
Abstract
Grazing exclusion (GE) has been widely considered as an effective avenue for restoring degraded grasslands throughout the world. GE, via modifying abiotic and biotic environments, inevitably affects phenological development. A five-year manipulative experiment was conducted in a Tibetan alpine meadow to examine the effects of GE on phenological processes and reproductive success. The study indicated that GE strongly affected phenological development of alpine plant species. Specifically, the low-growing, shallow-rooted species (LSS), such as Kobresia pygmaea, are more sensitive to GE-caused changes on upper-soil moisture and light. GE advanced each phonological process of K. pygmaea, except in the case of the treatment of fencing for 5 years (F5), which postponed the reproductive stage and lowered the reproductive success of K. pygmaea. Increased soil moisture triggered by GE, especially in the upper soil, may stimulate growth of LSS. However, the thick litter layer under the F5 treatment can influence the photoperiod of LSS, resulting in suppression of its reproductive development. These findings indicate that plant traits associated with resource acquisition, such as rooting depth and plant height, mediate plant phenology and reproductive responses to grazing exclusion treatments.
Similar content being viewed by others


Introduction
Grasslands cover approximately 40% of the earth’s terrestrial surface. They play a significant role in regulating and feedback to climates1,2. Globally, grazing by livestock is one of the most common human disturbances to grasslands. Overgrazing can cause severe grassland degradation3 and reduce both productivity and resilience4,5. After extensive practices, grazing exclusion (GE) has gradually been recognized as a simple, effective method for restoring degraded grasslands throughout the world6.
Recent research has focused on the effects of GE on vegetation succession, plant diversity, community structure and productivity6,7 and soil biogeochemical processes8,9. Vegetation development processes and reproductive success rates, as important aspects of plant phenology, have profound implications for ecological functions10. For example, the unfolding and coloring of leaves are processes closely related to the length of the vegetation growing season (GSL)11, which can substantially affect ecosystem processes such as carbon and water cycling12,13. To date, few studies have examined the effects of GE on plant phenology.
Abiotic environmental factors, such as temperature14, water (precipitation and soil moisture)15, photoperiod16 and soil nutrients17, combined with the biotic factors, such as plant life history15, influence phenological processes and reproductive success18. GE may alter phenology indirectly by modifying microclimates, such as soil nutrients19, soil moisture20, soil temperature21 and photoperiod22. For example, by encouraging a thicker litter layer, GE would decrease water loss through soil evaporation23 and possibly advance vegetation green-up. Although changes in abiotic factors can lead to significant consequences for phenological development, there is still a severe shortage of information on responses of phenological processes and reproductive success to GE, especially for the alpine grassland ecosystem.
Grasslands cover approximately 40% of the land in China24. Over the past 20 years, the Chinese government has invested heavily in the restoration of degraded grasslands. i For example, the ‘Start-up Re-grass Program’ on the Tibetan Plateau25 has been in place since 2004 to protect grasslands from heavy grazing through GE. Previous research in this region has focused on the impacts of GE on vegetation restoration and soil properties26,27. To enhance our understanding of the efficiency of GE treatments, knowledge of its effects on plant phenology are necessary. In order to meet this need, a five-year manipulative experiment was conducted in a Tibetan alpine meadow to examine the effects of GE treatment on phonological processes and reproductive success. Three major questions were addressed: (i) How does GE influence phenological timing and duration? (ii) What impacts does GE exert on plant reproductive success? (iii) What are the physiological mechanisms underlying the above two processes?
Materials and Methods
Study area
This experiment was conducted in a typical alpine meadow grassland at Naqu, northern Tibet, China (31°38.513′ N, 92°0.921′ E), at an approximate elevation of 4600 m. The mean annual temperature is −1.2 °C. The mean annual precipitation is 430 mm and occurs mainly during the summer season from June to September. Winter precipitation, which typically falls as snow, is low in this region15. The growing season normally starts in mid-May and lasts until mid-September. The vegetation is dominated by Kobresia pygmaea, accompanied by Potentilla saundersiana, Potentilla cuneata and Stipa purpurea.
