
Abstract
The gut microbiota plays a crucial role in the bi-directional gut–brain axis, a communication that integrates the gut and central nervous system (CNS) activities. Animal studies reveal that gut bacteria influence behaviour, Brain-Derived Neurotrophic Factor (BDNF) levels and serotonin metabolism. In the present study, we report for the first time an analysis of the microbiota–gut–brain axis in zebrafish (Danio rerio). After 28 days of dietary administration with the probiotic Lactobacillus rhamnosus IMC 501, we found differences in shoaling behaviour, brain expression levels of bdnf and of genes involved in serotonin signalling/metabolism between control and treated zebrafish group. In addition, in microbiota we found a significant increase of Firmicutes and a trending reduction of Proteobacteria. This study demonstrates that selected microbes can be used to modulate endogenous neuroactive molecules in zebrafish.
Similar content being viewed by others

Introduction
There is an emerging understanding of the bi-directional crosstalk governing gut-to-brain communication in health and disease of both organs. Accordingly, not only the brain can affect gut functions, but the gut can also induce changes in the central nervous system (CNS) and there is now compelling evidence for various links between the enteric microbiota and brain function1,2,3. This connection is becoming increasingly relevant in novel therapeutic strategies to target psychiatric disorders such as depression and anxiety disorders. As such, it has been shown that absence or modification of the enteric microbiota affects stress-associated anxiety-like and depressive-like behaviours4,5. This has lead to the postulation of psychobiotics, i.e. live microorganisms that have beneficial psychotropic effects on the host6. To date, multiple probiotic bacteria with psychotropic potential have been identified, including strains of the genera Bifidiobacterium7,8,9,10, Lactobacillus9,10,11,12 and Enterococcus13. A very promising potential psychobiotic with demonstrated effectivity in mice and humans is Lactobacillus rhamnosus11,12.
Importantly, the microbiota has been implicated in altering neurotrophic factors7,8,14,15,16,17, a class of proteins playing roles in controlling neuronal function and maintaining cellular integrity, survival, differentiation and synaptic plasticity18,19,20. Among all neurotrophins, Brain Derived Neurotrophic Factor (BDNF) is the most well conserved throughout vertebrate evolution1,21,22. As such, the primary amino acid sequences of zebrafish (Danio rerio) and human BDNF are 91% identical22,23,24. While the distribution pattern of bdnf mRNA and protein has been described in the CNS of rat25, mouse26, human27 and zebrafish28, BDNF has also been observed in other organs and tissues of adult and developing zebrafish29. Changes in BDNF signalling is relevant to a range of human neuronal and psychiatric disorders30, as well as to biological systems involved in the stress response31.
Using rodents as model organisms, multiple studies have also implicated the microbiota in regulation of serotonin signalling and metabolism16,32,33. The monoamine serotonin, or 5-hydroxytryptamine (5-HT), is one of the primary neurotransmitters modulating physiological and behavioural processes in the CNS34,35,36 and the serotonergic system is highly conserved in vertebrate species. Also zebrafish Danio rerio possesses a complex serotonergic system featuring all major genes for 5-HT synthesis, metabolism and signalling37, similar to those observed in humans and rodents38. As such, zebrafish possess three copies of the tph gene (tph1a, tph1b and tph2) encoding tryptophan hydroxylase, the rate-limiting enzyme in serotonin synthesis, two genes encoding serotonin transporters homologous to those of vertebrates, slc6a4a and slc6a4b (previously serta and sertb)34 and multiple receptors homologs, e.g. htr1aa, homologue of the mammalian serotonin receptor 1A and htr1ab, specific of fishes. Zebrafish also have one functional monoamine oxidase gene (mao) exhibiting a strong affinity profile for serotonin37,39. Zebrafish is a well-known model organism, increasingly used in translational neuroscience and behavioural research40. Interestingly, it is a highly social species and shoaling is a typical group forming behaviour often seen in cyprinids (Cyprinidae). Shoaling behaviour has been observed both in nature41 and in the laboratory42,43 as one of the most robust and consistent behavioural features of this species40.
To explore probiotics as potential therapeutic psychotropic agents in psychiatric diseases and to further highlight the use of the zebrafish model to describe the impact of psychobiotics on brain function and behaviour, we here assessed the effects of the probiotic strain Lactobacillus rhamnosus IMC 501 on shoaling behaviour in healthy, wild-type zebrafish. This strain was already used in zebrafish, where it generates beneficial effects such as reduction of hepatic cholesterol level, improved adsorption of nutrients and increased production of short-chain fatty acids44. Moreover, we investigate central and peripheral expression levels of the bdnf gene and of the serotonergic genes tph1a, tph1b, tph2, htr1aa, slc6a4a and mao, in brain and gut of zebrafish. We also evaluated the microbiota profile by using 16S rRNA sequencing after probiotic administration.
