
Abstract
Lysophosphatidic acid (LPA), a growth factor-like phospholipid, regulates numerous physiological functions, including cell proliferation and differentiation. In a previous study, we have demonstrated that LPA activates erythropoiesis by activating the LPA 3 receptor subtype (LPA3) under erythropoietin (EPO) induction. In the present study, we applied a pharmacological approach to further elucidate the functions of LPA receptors during red blood cell (RBC) differentiation. In K562 human erythroleukemia cells, knockdown of LPA2 enhanced erythropoiesis, whereas knockdown of LPA3 inhibited RBC differentiation. In CD34+ human hematopoietic stem cells (hHSC) and K526 cells, the LPA3 agonist 1-oleoyl-2-methyl-sn-glycero-3-phosphothionate (2S-OMPT) promoted erythropoiesis, whereas the LPA2 agonist dodecyl monophosphate (DMP) and the nonlipid specific agonist GRI977143 (GRI) suppressed this process. In zebrafish embryos, hemoglobin expression was significantly increased by 2S-OMPT treatment but was inhibited by GRI. Furthermore, GRI treatment decreased, whereas 2S-OMPT treatment increased RBC counts and amount of hemoglobin level in adult BALB/c mice. These results indicate that LPA2 and LPA3 play opposing roles during RBC differentiation. The pharmacological activation of LPA receptor subtypes represent a novel strategies for augmenting or inhibiting erythropoiesis.
Similar content being viewed by others

Introduction
Lysophosphatidic acid (LPA) is generated by the lysophospholipase D enzyme autotaxin (ATX) and present in biological fluids at concentrations that can reach micromolar levels in serum. At least six G-protein-coupled LPA receptors (LPARs) have been described. The LPA1, LPA2 and LPA3 GPCR are belong to the endothelial differentiation gene family1, whereas LPA4, LPA5 and LPA6 belong to in the purinergic receptor cluster3. It has been shown that LPARs are involved in stem cell differentiation4. Previous results have indicated that LPA induces embryonic stem cell proliferation and differentiation by activating the phospholipase C (PLC)/Ca2+ signaling axis6. LPARs have also been identified in murine and human pluripotent stem cells, including in hematopoietic and embryonic stem cells7. Furthermore, it has been reported that induced pluripotent stem cells (iPSC) express LPA1−4 GPCR and that LPA induces iPSC differentiation and proliferation by activating the Rho/ROCK pathway8. However, the role of LPA and its GPCR during hematopoiesis remain elusive.
Hematopoiesis occurs in two waves during vertebrate development, a short-lived “primitive wave” that is characterized by embryonic globin expression; and a later “definitive wave” that develops intraembryonically in the aorto-gonadal-mesonephros region of the embryo9,10,11. The primitive wave of hematopoiesis is a transient process during embryonic development. This process produces unipotent blood cells that act as oxygen transporters and it is therefore essential for the viability of the embryo12. The definitive wave of hematopoiesis occurs in the fetal liver to produce hematopoietic stem cells (HSCs) that support the differentiation of all blood cell lineages13,14. Recent studies suggest that LPA plays an important role in regulating primitive hematopoiesis through activation of LPA115. It has also been shown that LPA promotes myeloid differentiation in the human bone marrow microenvironment16. LPA was recently shown to promote the differentiation of the myeloid/macrophage lineage from human CD34+ hematopoietic progenitors via activation of LPA2 in vitro4. Thus, corollary evidence suggests that LPA might regulate HSC differentiation and blood cell homeostasis.
We have demonstrated previously the role of LPA3 in red blood cell (RBC) differentiation17. In the present study, we report the pharmacological dissection of specific roles of LPA GPCR subtypes in three different species. Our results demonstrate that LPA2 and LPA3 exert opposing roles on RBC differentiation in vitro and in vivo. We also highlighted that 2S-OMPT, a LPA3 agonist, is a potential drug candidate that enhances erythropoiesis in vivo.