Study design
The GE manipulation treatment was started in 2010 using a chain link fence in a flat area covering approximately 1 × 1 km2. The GE treatments were arranged using a randomized block design, each block covering an area of 60 × 60 m2. By 2015, there were a total of six blocks. The treatments considered in this study include grazing (G); fenced for 1 year (F1); fenced for 2 years (F2); fenced for 3 years (F3); fenced for 4 years (F4); and fenced for 5 years (F5). Within each block, six 2 × 2 m2 plots in a diagonal direction, while avoiding block edges, were randomly delineated. On an overall basis, this design included 36 plots under six treatment levels and six replicates for each treatment. While the experiment was conducted over a five-year period, all measurements were taken in 2015.
Data collection
In one of the reference blocks, soil temperature (°C) and volumetric water content (%) were continuously monitored at a 5 cm depth in 2015 using Decagon EC-TM sensors (Decagon Devices, Pullman, Washington, USA). Two soil sensors were installed for each treatment and the average of the two readings was recorded. Litter from six 30 × 30 cm2 subplots within each block was collected twice, during the early and late growing seasons. Collected litter was oven-dried at 65 °C to a constant weight. Litter layer depth (cm) was measured using a ruler. Soil samples were collected with a soil auger at three depths (0–10, 10–20 and 20–30 cm). Soil sampling was repeated three times for each block. 54 soil samples were rinsed from roots under running water over a 2-mm screen and dried at 105 °C for 12 h; their C and N concentrations were then measured using an Elementar Vario EL C/Nanalyzer (Elementar, Hanau, Germany). The soil P content was determined using the H2SO4-HClO4 fusion method.
K. pygmaea, P. saundersiana, P. cuneata, S. purpurea and Festuca coelestis were selected as the focal species, whose coverage and biomass account for more than 90% of the community. In May, 2015, ten individuals of K. pygmaea and five individuals of other species were selected from each plot and then marked using a color-coded tag. The phenological processes of each selected individual were scored every 3–5 days using a scoring method modified from Dunne et al. (2003) and Xia et al.28,29. For forbs, the following codes were recorded: sprout-out leaf: 0; unopened buds: 1; opened flowers: 2; old flowers: 3; initiated fruits: 4; enlarged fruits: 5; dehisced fruits: 6; withered plants: 7. The following codes were applied to grasses: sprout-out leaf: 0; plant booting stage: 0.5; presence of spikelets: 1; exerted anthers or styles: 2; past the presence of anther and styles: 4; disarticulated florets: 6; and withered plants: 7. On each census day, unweighted averages of phenological scores for each individual plant were calculated30. For example, a plant with one bud (‘1’), three old flowers (‘3’) and four expanding fruits (‘5’) received a phenological score of 3.031.
Based on their morphological and life-history traits, K. pygmaea were classified as either low-growing (3.2 cm height), shallow-rooted (10 cm depth) or early-flowering species (DOY: 155 ± 0.8). In a similar vein, P. saundersiana and P. cuneata were classified as low-growing (4.0 cm height), shallow-rooted (10 cm depth) or mid-flowering species (DOY: 168 ± 0.4; DOY: 172 ± 1.1). Finally, S. purpurea and F. coelestis were classified as tall-growing (25 cm height), deep-rooted (30 cm depth) or late-flowering species (DOY: 193 ± 0.5; DOY: 197 ± 1.2; DOY: 208 ± 0.4).
Data analysis
It is most difficult to directly obtain the exact timing of flowering and fruiting on the basis of observations taken at 3–5 day intervals29. Usually these data are extracted by fitting the observed data to statistical models, such as a linear regression model30, or the Richards growth equation. For example, the latter has been successfully applied to derive daily plant phenology in a tallgrass prairie in North America32 and a semi-arid temperate steppe in Inner Mongolia, China29. The relevant equation is:

where K is the maximum growth; a is a parameter related to the first observation date; b is the growth rate over time X in days; and m is a parameter related to the curve shape29. The timing of each phenological event can be calculated from Equation 1 as:

In this study, the Richards equation with the contraction–expansion algorithm was applied to fit phenological scores (Y) of each species against the day for each plot; this was accomplished utilizing Matlab (Mathworks, Natick, MA, USA). Best parameter estimates of K, a, b and m were obtained for each species. Equation (2) was applied to calculate the vegetative timing (i.e., green-up, ‘0’), reproductive timing (i.e., flowering and fruiting, ‘2’ and ‘3.5’) and the growing season length (GSL).