Results
L. rhamnosus administration changes shoaling behaviour
Zebrafish shoaling behaviour was analysed for Average Distance (AD), Distance Variance (DV), Nearest Distance (ND), Occupied Area (OA), Column Preference (CP) and Shuttling Frequency (SF) at day 0 and after 4 weeks in both control (CTRL) and probiotic treated (PROBIO) groups. Control group (CTRL) displayed behavioural adaptation to the tank during the 4 weeks of treatment (Fig. 1). In particular, AD and DV significantly increased, while CP significantly decreased. As shoal size area (OA) did not change, these data indicate that after adaptation animals were less uniformly distributed in the shoal and tended to spend most of their time in the upper part of the tank. Shoal cohesion of zebrafish fed with probiotics (PROBIO) was significantly different from controls (CTRL) (Fig. 1). Although AD was not significantly different between the two groups for treatment, DV (Finteraction = 17.79, p < 0.0001) and ND (Ftreatment = 7.948, p = 0.0059) showed significant dependence on treatment and time (Table S1). OA, was also significantly increased in the PROBIO group (Finteraction = 4.423, p = 0.0483). The two groups showed completely different preferences for location in the water column (CP) (Finteraction = 377.8, p < 0.0001) (Table S1). More importantly, SF (Finteraction = 7.862, p = 0.0127) was strongly increased in the PROBIO group. Together, these data suggest that feeding of the probiotic L. rhamnosus IMC 501 significantly alters social and explorative behaviour in zebrafish (Video S1).

Probiotic treatment alters zebrafish shoaling behaviour.
Average Distance (AD), Distance Variance (DV), Nearest Distance (ND), Occupied Area (OA), Column Preference (CP) and Shuttling Frequency (SF) measurements (Supplementary Figure 1) at the beginning (T0) and end of 4 weeks (T4) of probiotic treatment, are shown from both control (CTRL) and treated (PROBIO) animals. Bars represent SEM. #indicates significant difference from T0 (Two-way ANOVA with Sidak’s post-hoc test, p < 0.05); asterisks indicate statistical significant difference between the CTRL and PROBIO groups (Two-way ANOVA with Sidak’s post-hoc test: ***p < 0.001, **p < 0.01, *p < 0.05). DV Finteraction = 17.79, p < 0.0001; ND Ftreatment = 7.948, p = 0.0059; OA Finteraction = 4.423, p = 0.0483; CP Finteraction = 377.8, p < 0.0001; SF Finteraction = 7.862, p = 0.0127.
Probiotic administration modulates the gut microbial communities
The α-diversity indices Chao1 (138.02 ± 40.36 CTRL, 133.25 ± 30.00 PROBIO) and Shannon (4.12 ± 0.59 CTRL, 3.92 ± 0.97 PROBIO) of the gut microbiota in CTRL and PROBIO zebrafish were not significantly different, indicating that the dietary administration of L. rhamnosus does not affect overall richness of the zebrafish gut bacterial community. Similarly, weighted two-dimensional Principal Coordinates Analysis showed that PROBIO samples largely clustered with CTRL samples, demonstrating overall similarity of the gut bacterial communities between the two groups (Fig. 2A). However, we noticed differences at the phylum and class levels (Fig. 2B,C). Fusobacteria dominated both groups, accounting for 75.83% (CTRL) and 61.18% (PROBIO) of the OTUs retrieved. Proteobacteria (17.24% CTRL, 9.01% PROBIO), Firmicutes (4.10% CTRL, 28.75% PROBIO) and Cyanobacteria (0.97% CTRL, 0.19% PROBIO) were also present in both groups, yet at different abundances. Actinobacteria were present only in the CTRL (1,86%) and Bacteroidetes (0.78%) only in the PROBIO samples. The abundance of Lactobacillus (8,3%) in the PROBIO group is likely representative of the dietary administration of L. rhamnosus (Phylum: Firmicutes, Class: Bacilli), suggesting that the fed probiotic bacteria colonised the gut, in agreement with previous studies45,46,47. In addition, in the PROBIO group there is a significant increase of the Streptococcus genus (t-test p value 0.025, Table S2; Fig. 2D). The abundance of both Lactobacillus and Streptococcus might be related to the symbiotic relationship between L. rhamnosus and Streptococcus thermophilus described in zebrafish after probiotic administration44,46,47. On average, Proteobacteria, that include potentially pathogenic genera (e.g. Vibrio and Plesiomonas, Fig. 2D), were less represented in the PROBIO group (9.01%) than in the CTRL (17.84%), although this difference was not statistically significant.

Whole intestine bacterial community analysis of adult zebrafish after probiotic treatment.
(A) Principal coordinates analysis (PCoA) plots using weighted UnicFrac distances. Stacked bar chart representing the relative abundance of bacterial phylum (B) and classes (C). (D) Fold change of the genus Lactobacillus, Streptococcus, Plesiomonas and Vibrio in the PROBIO relative to the CTRL group.
Probiotic feeding induces changes in neuronal gene expression in brain and gut
Previous studies on the gut-brain axis in rodent models have repeatedly shown altered expression of bdnf in certain brain regions under various conditions7,8,14,15,17. In addition, BDNF signalling is known to be involved in social behaviour48,49. As such, the expression of bdnf in the brains of rodent models is very plastic and regulated in a dynamic way. A very well-studied molecular pathway resulting in elevated levels of bdnf mRNA is the CREB (cAMP response element binding protein) signalling pathway that gets activated upon neuronal excitation. Many behavioural paradigms have been shown to induce bdnf expression in various brain regions including novelty exposure (e.g. social novelty, novel environment, environmental enrichment) and learning tasks. On the other hand, a reduction in bdnf levels is often a signature of reduced cognitive abilities. Importantly, reduced levels of bdnf have been associated with depression and it has been suggested that conventional antidepressants and ketamine mediate their antidepressant-like effects by increasing bdnf in particular in the hippocampus, making it an essential determinant of antidepressant efficacy50. We therefore determined if the expression of bdnf was altered after probiotic administration in zebrafish. In the brain, the PROBIO group showed a significant (near two-fold) increase of bdnf expression compared to the CTRL group (t-test p value 0.001) (Fig. 3A). In contrast, we did not detect any significant difference between the PROBIO and CTRL groups in the gut, although there was a trend towards decreased expression.

