
Abstract
Introduced predators have caused declines and extinctions of native species worldwide, seemingly able to find and hunt new, unfamiliar prey from the time of their introduction. Yet, just as native species are often naïve to introduced predators, in theory, introduced predators should initially be naïve in their response to novel native prey. Here we examine the response of free-living introduced red foxes (Vulpes vulpes) to their first encounter with the odour cues of a novel native prey, the long-nosed bandicoot (Perameles nasuta). Despite no experience with bandicoots at the study site, foxes were significantly more interested in bandicoot odour compared to untreated controls and to a co-evolved prey, the black rat (Rattus rattus). So what gives introduced predators a novelty advantage over native prey? Such neophilia towards novel potential food sources carries little costs, however naïve native prey often lack analogous neophobic responses towards novel predators, possibly because predator avoidance is so costly. We propose that this nexus between the costs and benefits of responding to novel information is different for alien predators and native prey, giving alien predators a novelty advantage over native prey. This may explain why some introduced predators have rapid and devastating impacts on native fauna.
Similar content being viewed by others

Introduction
The decline and extinction of native species has repeatedly coincided with the arrival and spread of introduced predators1, which have at least double the impact on prey populations as native predators2. Introduced predator impacts are often most severe during the acute phase of the invasion process, compared to the chronic phase where native prey may learn and adapt to novel enemies. Hence the initial response of predators and prey to each others’ cues is arguably the most important phase in the predation sequence3. The exaggerated impacts of alien predators are usually attributed to a lack of effective anti-predator strategies by native prey towards these predators (naïve prey hypothesis)4, which hinges on the assumption that introduced predators enjoy a novelty advantage that facilitates their invasion success5. However, given that naiveté towards novel species can go both ways, introduced predators should also be initially naïve to native prey5.
Behavioural flexibility is thought to be a key factor in the success of introduced predators, enabling invaders to rapidly respond to novel situations6. Such flexibility can include a predilection to consume unfamiliar foods and a positive responsiveness to novelty7. When invading a new environment, there is likely to be strong selection for individuals that demonstrate behavioural flexibility to readily switch between prey types and investigate cues of unfamiliar prey, allowing them to quickly become efficient predators of novel prey types. For example, actively invading house sparrows (Passer domesticus) are more willing to approach and consume novel foods than individuals in an established population7. However, although there are numerous studies on animals rapidly learning about novel food items in a laboratory environment (e.g.8,9), there is a general lack of understanding of how wild living invasive predators respond to novel prey cues on their initial encounter.
Olfactory cues are crucial to reciprocal recognition in many mammalian predator-prey interactions, yet the role of a shared evolutionary history in the exploitation of odours is not well understood10. Field-based experiments on free-living predator responses to evolutionary novel prey cues are an obvious gap in the literature. The few studies that have attempted to fill this niche are predominantly aquatic studies (e.g.11) and can not rule out the possibility that predators may have a pre-existing learned associative relationship with the novel prey due to a past encounter.
Here we test the responses of a wild free-living introduced predator, the red fox (Vulpes vulpes), to their first encounter with odour cues of a vulnerable Australian native prey species, the long-nosed bandicoot (Perameles nasuta). Foxes have been implicated as a major cause of declines and extinctions of Australian critical weight range (CWR; 35–5500 g) native mammals1 and five of the other seven bandicoot species in Australia are extinct or critically endangered due to alien predators12. Long-nosed bandicoots are coastal CWR mammals that are naïve to foxes13 and vulnerable to fox predation, which is known to suppress their populations14. We also examine fox responses on first encounter with a co-evolved prey species, the black rat (Rattus rattus). Black rats arrived in Australia ca. 200 years ago with settlers from Europe15 where they have coexisted with foxes for thousands of years16, and are eaten by foxes elsewhere in Australia (e.g. see17). Our remote inland study site is well outside the known ranges of both prey species and their close relatives18 and so local foxes would have no experience with these species. We test predictions of two alternate hypotheses (1) that foxes will investigate bandicoot and rat odours despite a lack of familiarity (behavioral flexibility hypothesis) or (2) that foxes will not investigate the evolutionary novel bandicoot odours (naiveté hypothesis).
Results
Foxes investigated significantly more sand plots with bandicoot odours than sand plots with rat or control odours (Pearson Chi-Square X2 = 7.54, DF = 2 exact sig. (2-sided) P = 0.024, adjusted standardized residual of 2.7). More than 80% of sand plots with bandicoot odour were investigated by foxes after two nights, compared to just 17% and 20% of the rat odour and control sand plots respectively (Fig. 1).

Foxes investigated sand plots with bandicoot odours more frequently than sand plots with rat or control odours.