Statistical analysis
A two-way analysis of variance (ANOVA) with Turkey’s test was applied to disentangle the effects of the plant species (K. pygmaea, P. saundersiana, P. cuneata, S. purpurea and Festuca coelestis) and treatments for grazing (G) and grazing exclusion (F5, F4, F3, F2, F1) on green-up, flowering, timing of fruiting and growing season length. A one-way ANOVA with Tukey’s HSD test were used to analyze the effects of grazing and grazing exclusion treatments on litter depth, litter weight, soil conditions, phenological stages and reproductive success. All comparisons were considered significantly different at P < 0.05. All statistical tests were run using SPSS (v. 19.0; Chicago, USA).
Results
Microclimate, litter layer and soil nutrients
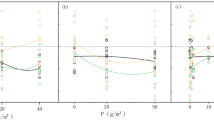
The consequences of each fencing treatment were compared to the grazing control treatment. The F5, F4, F3, F2 and F1 treatments increased growing season mean soil moisture at 5 cm depth by 6.0, 4.9, 3.5, 2.2, 1.7%, respectively (Fig. 1A). The F5, F4, F3, F2 and F1 decreased mean soil temperature at 5 cm depth by 1.5, 1.1, 0.8, 0.6 and 0.5 °C, respectively (Fig. 1B). The F2 and F1 treatments did not significantly change the amount of litter (Fig. 2A, P > 0.05). The F5, F4 and F3 treatments significantly increased the litter layer weight by 308%, 195% and 127%, respectively (Fig. 2A, P < 0.05). Compared to the grazing treatment, grazing exclusion (GE) significantly increased litter layer depth (P < 0.05), except for F1 (Fig. 2B, P > 0.05).There were significant differences in litter layer weight and depth between treatments F5 and F4, F3 (Fig. 2, P < 0.05).The five fencing treatments caused no significant effects on total soil content of C, N, P or the C/N ratio (Table 1, P > 0.05).

Mean soil moisture (%) at 5 cm depth [A], soil temperature at 5 cm depth (°C) [B] and daily precipitation (mm) [A] during the growing season under different fencing treatments in 2015. F5, F4, F3, F2, F1 and G represent plots with fencing for 5, 4, 3, 2 and 1 year (s) and grazing, respectively.

Mean weight (A) and depth (B) of the litter layer under different fencing treatments during the growing season of 2015. Different letters indicate significant differences at 5% level among treatments. F5, F4, F3, F2, F1 and G represent plots with fencing for 5, 4, 3, 2 and 1 year (s) and grazing, respectively.
Phenological stages
The results of two-way ANOVAS showed that the grazing exclusion (GE) treatments and plant species both exerted significant effects on the green-up, flowering and fruiting time, as well as length of growing season length (Table 2, P < 0.01), also significant interactive effects on the phenological events (Table 2, P < 0.01). For the phonological events, consequences of each fencing treatment were compared only with the control grazing treatment. The GE did not significantly affect plant phenology of Sp and Fc (Fig. 3A, P > 0.05). The F1 and F2 treatments did not significantly change plant phenology of any of the five selected species (Fig. 3, P > 0.05). The F3, F4 and F5 treatments significantly advanced the green-up dates for Kp, Ps and Pc (Fig. 3A, P < 0.05). The F3 and F4 treatments significantly advanced the flowering dates for Kp, Ps and Pc (Fig. 3B, P < 0.05). In contrast, the F5 treatment significantly delayed the flowering dates for Kp, Ps and Pc by 9.5 (±1.3 SE), 6.7 (±1.0) and 6.0 (±0.7) days, respectively (Fig. 3B, P < 0.05). The GE treatments did not significantly change the fruiting date for any of the five species (Fig. 3C, P > 0.05), with the notable exception of F5, which significantly delayed the fruiting date in Kp, Ps and Pc by 4.5 (±0.7), 3.7 (±0.5) and 3.3 (±0.7) days, respectively (Fig. 3C, P < 0.05). With respect to growing season length (GSL), the F1 and F2 treatments did not significantly change the GSL for any of the five species (Fig. 4, P > 0.05). The F3, F4 and F5 significantly extended the GSL of Kp, Ps and Pc (Fig. 3A, P < 0.05). For example, the GSL of Kp was extended by 8.2 (±1.2), 9.5 (±1.5) and 10.6 (±1.3) days under treatments F3, F4 and F5, respectively (Fig. 4, P < 0.05).