Probiotic feeding induces changes in neuronal gene expression in brain and gut.
Relative expression of the D. rerio bdnf (A) and tph1a, tph1b, tph2, htr1aa, slc6a4a, mao (B) genes in brain and gut of zebrafish treated with L. rhamnosus (PROBIO) and controls (CTRL). The mean relative expression levels of the PROBIO group are normalized to the mean relative expression of the CTRL group. AU, arbitrary units. Bars represent SEM. Asterisks indicate statistical significant difference between the CTRL and PROBIO groups: **t-test p < 0.01, *t-test p < 0.05.
Furthermore, socio-emotional behaviour in vertebrates is heavily dependent on the serotonergic system51,52. Moreover, microbial influence on the serotonergic system has been proposed as a key mediator of gut-brain signalling16,33. Therefore, we next investigated potential changes in expression of selected genes involved in serotonin signalling and metabolism (tph1a, tph1b, tph2, htr1aa, slc6a4a and mao), both in the gut and in the brain of zebrafish fed with L. rhamnosus IMC 501 (Fig. 3B). In the brain, five genes showed a statistically significant increase of the relative expression levels in the PROBIO group (t-test p values: tph1a 0.03, tph2 0.01, htr1aa 0.01, slc6a4a 0.004 and mao 0.0003), indicating that the probiotic administration changes the expression levels of genes involved in the serotonergic system. In contrast, in the gut, we detected a reduction of the expression levels for the genes tph1a (t-test p value 0.001), tph1b (p = 0.001) and htr1aa (p = 0.03), while the other genes did not show significant differences between the PROBIO and CTRL groups. Together, these data suggest tissue-specific modulation of the serotonergic system.
Discussion
Zebrafish is an increasingly important model organism in neuroscientific basic and translational research53. Zebrafish digestive tract is similar to that of mammals in its development, organisation and function and zebrafish are well suited for studying host-microbe interactions as they have innate and adaptive immune systems similar to higher vertebrates54. Surprisingly, to our knowledge, there have been no studies yet investigating the influence of beneficial or commensal gut microbes on the gut-brain axis in this model organism. Yet, host-microbe interactions in the gut are now widely accepted to affect brain function and behaviour and considered as a potential target in psychiatric diseases55. We have therefore decided to explore the effects of feeding healthy, wild-type zebrafish with the probiotic strain L. rhamnosus IMC 501 on shoaling behaviour, a measure of basal exploratory and social behaviour.
In our study the shoaling pattern of zebrafish differed significantly from controls after 4 weeks of probiotic treatment. In particular, the probiotic treated group was more uniform and showed more homogeneity within the shoal as evidenced by a decreased average distance variance (DV). In addition, the nearest distance (ND) and occupied shoal area (OA) were significantly increased after treatment. Concerning the water column preference (CP), the PROBIO group showed a completely different behavioural pattern. While the control group changed significantly to prefer the upper parts of the water tank at the end of the 4 weeks of observation, which is likely due to habituation to the aquarium, the probiotic treated group continued to prefer the medium/deeper part of the tank. Furthermore, a significant increase in shuttling frequency (SF) indicates a strong increase in locomotor activity. These data indicate an increased exploratory behaviour in the PROBIO animals56. The different behaviour between CTRL and PROBIO group suggests a higher exploration capability and attention/alert in the PROBIO group.
Notably, the detected pronounced changes in behaviour were accompanied by strongly increased expression levels of bdnf and several genes involved in serotonin signalling metabolism in the brain, while their expression in the gut was reduced. In fact, BDNF and the serotonergic system in the brain work synergistically and influence each other mutually on the transcriptional level, via epigenetic mechanisms and by direct protective and regenerative effects of BDNF on serotonergic neurons throughout development and in later life57.
Most well established, the serotonergic system is linked to many behavioural traits58. As such, selective serotonin reuptake-inhibitors (SSRIs) are the first-line treatment in severe depression and also recommended to treat generalized anxiety disorder59. Increased serotonin production and receptor expression in the zebrafish brain is therefore perfectly in line with the observed behavioural change induced by L. rhamnosus administration. In fact, both, the serotonergic system as well as BDNF levels, are well established effectors of the microbiota in the host. As such, Neufeld et al.60 found decreased expression of the serotonin receptor 1a (Htr1a) in germ-free mice60. In addition, a recent study showed increased levels of serotonin in the hippocampus of germ-free mice16, likely due to increased tryptophan uptake in the absence of tryptophan-metabolising microorganisms in the gut16. This study also revealed altered BDNF levels, which has now commonly been found in animal models with altered microbiota7,8,14,16. Moreover, the microbiota is directly involved in stimulating serotonin release from enterochromaffin cells in the gut epithelium, regulating enteric nervous system and vagus nerve activity33. Thus, our findings support existing literature connecting the microbiota with these systems also in the zebrafish.
Of note, we found different expression of bdnf and serotonin-related genes in the gut and the brain.