Results
LPA2 and LPA3 expression is differentially regulated during hemin-induced erythropoiesis in K562 cells
Using human hematopoietic stem cells we have demonstrated previously that LPA enhances erythropoiesis by activating LPA317. To examine the expression patterns of LPAR during differentiation, we applied hemin-induced erythropoiesis model system using the K562 cell line18. We first evaluated mRNA expression of lpar1, lpar2 and lpar3. Under unstimulated conditions, K562 cells abundantly expressed lpar2, a moderate level lpar3; however, lpar1 was barely detectable (Supplemental Fig. 1a). Upon hemin induction, erythropoiesis took place indicated by upregulation of γ-globin mRNA19 (Supplemental Fig. 1b). During differentiation, mRNA expression levels of lpar2 decreased significantly (p < 0.05) after 48 h of culture (Supplemental Fig. 1c). In contrast, lpar3 mRNA expression level remained unchanged (Supplemental Fig. 1d).
Opposing regulation of erythropoiesis by LPA2 and LPA3
The decrease of lpar2 expression during differentiation suggests its role in the regulation of erythropoiesis. We hypothesized that downregulation of LPA2 might represent disinhibition signal and promotes the erythroid differentiation of K562 from the undifferentiated status. Using lentiviral transduction, we selected K562 cells that stably express LPAR2 shRNA (shLPAR2) with 80% knockdown efficiency compared to scramble controls (Control) (Supplemental data 1e). Flow cytometry showed that shLPAR2 expressing cells displayed higher expression of the erythroid markers CD71 and glycophorin A (GlyA) compared to controls (Fig. 1a–c and Supplemental Fig. 2a–c), indicating that knockdown of LPA2 promoted erythropoietic differentiation of K562 cells. Based on our previous observation in HSC demonstrating that knockdown of LPA3 suppressed erythropoiesis17, we transfected LPA3 siRNA (siLPAR3) into K562 cells and found downregulation of the erythropoietic markers CD71 and GlyA (Fig. 1d–f and Supplemental Fig. 2d–f). These results taken together indicate that knockdown of LPA2 and LPA3 exert opposing effects on erythropoiesis in the K562 model.
Pharmacological manipulation of LPAR of hemin-induced erythropoiesis in K562 cells
Because LPA2 and LPA3 appeared to play opposing roles in the regulation of erythroid differentiation, we explored the use of pharmacological manipulation specifically targeting LPA2 and LPA3 receptors to confirm our finding from the knockdown experiments. We stimulated K562 cells with the LPA2-selective agonist, DMP and found that it significantly decreased the expression of γ-globin mRNA (Fig. 2a). The erythroid differentiation marker CD71 was also downregulated by DMP treatment (Fig. 2b,c). However, the expression of GlyA was unaffected (Fig. 2d,e). These results are consistent with the hypothesis that the activation of LPA2 inhibits erythroid differentiation. We also found that erythropoiesis was enhanced by treating K562 cells with the LPA3-specific agonist, 2S-OMPT. We found that exposure to as low as 80-to-100 nM 2S-OMPT resulted in significant upregulation of γ-globin expression (Fig. 2f). Flow cytometry analysis showed that 2S-OMPT increased GlyA protein expressions in a dose-dependent manner (Fig. 2i,j). Furthermore, we found that CD71 was induced by combined treatment with 2S-OMPT and hemin (Fig. 2h) whereas, it was unaffected in the absence of hemin (Fig. 2g). Considering that LPA1 expression in K562, is almost undetectable, these finding might suggest that activation of LPA3 but not LPA1, enhances erythroid differentiation. The mRNA expression profiling of LPA2 and LPA3 was also evaluated following LPAR agonist treatment to exclude transcriptional alterations in LPAR expression due to ligand exposure. The LPA2 specific agonist GRI977143 (Supplemental Fig. 3a,b) and the LPA3 agonist 2S-OMPT (Supplemental Fig. 3e,f) caused no detectable alteration in the expression patterns of LPA2 and LPA3 mRNA. However, the LPA2 agonist DMP significantly inhibited LPA2 mRNA expression (Supplemental Fig. 3c,d). The mechanism of decreased LPA2 mRNA expression remains unclear. Nonetheless, these results confirm the opposing regulation of erythropoiesis by LPA2 and LPA3 in K562 cell line model.