Changes (in days) in the green-up (A), flowering (B) and fruiting (C) times for five fencing treatments compared with the grazing plot in 2015. A positive value indicates later green-up, flowering or fruiting than that in the grazing plot; while a negative value indicates earlier green-up, flowering or fruiting than in the grazing plot. Data are mean ± SE for advanced or delayed phenology. * indicates significant differences at the 5% level between grazing exclusion and grazing treatments. F5, F4, F3, F2, F1 and G represent plots with fencing for 5, 4, 3, 2, 1 years and grazing, respectively. Kp, Ps, Pc, Sp, Fc represent Kobresia pygmaea, Potentilla saundersiana, Potentilla cuneata, Stipa purpurea and Festuca coelestis, respectively.

Changes in the growing season length (GSL) in five fencing treatments compared to grazing plot in 2015.
Negative values (−) indicate shortened a GSL and positive values (+) indicate an extended GSL. Data are mean ± SE for shortened or extended days. * indicate significant differences at 5% level between grazing exclusion and grazing treatments. F5, F4, F3, F2, F1 and G represent plots with fencing for 5, 4, 3, 2, 1 years and grazing, respectively. Kp, Ps, Pc, Sp, Fc represent Kobresia pygmaea, Potentilla saundersiana, Potentilla cuneata, Stipa purpurea and Festuca coelestis, respectively.
Reproductive success
Compared to the grazing treatment, the F1, F2 and F5 treatments did not significantly affect the maximum number of flowers in K. pygmaea and P. saundersiana (Fig. 5A,D, P > 0.05), while the F3 and F4 significantly increased the number of flowers of these two species (Fig. 5A,D, P < 0.05).The maximum number of fruits in K. pygmaea and P. saundersiana was higher under the F3 and F4 treatments than under the grazing treatment (Fig. 5B,E, P < 0.05). However, the F5 treatment significantly reduced the maximum number of fruits in K. pygmaea and P. saundersiana compared with treatments F3 and F4 (Fig. 5B,E, P < 0.05). Exclusion of grazing (GE) significantly increased reproductive success for K. pygmaea (Fig. 5C, P < 0.05). The reproductive success of P. saundersiana was not affected by any treatment when compared to the grazing treatment (Fig. 5F, P > 0.05).

Effects of six experimental treatments on the maximum number of flowers, fruits and reproductive success of Kobresia pygmaea (A–C), Potentilla saundersiana (D–F) in 2015. Data are mean ± SE. Different letters indicate significant differences at the 5% level among treatments. F5, F4, F3, F2, F1 and G represent plots with fencing for 5, 4, 3, 2, 1 years and grazing, respectively.
Discussion
Microclimate, litter layer and soil nutrients
Decomposition of plant litter is a key process in nutrient and carbon cycling for terrestrial ecosystems33. The litter decomposition in cold biomes, such as alpine meadows, is strongly limited by temperature34,35. As shown in the present study, the weight and depth of the litter layer increased with the length of the grazing exclusion period. Thus by thickening the litter layer, grazing exclusion lowered water loss from soil evaporation23 and ameliorated upper-soil moisture conditions. Soil temperature in the grazing exclusion plots was lower than that in the grazing plots, which is in line with previous studies on the Tibetan plateau35. Grazing exclusion has been considered to be an effective management practice capable of boosting soil C and N contents in rangelands36. This study found that grazing exclusion failed to alter total C, N, P content in the soil as well as the C/N ratio8, which might be a consequence of the short period of grazing exclusion treatment in this work. In the northern Tibetan grasslands, temperature is extremely low and litter decomposes slowly. As a result, a large proportion of the enriched litter in grazing-excluded sites have not as yet entered the soil after several years of grazing exclusion.