Recent findings show that BDNF plays an important role in gastrointestinal function and an increased level of BDNF in the colon of diarrhea-predominant irritable bowel syndrome (IBS) patients with abdominal pain was described61. However, the regulation of the expression of bdnf in gut is still unclear. Additionally, serotonin release in the gut and the enteric nervous system is regulated independently of regulation in the CNS62. It is therefore likely that L. rhamnosus supplementation has independent effects on these two systems, which does not preclude the possibility that both systems directly or indirectly contribute to the observed change in shoaling behaviour.
Analysis of the microbiota composition revealed that Proteobacteria, Fusobacteria and Firmicutes appeared consistently in the gut microbiota of zebrafish. Proteobacteria and Fusobacteria are common members of the gut microbiota in adult zebrafish and are especially well adapted to conditions of the fish gastro-intestinal tract in a surrounding aquatic environment63. The Firmicutes phylum (including Streptococcus spp. and Lactobacillus spp.) increased significantly in PROBIO group (t-test p value 0.038, Table S2) at the expense of representatives of the Proteobacteria fraction (including Vibrio spp. and Plesiomonas spp.) (Fig. 2D). This result is in agreement with previous studies reporting increased abundance of Lactobacillus and Streptococcus in zebrafish after probiotic administration, resulting in several beneficial effects44,46,47. Interestingly, a higher proportion of Proteobacteria, together with a lower abundance of Firmicutes, characterizes the intestinal microbiota dysbiosis in zebrafish model with inflammatory bowel disease (IBD)-like colitis and correlates significantly with enterocolitis severity, which mirrors changes in the human gut microbiota in IBD64.
Conclusion
So far most findings linking the microbiota with virtually all of the host physiological processes have originated from rodent studies. Compelling and rapidly accumulating evidence highlights the strong impact of the presence and activity of certain microbes in the gut on host physiology and behaviour. Notably, many studies have made use of animals raised in a sterile, germ-free environment. Using this approach it was found that the microbiota is critical for the host’s serotonergic system and tuning the stress response at a behavioural and endocrine level, which has further implications for our perception of stress-related psychiatric and gastrointestinal diseases. While these findings are stimulating, it has to be acknowledged that germ-free animals represent a highly artificial model that is uniquely useful to determine physiological processes influenced by the microbiota in a black-or-white manner, rather than a translatable model for human disease. Taken together, our data reveal specific, so far undescribed behavioural and neurochemical changes in healthy zebrafish, induced by alteration of the enteric microbiota using a potential psychobiotic bacterium (L. rhamnosus IMC 501), demonstrating psychotropic effects on the behavioural and molecular level. We may assert that the bidirectional signalling between microbiota and brain exists also in zebrafish, consolidating it as a valuable animal model for translational medicine.
While our data highlight the fact that supplementation with a probiotic is sufficient to induce sizeable effects on at least three levels of readout, the microbiota, neurochemistry and behaviour, further experimentation is necessary to connect these domains in a causal manner. Another limitation of this study, as in most other studies investigating gut-brain communication, is incomplete mechanistic insight into the routes connecting the increased abundance of the probiotic with altered neurochemistry and behaviour. In addition to modulation of the immune system via secreted metabolites and microbial cell wall material, especially differential stimulation of the vagus nerve have been shown to be among the routes of communication that mediates, for example, the antidepressant-like effects of L. rhamnosus in mammals11.
A deeper understanding of these communication mechanisms will be crucially important for the development of any microbiota-based therapeutic strategy for psychiatric diseases in human and animal health and for the discovery of alternative pathways and substrates to treat brain disorders that do not respond to available drugs.
Methods
Ethics Statement
All fish were treated in accordance with the Directive of the European Parliament and of the Council on the Protection of Animals Used for Scientific Purposes (directive 2010/63/EU) and in agreement with the Bioethical Committee of University of Napoli Federico II. All experiments involving fish were approved by the Bioethical Committee of the University of Naples Federico II (authorization protocol number 47339-2013). Following behavioural testing, the animals were euthanized by immersion in overdose 500 mg/ L-1 of 3-aminobenzoic acid ethyl ester (MS-222) buffered to pH 7.4 (Sigma–Aldrich, USA).
Animals and husbandry
Adult 4–6-month-old male and female zebrafish of heterozygous “wild type” strain were obtained from local commercial distributors (Carmar, Napoli, Italy). All fish were given at least 14 days to acclimate to the laboratory environment and housed in groups of 12 fishes per 30-L tank. All tanks were filled with deionized water before introducing fish. Fish were fed two times daily with sterilized commercial food (Sera Vipagran, Germany). The room and water temperatures were maintained at 25–27 °C. Illumination (1010 ± 88 lx) was provided by ceiling-mounted fluorescent light tubes on a 14-h cycle (D:N = 14h:10h) consistent with the standards of zebrafish care65. All fish used in this study were experimentally naïve. Two experimental groups were evaluated: a control group (CTRL, n = 12) and a probiotic-treated group (PROBIO, n = 12). The experiment was repeated twice to obtain biological duplicates.
