pathway for improving lysine production of Corynebacterium glutamicum by rational design of the coenzyme specificity of glyceraldehyde 3-phosphate dehydrogenase. Metab. Eng. 25, 30–37 (2014).
Neuner, A. et al. Production of l-lysine on different silage juices using genetically engineered Corynebacterium glutamicum. J. Biotechnol. 163, 217–224 (2013).
Geng, F., Chen, Z., Zheng, P., Sun, J. & Zeng, A.-P. Exploring the allosteric mechanism of dihydrodipicolinate synthase by reverse engineering of the allosteric inhibitor binding sites and its application for lysine production. Appl. Microbiol. Biotechnol. 97, 1963–1971 (2013).
Chen, Z., Rappert, S., Sun, J. & Zeng, A.-P. Integrating molecular dynamics and co-evolutionary analysis for reliable target prediction and deregulation of the allosteric inhibition of aspartokinase for amino acid production. J. Biotechnol. 154, 248–254 (2011).
Chen, Z., Meyer, W., Rappert, S., Sun, J. & Zeng, A.-P. Coevolutionary analysis enabled rational deregulation of allosteric enzyme inhibition in Corynebacterium glutamicum for lysine production. Appl. Environ. Microbiol. 77, 4352–4360 (2011).
Becker, J., Zelder, O., Häfner, S., Schröder, H. & Wittmann, C. From zero to hero—Design-based systems metabolic engineering of Corynebacterium glutamicum for l-lysine production. Metab. Eng. 13, 159–168 (2011).
Jakobsen, Ø. M. et al. Overexpression of wild-type aspartokinase increases l-lysine production in the thermotolerant methylotrophic bacterium Bacillus methanolicus. Appl. Environ. Microbiol. 75, 652–661 (2009).
Becker, J., Klopprogge, C., Zelder, O., Heinzle, E. & Wittmann, C. Amplified expression of fructose 1,6-bisphosphatase in Corynebacterium glutamicum increases in vivo flux through the pentose phosphate pathway and lysine production on different carbon sources. Appl. Environ. Microbiol. 71, 8587–8596 (2005).
Liu, P. et al. Enzymatic production of 5-aminovalerate from l-lysine using l-lysine monooxygenase and 5-aminovaleramide amidohydrolase. Sci. Rep. 4, 5657 (2014).
Pukin, A. V., Boeriu, C. G., Scott, E. L., Sanders, J. P. & Franssen, M. C. An efficient enzymatic synthesis of 5-aminovaleric acid. J. Mol. Catal. B: Enzym. 65, 58–62 (2010).
Park, S. J. et al. High-level conversion of l-lysine into 5-aminovalerate that can be used for nylon 6,5 synthesis. Biotechnol. J. 9, 1322–1328 (2014).
de Carvalho, C. C. R. Enzymatic and whole cell catalysis: finding new strategies for old processes. Biotechnol. Adv. 29, 75–83 (2011).
Boyarskiy, S. & Tullman-Ercek, D. Getting pumped: membrane efflux transporters for enhanced biomolecule production. Curr. Opin. Chem. Biol. 28, 15–19 (2015).
Rauschmeier, M., Schüppel, V., Tetsch, L. & Jung, K. New insights into the interplay between the lysine transporter LysP and the pH sensor CadC in Escherichia coli. J. Mol. Biol. 426, 215–229 (2014).
Meyer, A., Eskandari, S., Grallath, S. & Rentsch, D. AtGAT1, a high affinity transporter for γ-aminobutyric acid in Arabidopsis thaliana. J. Biol. Chem. 281, 7197–7204 (2006).
Jack, D. L., Paulsen, I. T. & Saier, M. H. The amino acid/polyamine/organocation (APC) superfamily of transporters specific for amino acids, polyamines and organocations. Microbiology 146, 1797–1814 (2000).
Reddy, M. M., Vivekanandhan, S., Misra, M., Bhatia, S. K. & Mohanty, A. K. Biobased plastics and bionanocomposites: Current status and future opportunities. Prog. Polym. Sci. 38, 1653–1689 (2013).
Sudesh, K. & Iwata, T. Sustainability of biobased and biodegradable plastics. Clean-Soil Air Water 36, 433–442 (2008).