The timing of green-up
The grazing exclusion treatments, such as F3, F4 and F5, advanced the green-up date for low-growing, shallow-rooted species. In the alpine meadows of Northern Tibet, these species normally turned green in late May, a dry and pre-monsoon period. The ameliorated soil moisture condition23, especially in the upper soil, under grazing exclusion treatment may stimulate growth of low-growing, shallow-rooted plants such as Kp, Ps and Pc. This is in accord with the previous findings that the shallow-rooted plants in the northern Tibetan plateau are more sensitive to upper-soil moisture15,37. In addition, spring phenology responses to climate change appear not to be strongly constrained by photoperiod38. Grazing exclusion, via a thickened litter layer, may influence photoperiod16,22, but in this study it had no effect on the timing of green-up. Tall-growing, deep-rooted species, such as Sp and Fc, are able to utilize deep soil water. Their phenological development is less constrained by water availability compared to that of low-growing, shallow-rooted species15. The increased soil moisture under grazing exclusion may not reach the level to stimulate the phenological processes of the deep-rooted species.
The timing of flower and fruit
Grazing exclusion treatments, such as F3 and F4, advanced the flowering date for the low-growing, shallow-rooted species. Through thickening of the litter layer, grazing exclusion increased soil moisture, resulting in accelerated plant growth and consequently advancing the flowering date39. Soil temperature in the grazing exclusion plots was lower than that of the grazing plots during the growing season, yet phenology was still advanced under treatments F3 and F4. Thus soil moisture, rather than soil temperature, was likely the major phenological cue for the alpine grassland in Northern Tibet15,40. The process of producing fruit follows several other predecessor processes. Although the green-up and flowering times of low-growing, shallow-rooted species were advanced under the F3 and F4 treatments, plants may compensate for changes in individual phenological events. As a result, the fruit timing was unaltered41.
Plant phenology is affected by a variety of climatic factors, including humidity15 (precipitation and soil moisture), photoperiod16, temperature14 and winter chilling42. Interestingly, the flowering and fruiting responses of the low-growing, shallow-rooted species to the grazing exclusion period were reversed for the fifth year of grazing exclusion. Photoperiod responses to plant phenology are driven by the circadian clock43 and these responses may serve as a buffer to avoid, for example, the immediate response of phenology to temperature. This acclimation might be a potential reason for the reversed responses of plant phenology for the fifth year of grazing exclusion. Another reason why the pattern was reversed under the F5 treatment may be decreased radiation due to a thickened litter layer44. The low-growing, shallow-rooted species are buried under the litter layer, especially by the fifth year of grazing exclusion. The tall-growing, deep-rooted species can utilize their height advantage to spike through the litter layer. Thus their reproductive development is not constrained by photoperiod and radiation amount. The differentiated phenology responses to the grazing exclusion period gradient illustrated that the photoperiod and radiation level are the main factors regulating plant reproductive growth under grazing exclusion treatments.
The growing season length and reproductive success
Climate warming has been reported to extend the length of the growing season32,45, principally through an earlier beginning or later termination thereof46. For the alpine meadow ecosystem in Northern Tibet, grazing exclusion extended growing season lengths (GSLs) of low-growing, shallow-rooted species by advancing their green-up timing, which is consistent with findings for Northern Tibet based on remote sensing47.
The grazing exclusion treatments, such as F3 and F4, increased the number of flowers and fruits in K. pygmaea and P. saundersiana. Dorji et al. (2013) have reported that warming, via decreasing soil moisture, can suppress the reproductive effort of K. pygmaea15. Conversely, grazing exclusion, via increased soil moisture, particularly in the upper soil, can stimulate the reproductive efforts of low-growing, shallow-rooted species in Northern Tibet. The F3 and F4 treatments also significantly increased reproductive success in K. pygmaea. Such improved success resulted mainly from the combined effects of changes in soil moisture and presence of grazing animals31. However, grazing exclusion failed to alter the reproductive success rate of P. saundersiana. Previous studies have reported that feeding selection by livestock can influence species composition and community structure48. In Northern Tibet, grazing livestock tend to feed on species of Cyperaceae (K. pygmaea) and Gramineae (Stipa purpurea). Grazing exclusion, through removal of livestock, would be more likely to increase reproductive success of K. pygmaea, while having less (or in the case of this study, no) effects on forbs such as P. saundersiana.