Probiotic administration
The control group (CTRL) was fed twice per day with the commercial diet only and the probiotic-treated group (PROBIO) was fed twice per day with the commercial diet and twice per day with the lyophilized probiotic strain L. rhamnosus IMC 501, provided by Synbiotec (Camerino, Italy) via rearing water at a final concentration of 106 colony-forming units/g (0,01 g/l), according to the manufactory suggestions, for 28 days. Before the administration of the lyophilized L. rhamnosus IMC 501, a viability test was performed (see Supplementary information, Supplementary methods).
Behavioral testing
Fish swimming behaviour was video-recorded on the first day of probiotic treatment (T0) and at the end of treatment at 28 days (T4). Recording was performed always at the same hour (between 14:00 and 15:00) with a Nikon D7000 camera for 6 min, acquired and analysed with 2D video tracking analysis and modelling tool (Tracker, California, USA) (www.cabrillo.edu/~dbrown/tracker/). In line with Blaser and Gerlai66 and Gerlai40, the video tracking data were used to determine the following behavioural measures: Average Distance (AD), Distance Variance (DV), Nearest Distance (ND), Occupied Area (or Shoal size area) (OA), Water Column Position (CP) and Shuttling Frequency (SF) (Supplementary Figure 1). In particular, AD represents the inter-individual distance and defines and calculates the average of all distances between a fish and its shoal members. Each fish thus will get an AD value and this value is calculated for all fish within a shoal and for any given moment of time sampled. As it takes the position of all fish in the shoal, AD is a powerful measure of shoal cohesion40. DV is the AD variance for each fish. Notably, this variability depends on the relative position of each fish within a shoal, so that its mean value, when calculated for the entire shoal, represents the homogeneity of the distribution of fish within that shoal. The less uniformly the shoal members are distributed, the larger the variance will be. ND calculates the nearest distance of each single fish to its closest neighbour and represents a measure of the shoal cohesion less powerful than AD, but independent from the shoal size40. OA, which also measure shoal cohesion, calculates the occupied area of all animals in a temporal unit on a two-dimensional plane. CP represents the water column position (surface = 0) and indicates the deep preference of shoaling animals in the tank56. SF, which measures the locomotor activity of animals in the shoal66, is the total number of times that the fish entered the left, the right, the upper and the lower halves of the tank. For AD, DV and ND measurements each fish was tracked on six frames randomly selected from each video. For SF measurements, five 10 sec intervals randomly selected from each video were analysed. For OA and CP analysis, ImageJ 1.49 software (National Institute of Mental Health, Bethesda, Maryland, USA) was used to select the areas occupied and the deep position of each fish in the water column in all video collected. Here, ten images were randomly selected from each video, one every twenty seconds.
Behaviour was evaluated by two independent, trained observers (intra-rater reliability > 0.90). Results are reported as means ± SEM. Two-way ANOVA with Sidak’s post-hoc test was used to evaluate the significance of the effect of the probiotic treatment, using GraphPad Prism version 6.00 for Windows (GraphPad Software, La Jolla California USA, www.graphpad.com).
Gut microbiota analysis
Fish were dissected, brain and gut were entirely excised and stored in RNAlater (Ambion, Carlsbad CA, USA) at −80 °C. The detailed description of the aseptic dissection procedure is reported in the Supplementary information (Supplementary methods).
After 30 min lysozyme treatment (50 mg/ml) at 37 °C, DNA was extracted from zebrafish gut (60 mg) using the QIAamp Stool Mini Kit (Qiagen, West Sussex, UK) with minor modifications44 and quantified using a Nanodrop TM-1000 Spectrophotometer (Thermo Scientific Ltd, DE, USA).
PCR was conducted to amplify the V3 region of the 16S rRNA gene using the primer pair P3-P267 and the V1-V2 region using the primer pair 27F-338RI/338RII63 (Table S2). PCR reactions consisted of 25 μl ReadyMix Taq PCR Reaction Mix with MgCl2 (Sigma-Aldrich), 50 pmol of each of primer, 70 ng of DNA in a final volume of 50 μl. Touchdown thermal cycling was conducted using a GeneAmp PCR System 9700 (Perkin-Elmer, CA, USA), applying the conditions previously described67.
PCR products (PROBIO n = 5, CTRL n = 5 for each experiment, resulting in a total of ten samples for each group, PROBIO and CTRL, respectively) were purified using the QIAquick PCR Purification Kit (Qiagen). Libraries for sequencing were prepared using the Ion Plus Fragment Library Kit (Life Technologies, USA) and quantified using the Ion Library Quantitation Kit (Life Technologies) and concentrations were adjusted to 26 pM. The emulsion PCR was carried out using the Ion PGM Template OT2 400 Template Kit (Life Technologies) according to the manufacturer’s instructions. Multiplexed sequencing was carried out on a 318 chip using the Ion Torrent PGM system and employing the Ion Sequencing 400 kit (Life Technologies) according to the supplier’s instructions. After sequencing, the sequence reads were filtered and trimmed by the PGM software to remove low quality and polyclonal sequences and quality control was performed using Quantitative Insights Into Microbial Ecology v1.8.0 (QIIME) (quality score >25). In order to calculate downstream diversity measures (alpha and beta diversity indices, UniFrac analysis), 16S rRNA Operational Taxonomic Units (OTUs) were defined at >97% sequence identity. All reads were classified to the lowest possible taxonomic rank using QIIME and a reference dataset from the Ribosomal Database Project. OTUs were assigned using Uclust. The hierarchical clustering based on population profiles of most common and abundant taxa was performed using UPGMA clustering on the distance matrix of OTU abundance. Ecological metrics were calculated on rarefied OTU tables with QIIME to assess sampling depth coverage using observed species, Chao1, Shannon’s diversity index and Good’s coverage. QIIME was also used to calculate Beta diversity metrics among samples using weighted Unifrac distances22 and Bray-Curtis similarity44. The distance matrix was represented by a dimensional principal coordinates analysis (PCoA) plots. Taxonomy of intestinal bacterial communities data are presented as means ± standard error of the mean (SEM). Statistical analysis was performed using the two-tailed t test.