Discussion
Despite no prior experience with bandicoots, foxes showed greater interest in the evolutionary unfamiliar bandicoot odour than to untreated controls, supporting the behavioral flexibility hypothesis. Strong fox interest in the nesting odours of a marsupial species such as bandicoots, which are evolutionary distinct from any prey that foxes have co-evolved with in their native range, may help to explain the success and rapid impacts of introduced foxes in Australia and elsewhere. While foxes used in this study were from established populations and it is possible that they may behave differently on the invasion front, curiosity to novelty is likely to be similar and many of the foxes at our study site would have been dispersing individuals moving through unfamiliar environments. Identifying evolutionary processes of long-established introduced species is difficult. Although there is a remote possibility that the ancestors of the foxes at our study site could have encountered bandicoot sp. as they spread from their release site near Melbourne, Victoria in the 1870s19, it is unlikely. Foxes spread rapidly across Victoria after release, arriving in Walpeup within approximately 20 years19, limiting the opportunity for genetic selection and retention of specific recognition of bandicoot odour. Furthermore, although there is speculation that other bandicoots species might have once occurred in this region, their exact distributions are unknown20. We argue that it seems more likely that the rapid attraction to bandicoot odour by foxes reflects a general predator strategy, particularly of introduced predators, to be attracted to novelty. So what gives introduced predators the novelty advantage over native prey?
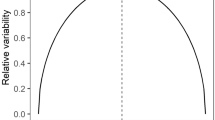
Investigating unfamiliar prey cues (neophilia) can carry relatively little cost to introduced predators for a potentially large reward; if investigation is rewarded, predators can rapidly develop search images21 that can be devastating for naïve prey. In contrast, native prey may suffer considerable missed opportunity costs should they forgo foraging by performing unnecessary anti-predator responses upon initial encounter with unfamiliar predator cues (neophobia22) and will rapidly habituate to cues of risk that are not realised23. We propose that this represents a neophilia-neophobia nexus (Fig. 2) reflecting an imbalance in the cost:benefit relationship for alien predators and native prey responding to novel information about their enemy. In time, naïve prey can develop refined recognition and responses to novel predators4 however alien predators displaying neophilia skip over this initial learning stage, giving them an advantage, which may help to explain why so many native species rapidly decline after the arrival of alien predators. Our results provide preliminary support for this model, however it should be considered as a theoretical basis for future experimental testing.

Predators should be attracted to prey cues and pay low MOC for doing so, whereas if prey respond to predator cues, this response carries high MOC.
That foxes showed greater interest in bandicoot than rat odours was surprising. The lack of fox interest in the odours of black rats may be because they were specifically recognised as an unprofitable prey. Black rats likely possess more effective anti-predator strategies against foxes compared to bandicoots given their shared evolutionary history, making them a more difficult prey option. It is also possible that attraction to the evolutionary novel prey may reflect a general inherent preference for novel prey by alien predators. Novel prey are likely to be naïve to foxes and have ineffective anti-predator responses5. There is some support for this idea: foxes continued to depredate an already reduced population of bettongs (another Australian CWR marsupial) amongst a high-density rabbit (Oryctolagus cunniculus) population (an evolutionary familiar prey)24.
The exaggerated impacts of introduced predators on native prey are commonly attributed to shortcomings in native prey5. However the outcome of a predator-prey interaction is determined by tactics of both predators and prey3. Our results suggest that neophilic tactics of alien predators towards novel information during initial encounters with potential prey may tip the novelty advantage towards predators. Alien predator impacts will be greatest when these neophilic tactics occur during the acute phase of invasion before native prey can develop effective anti-predator tactics. Innovative management strategies are needed to undermine this introduced predator novelty advantage (e.g.25).
Methods
Odour collection
Odours indicative of nesting long-nosed bandicoots and black rats were used, as both species rest in den sites and are vulnerable to olfactory predators14,26. Animal odours (body, urine, faeces) were collected on towels (20 × 30 cm) placed in cage traps for one night at North Head, Sydney, Australia. Scented towels were sealed in airtight bags and stored at −20 °C which is unlikely to affect odour degradation23. Controls were clean towels, and separate gloves were worn when handling samples to prevent cross-contamination of odours.
Experimental method
Seventy-two sand monitoring plots (2 m wide) were constructed >500 m apart using in situ sand across dirt roads over a 225 km2 area of agricultural land and remnant mallee woodland at Walpeup, north-western Victoria, Australia (>800 km from where odours were collected and >300 km from the nearest bandicoot record; rats can occur in major rural centres, for example Mildura, but rarely outside27).