Despite increased soil moisture, the F5 treatment significantly reduced the maximum number of flowers, fruits and reproductive success for K. pygmaea and P. saundersiana compared to the F3 and F4 treatments. Photoperiodic constraints on plant phenology can also affect photosynthetic activity of plants49. In northern Tibet, the F5 treatment, via an increased litter layer, can influence the photoperiod16,22 and photosynthetic activity of the low-growing, shallow-rooted species, further suppressing their reproductive development and consequently reducing the number of flowers, fruits and reproductive success.
Grazing exclusion strongly affected alpine plant phenology and reproductive success in the focal alpine system of this study. Species traits, such as low- vs. tall growing and shallow vs. deep rooting depth, can mediate how alpine plant species respond to grazing exclusion. The low-growing, shallow-rooted species, such as K. pygmaea, are more sensitive to grazing exclusion due to changes in upper-soil moisture and light resources. Grazing exclusion generally advanced the phenology of K. pygmaea, yet exclusion for five years delayed the reproductive timing and success of K. pygmaea via thickening of the litter layer and effect on the photoperiod. The tall-growing, deep-rooted species can spike through the litter layer due to their height advantage. Thus their reproductive development is not constrained by light limitations.
Related studies have reported that grazing exclusion can decrease species diversity and soil organic C sequestration in our targeted alpine system26,50. This study provided evidence that three and four years of grazing exclusion can advanced the phenology of low-growing, shallow-rooted species, while exclusion of grazing for five years can postpone their reproductive timing. Thus, three to four years of grazing exclusion may be an efficient way to restore degraded grasslands in the Tibetan alpine meadow. As we did not directly investigate light quantity and photoperiod under GE treatments, results of this study were not able to distill out the effects of photoperiod on phenological development. Thus, future work is required to examine the sole effect of photoperiod on plant phenology under grazing exclusion measures.
Additional Information
How to cite this article: Zhu, J. et al. Effects of short-term grazing exclusion on plant phenology and reproductive succession in a Tibetan alpine meadow. Sci. Rep. 6, 27781; doi: 10.1038/srep27781 (2016).
References
Wright, I. J. et al. The worldwide leaf economics spectrum. Nature 428, 821–827 (2004).
Wang, W. & Fang, J. Y. Soil respiration and human effects on global grasslands. Global Planet. Change 67, 20–28 (2009).
Schönbach, P. et al. Grassland responses to grazing: effects of grazing and management system in an Inner Mongolian steppe ecosystem. Plant Soil 340, 103–115 (2011).
Li, W. J. et al. Plant communities, soil carbon and soil nitrogen properties in a successional gradient of sub-alpine meadows on the eastern Tibetan Plateau of China. Environ. Manage. 44, 755–765 (2009).
Zhou, Z. Y. et al. Dynamics of vegetation and soil carbon and nitrogen accumulation over 26 years under controlled grazing in a desert shrubland. Plant Soil 341, 257–268 (2011).
Golodets, C., Kigel, J. & Sternberg, M. Recovery of plant species composition and ecosystem function after cessation of grazing in a Mediterranean grassland. Plant Soil 329, 365–378 (2010).
Korkanc, S. Y. Effects of afforestation on soil organic carbon and other soil properties. Catena 123, 62–69 (2014).
Medina-Roldán, E., Paz-Ferreiro, J. & Bardgett, R. D. Grazing exclusion affects soil and plant communities, but has no impact on soil carbon storage in an upland grassland. Agr. Ecosyst. Environ. 149, 118–123 (2012).
Mekuria, W. & Aynekulu, E. Exclosure land management for restoration of the soils in degraded communal grazing lands in northern Ethiopia. Land Degrad. Dev. 24, 528–538 (2013).
Han, J. J. et al. The effects of grazing and watering on ecosystem CO2 fluxes vary by community phenology. Environ. Res. 144, 64–71 (2015).