RNA extraction and expression analysis
Total RNA was separately extracted from brain and gut samples using the Pure Link RNA Mini Kit (Ambion, Carlsbad CA, USA) and DNase treatment was performed using the Pure Link DNase Set (Ambion). RNA was quantified using a Nanodrop ND-2000 Spectrophotometer (Thermo Scientific). Total RNA was reverse transcribed using the SuperScript VILO cDNA Synthesis Kit (Ambion) and an oligo dT primer. Expression levels of the Danio rerio genes bdnf, tph1a, tph1b, tph2, htr1aa, slc6a4a and mao in gut and brain were evaluated by Real Time PCR. PCR amplifications were conducted on 30 ng of first-strand cDNA using the primer pairs listed in Table S3 and the Power SYBR Green Master mix (Ambion). The reactions were conducted in technical triplicates and biological duplicates and were run in the 7500 ABI Thermal Cycler (Applied Biosystems) using the thermal cycle: 50 °C, 2 min; 95 °C, 10 min; 40 cycles of 95 °C, 15 sec and 60 °C, 1 min, followed by a melting curve cycle.
The Real-Time PCR Miner online tool68 was used to calculate the PCR efficiency (E) and optimal threshold cycle (CT) for each well. The mean relative expression ratio and standard error of the target genes was calculated using the β-actin gene as the endogenous control applying the formula (1 + E target)–CT target/(1 + E control)−CT control.
Statistical significance of differences in expression levels of the target genes between CTRL and PROBIO groups was assessed by two-tailed t-tests.
Additional Information
How to cite this article: Borrelli, L. et al. Probiotic modulation of the microbiota-gut-brain axis and behaviour in zebrafish. Sci. Rep. 6, 30046; doi: 10.1038/srep30046 (2016).
References
O’Sullivan, E. et al. BDNF expression in the hippocampus of maternally separated rats: does Bifidobacterium breve 6330 alter BDNF levels? Benef Microbes 2, 199–207 (2011).
Al-Asmakh, M., Anuar, F., Zadjali, F., Rafter, J. & Pettersson, S. Gut microbial communities modulating brain development and function. Gut Microbes 3, 366–373 (2012).
Cryan, J. F. & Dinan, T. G. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci 13, 701–712 (2012).
Stilling, R. M., Dinan, T. G. & Cryan, J. F. Microbial genes, brain & behaviour - epigenetic regulation of the gut-brain axis. Genes Brain Behav 13, 69–86 (2014).
Dinan, T. G., Stilling, R. M., Stanton, C. & Cryan, J. F. Collective unconscious: how gut microbes shape human behavior. J Psychiatr Res 63, 1–9 (2015).
Dinan, T. G., Stanton, C. & Cryan, J. F. Psychobiotics: a novel class of psychotropic. Biol Psychiatry 74, 720–726 (2013).
Sudo, N. et al. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J Physiol 558, 263–275 (2004).
Bercik, P. et al. Chronic gastrointestinal inflammation induces anxiety-like behavior and alters central nervous system biochemistry in mice. Gastroenterology 139, 2102–2112 e2101 (2010).
Messaoudi, M. et al. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br J Nutr 105, 755–764 (2011).
Messaoudi, M. et al. Beneficial psychological effects of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in healthy human volunteers. Gut Microbes 2, 256–261 (2011).
Bravo, J. A. et al. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc Natl Acad Sci USA 108, 16050–16055 (2011).
Partty, A., Kalliomaki, M., Wacklin, P., Salminen, S. & Isolauri, E. A possible link between early probiotic intervention and the risk of neuropsychiatric disorders later in childhood: a randomized trial. Pediatr Res 77, 823–828 (2015).
Divyashri, G., Krishna, G., Muralidhara & Prapulla, S. G. Probiotic attributes, antioxidant, anti-inflammatory and neuromodulatory effects of Enterococcus faecium CFR 3003: in vitro and in vivo evidence. J Med Microbiol 64, 1527–1540 (2015).
Gareau, M. G. et al. Bacterial infection causes stress-induced memory dysfunction in mice. Gut 60, 307–317 (2011).
Diaz Heijtz, R. et al. Normal gut microbiota modulates brain development and behavior. Proc Natl Acad Sci USA 108, 3047–3052 (2011).
Clarke, G. et al. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol Psychiatry 18, 666–673 (2013).
Stilling, R. M. et al. Microbes & neurodevelopment–Absence of microbiota during early life increases activity-related transcriptional pathways in the amygdala. Brain Behav Immun 50, 209–220 (2015).
Gotz, R., Raulf, F. & Schartl, M. Brain-derived neurotrophic factor is more highly conserved in structure and function than nerve growth factor during vertebrate evolution. J Neurochem 59, 432–442 (1992).
Thoenen, H. Neurotrophins and neuronal plasticity. Science 270, 593–598 (1995).