Opposing effects of LPA2 and LPA3 in erythropoiesis of CD34+ hHSC
To further consolidate our hypothesis, we isolated CD34+ hHSC from umbilical cord blood20 and treated the cells with the LPA2 agonist GRI. In CD34+ hHSC, GRI inhibited CD71 mRNA expression during erythropoiesis elicited by erythropoietin and stem cell factor treatment. However, activation of LPA2 had no significant effect on GlyA mRNA expression (Fig. 3a). This might be due to the fact that CD71 is an early RBC marker as opposed to GlyA, which is a late RBC marker21. In contrast, the LPA3 agonist 2S-OMPT significantly enhanced the mRNA expression level of CD71 and GlyA (Fig. 3b). Furthermore, we also examined CD71 protein levels by flow cytometry and found that it paralleled changes in the mRNA expression pattern: GRI decreased, whereas 2S-OMPT increased CD71 protein expression, (Fig. 3c,d). These results are consistent with our findings in the K562 model, corroborating that LPA2 inhibits, whereas activation LPA3 promotes erythropoiesis in hHSC.
Pharmacological blockade of LPA signaling inhibits hematopoiesis in zebrafish
Our in vitro models have established that pharmacological activation of LPA3 enhances, whereas LPA2 suppresses erythroid differentiation. We evaluated these effects using the zebrafish model in vivo17. In zebrafish, the constitutive expression of LPA3 at all developmental stages suggests that it plays an important role during early embryogenesis22. To investigate whether LPA3 regulates erythropoiesis, we exposed zebrafish embryos to the LPA1/LPA3 mixed antagonist Ki16425. Expression of hemoglobin in zebrafish embryos were detected by o-dianisidine staining after 24 h of treatment with Ki16425. Ki16425 treatment resulted in moderate-to-severe phenotypes in 60% of the fish (Fig. 4a) suggesting that blockade of LPA1 and LPA3 reduced hemoglobin production in zebrafish embryos. However, no significant effect was observed in response to 2S-OMPT treatment (Fig. 4b). To further evaluate the importance of LPA3 signaling during zebrafish erythropoiesis, we blocked zLPAR3 by injecting 8 ng of morpholino (MO) zLPAR3, which targets the non-overlapping site near the translation initiation site. The WISH results showed no perturbation of the of primitive erythrocyte marker hbaa1 (α-A1 globin)23, myeloid marker hbbe1.1 (β-E1 globin)23 and ikzf1 (ikaros)24, which is a marker expressed both in primitive and definitive erythrocytes (Fig. 4c). These results indicate that the effect of LPA3 on erythropoiesis is not dominant at 48 hpf, which represents the primitive hematopoiesis stage.
Blockade of LPA3 perturbs erythroid differentiation during the definitive wave of hematopoiesis in zebrafish
The primitive wave mainly produces erythrocytes in the intermediate cell mass at 24 hpf, whereas the mature erythroids that differentiate in the definitive wave after 48 hpf11,25 are found in caudal hematopoietic tissue (CHT)26,27,28. Definitive erythrocytes are enriched in the posterior blood island from 3.5 days post-fertilization (dpf)29; hence, we selected 96 hpf as the sampling time point. Hemoglobin expression increased after injection with zLPAR3 mRNA or treatment with 2S-OMPT for 96 h. Exposure to 2S-OMPT also rescued the erythropoietic defect caused by zLPAR3 tMO injection (Fig. 5a,b). The expression of the definitive erythroid lineage markers hemoglobin genes, including hbae1 (α-E1 globin) and β-E1 globin, was increased (Fig. 5c). These results demonstrate that LPA3 is an important regulator of the definitive wave of erythropoietic differentiation in HSC.
Knockdown of LPA2 increases hemoglobin expression during definitive hematopoiesis in zebrafish
We also investigated the roles of LPA2 during erythropoiesis in zebrafish by injecting zLPAR2 tMO into the yolk of embryos at the one-cell stage. Non-injected and MO-injected embryos were collected at 4 dpf and subjected to o-dianisidine staining. Knockdown of zLPAR2 using MOs enhanced the hemoglobin levels in CHT (Fig. 6a,b). To confirm the results obtained with the MO-injected embryos, the embryos were treated with the LPA2 agonist GRI. These results indicated that the percentage of embryos with severe RBC defect increased with the concentration of GRI (Fig. 6c,d). Taken together, our findings indicate that LPA2 plays an inhibitory role in RBC development in zebrafish.