Penuelas, J. & Filella, I. Phenology-responses to a warming world. Science 294, 793–795 (2001).
Obrist, D. et al. Quantifying the effects of phenology on ecosystem evapotranspiration in planted grassland mesocosms using EcoCELL technology. Agric. For.Meteorol. 118, 173–183 (2003).
Xia, J. et al. Joint control of terrestrial gross primary productivity by plant phenology and physiology. Proc. Nat. Acad. Sci. USA 112, 2788–2793 (2015).
Wang, S. P. et al. Asymmetric sensitivity of first flowering date to warming and cooling in alpine plants. Ecology 95, 3387–3398 (2014).
Dorji, T. et al. Plant functional traits mediate reproductive phenology and success in response to experimental warming and snow addition in Tibet. Global Chang Biol. 19, 459–472 (2013).
Way, D. A. & Montgomery, R. A. Photoperiod constraints on tree phenology, performance and migration in a warming world. Plant Cell Environ. 38, 1725–1736 (2015).
Xi, Y. et al. Nitrogen addition alters the phenology of a dominant alpine plant in Northern Tibet. Arct. Antarct. Alp. Res. 47, 511–518 (2015).
Hülber, K., Bardy, K. & Dullinger, S. Effects of snowmelt timing and competition on the performance of alpine snowbed plants. Perspect. Plant Ecol. 13, 15–26 (2011).
Miller, J. J. et al. Influence of streambank fencing on the environmental quality of cattle-excluded pastures. J. Environ. Qual. 39, 991–1000 (2010).
Reeder, J. D. et al. Response of organic and inorganic carbon and nitrogen to long-term grazing of the shortgrass steppe. Environ. Manage. 33, 485–495 (2004).
Shao, C. et al. Spatial variation of net radiation and its contribution to energy balance closure in grassland ecosystems. Ecol. Process. 3, 1–11 (2014).
Kai, J. & Gutekunst, K. Effects of litter on establishment of grassland plant species: the role of seed size and successional status. Basic Appl. Ecol. 4, 579–587 (2003).
Miao, H. et al. Cultivation and grazing altered evapotranspiration and dynamics in Mongolia steppes. Agr. Forest Meteorol. 149, 1810–1819 (2009).
Fan, J. W. et al. Carbon storage in the grasslands of China based on field measurements of above- and below-ground biomass. Climatic Change 86, 375–396 (2008).
Zhong, X. H. et al. Research of Ecological Security on the Tibetan Plateau. J. Mt. Sci. 28, 1–10 (2010). (In Chinese with English abstract).
Shi, X. M. et al. Grazing exclusion decreases soil organic C storage at an alpine grassland of the Qinghai–Tibetan Plateau. Ecol. Eng. 57, 183–187 (2013).
Zeng, C. X., Wu, J. S. & Zhang, X. Z. Effects of grazing on above- vs. below-ground biomass allocation of alpine grasslands on the Northern Tibetan Plateau. PLoS ONE 10, e0135173 (2015).
Dunne, J. A., Harte, J. & Taylor, K. J. Subalpine meadow flowering phenology responses to climate change: Integrating experimental and gradient methods. Ecol. Monogr. 73, 69–86 (2003).
Xia, J. Y. & Wan, S. Q. Independent effects of warming and nitrogen addition on plant phenology in the Inner Mongolian steppe. Ann. Bot. 111, 1207–1217 (2013).
Price, M. V. & Waser, N. M. Effects of experimental warming on plant reproductive phenology in a subalpine meadow. Ecology 79, 1261–1271 (1998).
Han, J. J. et al. Grazing and watering alter plant phenological processes in a desert steppe community. Plant Ecol 216, 599–613 (2015b).
Sherry, R. A. et al. Divergence of reproductive phenology under climate warming. Proc. Nat. Acad. Sci. USA 104, 198–202 (2007).
Liski, J. et al. Climatic effects on litter decomposition from arctic tundra to tropical rainforest. Global Change Biol. 9, 575–584 (2003).
Aerts, R. The freezer defrosting: global warming and litter decomposition rates in cold biomes. J. Ecol. 94, 713–724 (2006).