Berninger, B., Schinder, A. F. & Poo, M. M. Synaptic reliability correlates with reduced susceptibility to synaptic potentiation by brain-derived neurotrophic factor. Learn Mem 6, 232–242 (1999).
Lanave, C., Colangelo, A. M., Saccone, C. & Alberghina, L. Molecular evolution of the neurotrophin family members and their Trk receptors. Gene 394, 1–12 (2007).
D’Angelo, L. et al. Brain-derived neurotrophic factor: mRNA expression and protein distribution in the brain of the teleost Nothobranchius furzeri. J Comp Neurol 522, 1004–1030 (2014).
Vissio, P. G., Canepa, M. M. & Maggese, M. C. Brain-derived neurotrophic factor (BDNF)-like immunoreactivity localization in the retina and brain of Cichlasoma dimerus (Teleostei, Perciformes). Tissue Cell 40, 261–270 (2008).
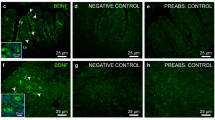
De Felice, E. et al. Localization of BDNF expression in the developing brain of zebrafish. J Anat 224, 564–574 (2014).
Timmusk, T. et al. Multiple promoters direct tissue-specific expression of the rat BDNF gene. Neuron 10, 475–489 (1993).
Hayes, V. Y., Towner, M. D. & Isackson, P. J. Organization, sequence and functional analysis of a mouse BDNF promoter. Brain Res Mol Brain Res 45, 189–198 (1997).
Liu, Q. R. et al. Rodent BDNF genes, novel promoters, novel splice variants and regulation by cocaine. Brain Res 1067, 1–12 (2006).
Heinrich, G. & Pagtakhan, C. J. Both 5′ and 3′ flanks regulate Zebrafish brain-derived neurotrophic factor gene expression. BMC Neurosci 5, 19 (2004).
Germana, A. et al. Expression of brain-derived neurotrophic factor and TrkB in the lateral line system of zebrafish during development. Cell Mol Neurobiol 30, 787–793 (2010).
Ventriglia, M. et al. Serum brain-derived neurotrophic factor levels in different neurological diseases. Biomed Res Int 2013, 901082 (2013).
Mondelli, V. et al. Stress and inflammation reduce brain-derived neurotrophic factor expression in first-episode psychosis: a pathway to smaller hippocampal volume. J Clin Psychiatry 72, 1677–1684 (2011).
Clarke, G. et al. Minireview: Gut microbiota: the neglected endocrine organ. Mol Endocrinol 28, 1221–1238 (2014).
Yano, J. M. et al. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 161, 264–276 (2015).
Norton, W. H., Folchert, A. & Bally-Cuif, L. Comparative analysis of serotonin receptor (HTR1A/HTR1B families) and transporter (slc6a4a/b) gene expression in the zebrafish brain. J Comp Neurol 511, 521–542 (2008).
Lillesaar, C. The serotonergic system in fish. J Chem Neuroanat 41, 294–308 (2011).
O’Mahony, S. M., Clarke, G., Borre, Y. E., Dinan, T. G. & Cryan, J. F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav Brain Res 277, 32–48 (2015).
Maximino, C. et al. in Serotonin: Biosynthesis, Regulation and Health Implications (ed S. Hall ) Ch. X, (Nova Science, 2013).
Kalueff, A. V., Echevarria, D. J. & Stewart, A. M. Gaining translational momentum: more zebrafish models for neuroscience research. Prog Neuropsychopharmacol Biol Psychiatry 55, 1–6 (2014).
Stewart, A. M. et al. Perspectives on experimental models of serotonin syndrome in zebrafish. Neurochem Int 62, 893–902 (2013).
Gerlai, R. Social behavior of zebrafish: from synthetic images to biological mechanisms of shoaling. J Neurosci Methods 234, 59–65 (2014).
Engeszer, R. E., Patterson, L. B., Rao, A. A. & Parichy, D. M. Zebrafish in the wild: a review of natural history and new notes from the field. Zebrafish 4, 21–40 (2007).
Buske, C. & Gerlai, R. Shoaling develops with age in Zebrafish (Danio rerio). Prog Neuropsychopharmacol Biol Psychiatry 35, 1409–1415 (2011).
Saverino, C. & Gerlai, R. The social zebrafish: behavioral responses to conspecific, heterospecific and computer animated fish. Behav Brain Res 191, 77–87 (2008).
Falcinelli, S. et al. Lactobacillus rhamnosus lowers zebrafish lipid content by changing gut microbiota and host transcription of genes involved in lipid metabolism. Sci Rep 5, 9336 (2015).
Avella, M. A., Olivotto, I., Silvi, S., Place, A. R. & Carnevali, O. Effect of dietary probiotics on clownfish: a molecular approach to define how lactic acid bacteria modulate development in a marine fish. Am J Physiol Regul Integr Comp Physiol 298, R359–371 (2010).
Gioacchini, G. et al. Probiotics can induce follicle maturational competence: the Danio rerio case. Biol Reprod 86, 65 (2012).
Gioacchini, G. et al. The influence of probiotics on zebrafish Danio rerio innate immunity and hepatic stress. Zebrafish 11, 98–106 (2014).
Desbonnet, L. et al. Gut microbiota depletion from early adolescence in mice: Implications for brain and behaviour. Brain Behav Immun 48, 165–173 (2015).