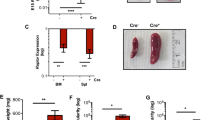
Activation of LPA receptors regulates erythropoiesis in mice
To further confirm the relationship between LPA2 and LPA3 GPCR in erythropoiesis of mammals, we injected 2S-OMPT and GRI intraperitonealy into 3-week-old BALB/c mice. The animals were sacrificed in post-injection day seven and blood samples were collected. The RBC count, HGB and HCT decreased significantly after GRI injection (Fig. 7a–c) and conversely increased under 2S-OMPT treatment (Fig. 7d–f). These results indicate that activation of LPA3 promotes whereas activation of LPA2 inhibits RBC differentiation in mice.

Effects of LPA2 and LPA3 on erythropoiesis in mice.
BALB/C mice were injected peritoneally with (a–c) 0.5 mg/kg 2S-OMPT and (d,e) 1 mg/kg GRI every day. The mice were sacrificed on post-injection day eight. Blood samples were isolated from the facial vein. (a,d) the RBC counts, (b,e) HGB and (c,f) HCT were determined. Data are normalized to the results obtained from day 0. The quantitative data represent the mean ± SD of at least four mice. *p < 0.05, *p < 0.01 and ***p < 0.005 indicate significant differences compared with vehicle control.
Discussion
Recent studies have reported the importance of LPA during mesenchymal stem cell differentiation30 and in the maintenance of multipotency of neuronal stem cells31. However, little is known about effect of LPA in hematopoiesis. Our previous study demonstrated that in the presence of EPO, LPA is an important regulator of erythropoiesis via LPA3 activation during EPO induction17. We also demonstrated the opposing roles of LPA2 and LPA3 during megakaryocytic differentiation in the K562 cell line32. Herein, using in vivo and in vitro models, we confirmed the role of LPA3 and the involvement of LPA2 in RBC differentiation. We also demonstrated for the first time that erythropoiesis can be modulated by treatment with specific agonists of LPARs.
2S-OMPT, a selective LPA3 agonist, has already been tested on renal function33, neurite growth34 and smooth muscle contraction35. We now have demonstrated that activation of LPA3 by 2S-OMPT also enhances hemoglobin mRNA transcripts, expression of erythroid markers and the number of circulating RBC. Interestingly, 2S-OMPT treatment appeared to be more effective on GlyA expression than on CD71 in K562 cells and hHSC. During early erythrocytic development, CD71 is upregulated in erythroblasts, whereas GlyA increases subsequently during the terminal differentiation stage21. The differential responses of 2S-OMPT on GlyA and CD71 expression suggests LPA3 exerts its effect on late erythropoiesis. Using the zebrafish model, hemoglobin levels were decreased within 24 h of Ki16425 treatments. However, activation of LPA3 by 2S-OMPT enhanced hemoglobin expression only during the late stage of embryonic development. Consistent with these observations, LPA1 has been reported as a developmental cue to regulate early erythropoiesis15. These results suggest that LPA1 and LPA3 regulate erythropoiesis during different stages of development. Based on our findings here we propose that LPA3 is primarily involved during definitive erythropoiesis and is responsible for terminal erythrocyte differentiation. Traditionally, administration of recombinant human EPO (rHuEPO) is the most common treatment for anemia. However, numerous side effects of EPO have been reported36. In cancer-related anemia, activation of EPO receptor might also promote cancer progression and metastasis37,38,39. Evidence exists that rHuEPO-induced erythropoiesis also has side effects on the sensitivity to radiotherapeutic or chemotherapeutic outcomes in cancer patients40. Our results demonstrated that 2S-OMPT-treated mice had more RBC, higher HGB expression and HCT ratio, suggesting a novel potential therapeutic strategy for the treatment of erythropoietic disorders. Appropriate administration of 2S-OMPT or similar specific LPA3 agonists might lower EPO dose necessary, thereby reducing the risk of undesirable side effects of long-term EPO therapy.