Adkins, J., Jordan, J. & Nielsen, D. R. Engineering Escherichia coli for renewable production of the 5-carbon polyamide building-blocks 5-aminovalerate and glutarate. Biotechnol. Bioeng. 110, 1726–1734 (2013).
Steffes, C., Ellis, J., Wu, J. & Rosen, B. P. The lysP gene encodes the lysine-specific permease. J. Bacteriol. 174, 3242–3249 (1992).
Ruiz, J., Haneburger, I. & Jung, K. Identification of ArgP and Lrp as transcriptional regulators of lysP, the gene encoding the specific lysine permease of Escherichia coli. J. Bacteriol. 193, 2536–2548 (2011).
Fujii, T., Aritoku, Y., Agematu, H. & Tsunekawa, H. Increase in the rate of l-pipecolic acid production using lat-expressing Escherichia coli by lysP and yeiE amplification. Biosci. Biotechnol. Biochem. 66, 1981–1984 (2002).
Chaabouni, E., Sarma, S. J., Gassara, F. & Brar, S. K. In Biotransformation of waste biomass into high value biochemicals (eds Brar, S. K., Dhillon, G. S. & Soccol, C. R. ) 473–489 (Springer, New York, 2014).
Qian, Z.-G., Xia, X.-X. & Lee, S. Y. Metabolic engineering of Escherichia coli for the production of putrescine: a four carbon diamine. Biotechnol. Bioeng. 104, 651–662 (2009).
Kind, S. et al. From zero to hero–Production of bio-based nylon from renewable resources using engineered Corynebacterium glutamicum. Metab. Eng. 25, 113–123 (2014).
Qian, Z.-G., Xia, X.-X. & Lee, S. Y. Metabolic engineering of Escherichia coli for the production of cadaverine: a five carbon diamine. Biotechnol. Bioeng. 108, 93–103 (2011).
Samsonova, N. N., Smirnov, S. V., Altman, I. B. & Ptitsyn, L. R. Molecular cloning and characterization of Escherichia coli K12 ygjG gene. BMC Microbiol. 3, 1 (2003).
Yorifuji, T., Koike, K., Sakurai, T. & Yokoyama, K.-i. 4-Aminobutyraldehyde and 4-guanidinobutyraldehyde dehydrogenases for arginine degradation in Pseudomonas putida. Agric. Biol. Chem. 50, 2009–2016 (1986).
Barker, H. A., D’Ari, L. & Kahn, J. Enzymatic reactions in the degradation of 5-aminovalerate by Clostridium aminovalericum. J. Biol. Chem. 262, 8994–9003 (1987).
Chen, S. The facile HPLC enantioresolution of amino acids, peptides on naphthylethylcarbamate-β-cyclodextrin bonded phases using the acetonitrile-based mobile phase after their pre-column derivatization with phenyl isothiocyanate: factors that affect the resolution. Amino Acids 26, 291–298 (2004).
Acknowledgements
The work was supported by Chinese National Program for High Technology Research and Development (2014AA021201), National Natural Science Foundation of China (31470199) and the Young Scholars Program of Shandong University (2015WLJH25).
Author information
Authors and Affiliations
Contributions
C.G., C.M. and P.X. participated in the design of the study and provided reagents and materials. Z.L., J.X., T.J., Y.G., P.L., M.Z. and Z.S. executed the experimental work. Z.L., C.G. and C.M. analyzed the data. Z.L., C.G., C.M. and P.X. wrote and revised the manuscript. All authors read and approved the final manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Li, Z., Xu, J., Jiang, T. et al. Overexpression of transport proteins improves the production of 5-aminovalerate from l-lysine in Escherichia coli. Sci Rep 6, 30884 (2016). https://doi.org/10.1038/srep30884
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep30884
This article is cited by
-
Efficient whole-cell catalysis for 5-aminovalerate production from L-lysine by using engineered Escherichia coli with ethanol pretreatment
Scientific Reports (2020)
-
Biotechnological production of mono- and diamines using bacteria: recent progress, applications, and perspectives
Applied Microbiology and Biotechnology (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