Foxes were habituated to the experimental set-up to enable separation of initial responses to the set-up from responses to odour treatments. This involved a familiarisation phase whereby a clean towel (20 × 30 cm) was pinned immediately adjacent to the edge of the sand plot with a tent peg and lightly covered with in situ sand to reduce visual conspicuousness. Foxes within the vicinity of a sand plot were considered to have familiarised to the plot if the level of interest in the clean towel decreased after 4–13 days.
Fox activity on the sand plot was classified as an ‘investigation’ of the experimental equipment based on the spatial arrangement, direction and concentration of footprints within one metre of the towel, (derived from a frequency histogram (Fig. 3); a distinct peak at eight footprints was indicative of an incidental passing i.e. the number of tracks taken to cross the sand pad in a straight line). An ‘investigation’ of the equipment met one of the following criteria: more than ten fox footprints or an obvious change in direction of footprints within one metre of the towel, or diggings at the towel (Fig. 4). All other fox activity on the sand plot was classified as an ‘incidental passing’.

A distinct peak at eight footprints was indicative of an incidental passing.

Investigation: (a) >10 footprints within 1 m of the towel, (b) obvious change in direction, (c) diggings at towel. Incidental passing: (d) ≤10 footprints, (e) no footprints within 1 m of the towel.
It was assumed that interest in the experimental set up had dropped if fox activity declined from being classified as investigations to incidental passing’s or if a plot was repeatedly passed without being investigated. Following these criteria, we considered foxes had become familiar to the experimental equipment at 40 plots.
All plots received treatments on the same day, which involved pegging a treatment towel (long-nosed bandicoot odour, black rat odour, or control) to the edge of the sand plot as in the familiarisation phase. Sand plots were monitored and re-raked daily for three days and an observer blinded to the odour treatments identified all animal prints present. Fox activity was classified as either an investigation or incidental passing as above (Fig. 4). Only plots where foxes had familiarised to the setup and then recorded fox activity during the three treatment nights were included in the analyses (n = 7, 12 and 10 for bandicoot, rat and control treatments respectively).
Statistical analyses
We used a Chi-Square test in PASW Statistic 18 using exact probability values followed by adjusted standardised residuals to identify treatment effects; values greater than two indicate a lack of fit to the null hypothesis in that cell28. Analyses were conducted on the total number of plots that had recorded fox activity after three nights. Plots were scored as investigated if they recorded a fox investigation on any of the three treatment nights, regardless of other visits (incidental passings) to that plot.
Ethics statement
All field methods were carried out in accordance with procedures that were approved by the University of New South Wales Animal Ethics Committee (08/153B).
Additional Information
How to cite this article: Bytheway, J. P. et al. Deadly intentions: naïve introduced foxes show rapid attraction to odour cues of an unfamiliar native prey. Sci. Rep. 6, 30078; doi: 10.1038/srep30078 (2016).
References
Dickman, C. R. Impact of exotic generalist predators on the native fauna of Australia. Wildlife Biology 2, 185–195 (1996).
Salo, P. et al. Alien predators are more dangerous than native predators to prey populations. Proceedings of the Royal Society B: Biological Sciences 274, 1237–1243, doi: 10.1098/rspb.2006.0444 (2007).
Endler, J. A. in Behavioural ecology: an evolutionary approach Vol. 3 (eds J. R. Krebs & Davies. N. B. ) 169–196 (Blackwell Scientific Publications, 1991).
Carthey, A. J. & Banks, P. B. Naivete in novel ecological interactions: lessons from theory and experimental evidence. Biological Reviews doi: 10.1111/brv.12087 (2014).
Sih, A. et al. Predator-prey naivete, antipredator behavior, and the ecology of predator invasions. Oikos 119, 610–621, doi: 10.1111/j.1600-0706.2009.18039.x (2010).
Wright, T., Eberhard, J., Hobson, E., Avery, M. & Russello, M. Behavioral flexibility and species invasions: the adaptive flexibility hypothesis. Ethology Ecology & Evolution 22, 393–404, doi: 10.1080/03949370.2010.505580 (2010).
Martin, L. B. & Fitzgerald, L. A taste for novelty in invading house sparrows, Passer domesticus. Behavioral Ecology 16, 702–707, doi: http://dx.doi.org/10.1093/beheco/ari044 (2005).
Galef, B. G. Jr. & Whiskin, E. E. Preference for novel flavors in adult Norway Rats (Rattus norvegicus). Journal of Comparative Psychology 117, 96, doi: 10.1037/0735-7036.117.1.96 (2003).
Sherwin, C., Heyes, C. & Nicol, C. Social learning influences the preferences of domestic hens for novel food. Animal Behaviour 63, 933–942 (2002).
Banks, P. B., Bytheway, J. P., Carthey, A. J., Hughes, N. K. & Price, C. J. In Carnivores of Australia: Past, Present and Future (eds A. S. Glen & C. R. Dickman ) 389–399 (CSIRO, 2014).