Branchi, I. The mouse communal nest: investigating the epigenetic influences of the early social environment on brain and behavior development. Neurosci Biobehav Rev 33, 551–559 (2009).
Bjorkholm, C. & Monteggia, L. M. BDNF-a key transducer of antidepressant effects. Neuropharmacology 102, 72–79 (2016).
Kiser, D., Steemers, B., Branchi, I. & Homberg, J. R. The reciprocal interaction between serotonin and social behaviour. Neurosci Biobehav Rev 36, 786–798 (2012).
Shams, S., Chatterjee, D. & Gerlai, R. Chronic social isolation affects thigmotaxis and whole-brain serotonin levels in adult zebrafish. Behav Brain Res 292, 283–287 (2015).
Fonseka, T. M., Wen, X. Y., Foster, J. A. & Kennedy, S. H. Zebrafish models of major depressive disorders. J Neurosci Res 94, 3–14 (2016).
Goldsmith, J. R. & Jobin, C. Think small: zebrafish as a model system of human pathology. J Biomed Biotechnol 2012, 817341 (2012).
Mayer, E. A., Knight, R., Mazmanian, S. K., Cryan, J. F. & Tillisch, K. Gut microbes and the brain: paradigm shift in neuroscience. J Neurosci 34, 15490–15496 (2014).
Blaser, R. E. & Goldsteinholm, K. Depth preference in zebrafish, Danio rerio: control by surface and substrate cues. Anim Behav 83, 953–959 (2012).
Homberg, J. R., Molteni, R., Calabrese, F. & Riva, M. A. The serotonin-BDNF duo: Developmental implications for the vulnerability to psychopathology. Neurosci Biobehav R 43, 35–47 (2014).
Olivier, B. Serotonin: a never-ending story. Eur J Pharmacol 753, 2–18 (2015).
Fournier, J. C. et al. Antidepressant drug effects and depression severity: a patient-level meta-analysis. JAMA 303, 47–53 (2010).
Neufeld, K. M., Kang, N., Bienenstock, J. & Foster, J. A. Reduced anxiety-like behavior and central neurochemical change in germ-free mice. Neurogastroenterol Motil 23, 255–264, e119 (2011).
Wang, P. et al. Increased production of BDNF in colonic epithelial cells induced by fecal supernatants from diarrheic IBS patients. Sci Rep-Uk 5, 10121 (2015).
Racke, K. & Schworer, H. Regulation of Serotonin Release from the Intestinal-Mucosa. Pharmacol Res 23, 13–25 (1991).
Roeselers, G. et al. Evidence for a core gut microbiota in the zebrafish. ISME J 5, 1595–1608 (2011).
He, Q. et al. Microbial fingerprinting detects intestinal microbiota dysbiosis in Zebrafish models with chemically-induced enterocolitis. BMC Microbiol 13, 289 (2013).
Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio Rerio). 4th edn, (University of Oregon Press, 2000).
Blaser, R. & Gerlai, R. Behavioral phenotyping in zebrafish: Comparison of three behavioral quantification methods. Behav Res Methods 38, 456–469 (2006).
Muyzer, G., de Waal, E. C. & Uitterlinden, A. G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol 59, 695–700 (1993).
Zhao, S. & Fernald, R. D. Comprehensive algorithm for quantitative real-time polymerase chain reaction. J Comput Biol 12, 1047–1064 (2005).
Acknowledgements
The authors are grateful to Dr. Daniel L. Merrifield and the Aquatic Animal Nutrition and Health. Research Group, School of Biological Sciences, Plymouth University, UK, for conducting the Ion Torrent NGS analysis. The authors are also grateful to Dr. Livia D’Angelo for scientific support in BDNF analysis. This work was supported by the grant D. G. 1 P.O. CAMPANIA FSE 2007/2013.
Author information
Authors and Affiliations
Contributions
L.B. designed and performed the experiments, analyzed the data and wrote the manuscript. S.A. analyzed the data and wrote the manuscript. S.D.P., L.D. and L.F.M. performed the experiments. C.A. participated to the data analysis. R.M.S., T.G.D., J.F.C. and A.F. contributed to the interpretation of the data and critically read the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Borrelli, L., Aceto, S., Agnisola, C. et al. Probiotic modulation of the microbiota-gut-brain axis and behaviour in zebrafish. Sci Rep 6, 30046 (2016). https://doi.org/10.1038/srep30046
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep30046
This article is cited by
-
Vitamin B12 produced by Cetobacterium somerae improves host resistance against pathogen infection through strengthening the interactions within gut microbiota
Microbiome (2023)
-
Garlic (Allium sativum) as a dietary ingredient can cause dysbiosis in the microbiota of the California yellowtail (Seriola dorsalis) at high concentrations
Aquaculture International (2023)
-
Interactive Effect of Dietary Heat-Killed Lactobacillus Plantarum L-137 and Berberine Supplementation on Intestinal Mucosa and Microbiota of Juvenile Black Sea Bream (Acanthopagrus Schlegelii)
Probiotics and Antimicrobial Proteins (2023)
-
Lactobacillus rhamnosus GG treatment potentiates ethanol-induced behavioral changes through modulation of intestinal epithelium in Danio rerio
International Microbiology (2023)
-
Zebrafish: an efficient vertebrate model for understanding role of gut microbiota
Molecular Medicine (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