Luo, C. Y. et al. Effect of warming and grazing on litter mass loss and temperature sensitivity of litter and dung mass loss on the Tibetan plateau. Global Change Biol. 16, 1606–1617 (2010).
Wu, G. L. et al. Long-term fencing improved soil properties and soil organic carbon storage in an alpine swamp meadow. Plant Soil 332, 331–337 (2010).
Schenk, H. J. & Jackson, R. B. Rooting depths, lateral root spreads and belowground/above-ground allometries of plants in water-limited ecosystems. J. Ecol. 90, 480–494 (2002).
Laube, J. et al. Chilling outweighs photoperiod in preventing precocious spring development. Global Change Biol. 20, 170–182 (2014).
Grime, J. P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 111, 1169–1194 (1977).
Körner, C. Alpine plant life: Functional plant ecology of high mountain ecosystems. Germany, Springer, Berlin. (2003).
Post, E. S. et al. Phenological sequences reveal aggregate life history response to climatic warming. Ecology 89, 363–370 (2008).
Robbirt, K. M. et al. Validation of biological collections as a sourceof phenological data for use in climate change studies: a casestudy with the orchid Ophryssphegodes. J. Ecol. 99, 235–241 (2010).
Víctor, R., James, H. & Anthony, H. Ecological implications of plants’ ability to tell the time. Ecol. Lett. 12, 583–592 (2009).
Facelli, J. M. & Pickett, S. T. A. Plant litter: light interception and effects on old-field plant community. Ecology 73, 1024–1031 (1991).
Parmesan, C. & Yohe, G. A. Globally coherent fingerprint of climate change impacts across natural systems. Nature 421, 37–42 (2003).
Piao, S. L. et al. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Global Biogeochem.Cy. 21, GB3018 (2007).
Song, C. Q. et al. Spatio-temporal variation of vegetation phenology in the Northern Tibetan Plateau as detected by MODIS remote sensing. Chin. J. Plant Ecol. 35, 853–863 (2011). (In Chinese with English abstract).
Noy-Meir, I., Gutman, M. & Kaplan, Y. Responses of Mediterranean grassland plants to grazing and protection. J. Ecol. 77, 290–310 (1989).
Bauerle, W. L. et al. Photoperiodic regulation of the seasonal pattern of photosynthetic capacity and the implications for carbon cycling. Proc. Nat. Acad. Sci. USA 109, 8612–8617 (2012).
Mao, S. J. et al. Response of the mountain performance in alpine grassland to enclosure on the Northern Tibetan Plateau. ActaPratacul. Sin. 24, 21–30 (2015). (In Chinese with English abstract).
Acknowledgements
This research was supported by the 973 Program of the Ministry of Science and Technology of China (2013CB956302, 2010CB950603); the National Natural Science Foundation of China (41571195) and the West Light Foundation of the Chinese Academy of Sciences.
Author information
Authors and Affiliations
Contributions
J.Z. carried out this research, collected field data, drafted the manuscript, performed data analysis and participated in the design of the study; Y.Z. conceived and designed the study, coordinated research and helped draft the manuscript; Y.L. collected field data. All authors gave final approval for publication.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zhu, J., Zhang, Y. & Liu, Y. Effects of short-term grazing exclusion on plant phenology and reproductive succession in a Tibetan alpine meadow. Sci Rep 6, 27781 (2016). https://doi.org/10.1038/srep27781
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep27781
This article is cited by
-
Effects of long-term grazing exclusion on vegetation structure, soil water holding capacity, carbon and nitrogen sequestration capacity in an alpine meadow on the Tibetan Plateau
Journal of Mountain Science (2023)
-
Exploring the most important indicators for environmental condition assessment using structural equation modeling and InVEST habitat quality model
Environmental Monitoring and Assessment (2023)
-
Plant phenology changes and drivers on the Qinghai–Tibetan Plateau
Nature Reviews Earth & Environment (2022)
-
CO2, CH4 and N2O fluxes in an alpine meadow on the Tibetan Plateau as affected by N-addition and grazing exclusion
Nutrient Cycling in Agroecosystems (2020)
-
Impact of long-term grazing exclusion on soil microbial community composition and nutrient availability
Biology and Fertility of Soils (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