Sanford, E. & Swezey, D. S. Response of predatory snails to a novel prey following the geographic range expansion of an intertidal barnacle. Journal of Experimental Marine Biology and Ecology 354, 220–230, doi: 10.1016/j.jembe.2007.11.007 (2008).
Woinarski, J., Burbidge, A. & Harrison, P. Action Plan for Australian Mammals. (CSIRO, 2014).
Russell, B. G. & Banks, P. B. Responses of four Critical Weight Range (CWR) marsupials to the odours of native and introduced predators. Australian Zoologist 33, 217–222 (2005).
Dexter, N., Meek, P., Moore, S., Hudson, M. & Richardson, H. Population responses of small and medium sized mammals to fox control at Jervis Bay, Southeastern Australia. Pacific Conservation Biology 13, 283–292 (2007).
Banks, P. B. & Hughes, N. K. A review of the evidence for potential impacts of black rats (Rattus rattus) on wildlife and humans in Australia. Wildlife Research 39, 78–88, doi: 10.1071/WR11086 (2012).
Baquero, R. A. & Tellería, J. L. Species richness, rarity and endemicity of European mammals: a biogeographical approach. Biodiversity and Conservation 10, 29–44, doi: 10.1023/A:1016698921404 (2001).
Glen, A., Fay, A. & Dickman, C. Diets of sympatric red foxes Vulpes vulpes and wild dogs Canis lupus in the Northern Rivers Region, New South Wales. Australian Mammalogy 28, 101–104 (2006).
Van Dyke, S. & Strahan, R. The mammals of Australia., (Reed New Holland, 2008).
Abbott, I. The importation, release, establishment, spread, and early impact on prey animals of the red fox Vulpes vulpes in Victoria and adjoining parts of south-eastern Australia. Australian Zoologist 35, 463–533 (2011).
Van Dyck, S. & Strahan, R. The Mammals of Australia 3rd edn, (New Holland Pub Pty Limited, 2008).
Tinbergen, L. The natural control of insects in pinewoods. I. Factors influencing the intensity of predation by songbirds. Archives Neerlandaises de Zoologie 13: 265–336. Ardea 68, 11–30 (1960).
Brown, G. E., Ferrari, M. C., Elvidge, C. K., Ramnarine, I. & Chivers, D. P. Phenotypically plastic neophobia: a response to variable predation risk. Proceedings of the Royal Society of London B: Biological Sciences, 280, 20122712 (2013).
Bytheway, J. P., Carthey, A. J. & Banks, P. B. Risk vs. reward: how predators and prey respond to aging olfactory cues. Behavioral Ecology and Sociobiology 67, 715–725, doi: 10.1007/s00265-013-1494-9 (2013).
Short, J., Kinnear, J. & Robley, A. Surplus killing by introduced predators in Australia—evidence for ineffective anti-predator adaptations in native prey species? Biol. Conserv. 103, 283–301, doi: 10.1016/S0006-3207(01)00139-2 (2002).
Price, C. J. & Banks, P. B. Exploiting olfactory learning in alien rats to protect birds’ eggs. Proceedings of the National Academy of Sciences 109, 19304–19309, doi: 10.1073/pnas.1210981109 (2012).
Rutherford, M., Harper, G. A. & Moller, H. Denning behaviour of ship rats (Rattus rattus) on Taukihepa, a seabird breeding island. New Zealand Journal of Zoology 36, 343–353, doi: 10.1080/03014220909510159 (2009).
Atlas of Living Australia http://www.ala.org.au/.
Agresti, A. A survey of exact inference for contingency tables. Statistical Science 7, 131–153, doi: 10.1214/ss/1177011454 (1992).
Acknowledgements
We thank NSW National Parks and Wildlife Service for their assistance in collecting prey odours. This study was funded by Australian Research Council Discovery Grants DP0881455 and DP140104413 awarded to PBB.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study design, carried out the fieldwork and drafted the manuscript. All authors gave final approval for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Bytheway, J., Price, C. & Banks, P. Deadly intentions: naïve introduced foxes show rapid attraction to odour cues of an unfamiliar native prey. Sci Rep 6, 30078 (2016). https://doi.org/10.1038/srep30078
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep30078
This article is cited by
-
Cats, foxes and fire: quantitative review reveals that invasive predator activity is most likely to increase shortly after fire
Fire Ecology (2023)
-
Exposure to a novel predator induces visual predator recognition by naïve prey
Behavioral Ecology and Sociobiology (2020)
-
Naïve, bold, or just hungry? An invasive exotic prey species recognises but does not respond to its predators
Biological Invasions (2018)
-
Native predators living in invaded areas: responses of terrestrial amphibian species to an Argentine ant invasion
Oecologia (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