In the present study, we reported significant downregulation of LPA2 transcripts during hemin-induced erythropoiesis. In contrast to 2S-OMPT, activation of LPA2 by the DMP or GRI agonists suppressed erythroid marker gene expression. Knockdown of LPA2 promoted erythrocyte differentiation both in vitro and in vivo, suggesting that downregulation of LPA2 may provide a signal for erythroid-lineage commitment. Paradoxically, CD71 was more responsive to the activation of LPA2 in both the K562 and hHSC models. We propose that LPA2 exerts an inhibitory control at the early megakaryocyte-erythroid progenitor (MEP) stage by blocking terminal erythroid differentiation. Indeed, our previous data showed that activation of LPA2 inhibits the megakaryocytic transcriptional programs32. The high expression levels of LPA2 in unstimulated K562 also suggest its role in the maintenance of undifferentiated status. We propose that LPA2 signaling might be a molecular switch, which determines early HSC cell fate. It was reported that activation of LPA2 by GRI mitigates hematopoietic radiation syndrome4, suggesting activation of LPA2 acts as a radioprotector for HSC self-renewal. Given that GRI treatment significantly reduces circulatory RBC numbers, our results suggest the potential application of LPA2 agonists on several myeloproliferative diseases, including thrombocythemia, polycythemia vera and primary myelofibrosis41.
Within the hematopoietic niche, several microenvironmental factors are necessary for HSC development42. Furthermore, interaction of cell surface molecules between bone marrow stromal cell and HSC also play an important role during hematopoiesis43. Previous studies reported that LPA promotes the expression of many of these factors during hematopoiesis44,45 in endothelial cells in the hematopoietic niche. Recent reports also demonstrated the role of LPA during early hematopoiesis15. In addition, the major catalytic enzyme of LPA, autotoxin, is highly expressed in the vicinity of the myeloid progenitors16. Therefore, LPA might be presented as a constitutive signal during myeloid differentiation. Our results demonstrated that activation of LPA2 and LPA3 exert opposing effects on erythropoiesis. We propose that decision of MEP cell fate might depend on the temporal pattern expression of these two LPA GPCR subtypes. Nonetheless, the downstream mechanisms originating from LPA2 and LPA3 that regulate erythropoiesis/megakaryopoiesis require further investigation.
Overall, our results suggest that agents similar to GRI and 2S-OMPT could become powerful drug candidates to regulate erythropoiesis. Additionally, we have also demonstrated the inhibitory role of LPA2 during RBC differentiation. These observations combined may facilitate the development of new treatment strategies for erythropoietic disorders.
Methods
Cell culture
K562 cells were cultured in RPMI supplemented with 10% FBS as described previously32. Mononuclear cells were isolated from umbilical cord blood by Ficoll-Paque (Amersham Biosciences, Uppsala, Sweden) density gradient centrifugation after obtaining the donor’s consent according to governmental regulations (“Guidelines for collection and use of human specimens for research”, Ministry of Health and Welfare, Taiwan) approved by the Institutional Review Board of the Taoyuan General Hospital, Taiwan. The isolation and culture methods for hHSC were described previously17. Briefly, human CD34+ hHSC were isolated from mononuclear cells by magnetic micro-bead isolating methods using Direct Progenitor Isolation Beads (Miltenyi Biotech, Bergisch Gladbach, Germany) and MACS LS-columns (Miltenyi Biotech). 2.5 × 105 hHSC cells were cultured in 5 ml of EDM (5 × 104 cells/ml) in the presence of SCF (50 ng/ml) and EPO (6 IU/ml) for 6 days.
Pharmacological treatment
A 1 mM stock solution of LPA (18:1, Sigma-Aldrich), 2S-OMPT (Cayman Chemical), Ki16425 (Cayman), GRI97714346, dodecyl monophosphate (DMP; Sigma-Aldrich) and sphingosine-1-phosphate (Sigma-Aldrich) was prepared respectively as described in our previous report32. K562 cells were serum-starved for 12–16 h and cultured in 0.5% FBS medium containing the agonist and antagonist in the presence of hemin for 48 h. CD34+ hHSC were cultured with the test compounds diluted in the medium with 0.005% fatty acid-free bovine serum albumin.
RNA isolation and real-time polymerase chain reaction (RT-PCR)
Total RNA was isolated using the TRIzol® reagent (Thermo-Fisher). Thirty zebrafish embryos were ground up and total RNA was collected from the whole embryos. The level of each target mRNA was evaluated using quantitative RT-PCR (qPCR) based on the real-time threshold cycle and normalized against the amount of GAPDH. The specific primer sequences were as follows: lpar1: forward (5′->3′): TTCAACTCTGCCATGAA CCCC, reverse (3′->5′): CTAAACCAC AGAGTGGTCATT; lpar2: forward (5′->3′): ACACTT CTGGCACTGC CTCT, reverse (3′->5′): AGGCTGAGTGTGGTCTCTCG; lpar3: forward (5′->3′): TCAGC AGGAGTGACACAGGCAG, reverse (3′->5′): GGAAGTGCTTTTATTGCAGACT; γ-globin: forward (5′->3′): GCAGCTTGTCA CAGTGCAGTTC, reverse (3′-> 5′): TGGCAAGAAGGTGCTGACTTC; CD71: forward (5′>3′): GGATAAAGCGGTTCTTGGTACC, (3′-> 5′): CCAGTAACCGGA TGCTTCACA; GlyA: forward (5′->3′): ACAGACAAATGATACGCACAAACGGG, reverse (3′->5′): GGGCTTTTCTTTAT CAGTCGGCGA; GAPDH: forward (5′->3′): AAGGTGAAGGTCGGAGTC, reverse (3′->5′): TGTAGTTGAGGTCAATGAAGG.
Flow cytometry
K562 cells and CD34+ hHSC were collected in phosphate-buffered saline (PBS) and stained with anti-CD71-FITC antibodies and anti-GlyA-PE antibodies (BD Bioscience Pharmingen; San Diego, CA, USA) for 30 min. Mouse bone marrow cells were collected in PBS and stained with anti-CD71-FITC antibodies (BD Bioscience Pharmingen) for 30 min. The stained cells were then washed three times and resuspended in PBS. All the cell samples were analyzed using a Cyto-flow instrument (Partec; Muenster, Germany) and the FCS Express software (De Novo; Los Angeles, CA, USA).
RNAi transfection
Production of lentiviral stocks was described in detail in our previous study47. The LPA2 shRNA template was inserted into the pLKO.1 lentiviral vector = purchased from the National RNAi Core Facility Platform, Academia Sinica. The shRNA target sequence was: 5′-CCTGGTCAAGACTGTTGTCAT-3′ (shLPAR2; TRCN0000221131). The transfection and selection methods for transient or stable knockdown cells were described in our previous study32.
Maintenance of zebrafish
The zebrafish were maintained according to an approved National Taiwan University Zebrafish Core Laboratory standard operating protocol for animal use. The zebrafish handling, breeding and staging methods were performed as described previously17,48.
Morpholino oligonucleotides
The MO stocks were prepared in sterile double-distilled water and 4-ng MO was injected with 5% phenol red. MO sequences were as follows: MO-zLpar2: 5′-CCAGCCCTAAAACACAGGAAGACAT-3′, MO-zLpar3: 5′-CAGCCCTAAAACAC AGGAAGACAT-3′. The MO stocks were prepared in sterile double-distilled water at a final concentration of 24 μg/ml. Each MO was injected at a 4-ng dose with 5% phenol red.
O-dianisidine staining
O-dianisidine staining was used to study the expression of globin49. The embryos were collected at various stages of development and stained for 15 min in the dark with o-dianisidine (0.6 mg/ml), 0.01 M sodium acetate (pH 4.5), 0.65% H202 and 40% (V/V) ethanol. The stained embryos were cleared with benzyl benzoate/benzyl alcohol (2:1, V/V) and examined by differential interference contrast microscopy50.
Whole-mount RNA in situ hybridization (WISH)
The embryos were digested with 10 μg/mL proteinase K and fixed with 4% paraformaldehyde. Next, the embryos were incubated with 0.015U of digoxigenin-labeled anti-sense RNA probes for 14–16 h at 60 °C. Anti-sense mRNA probes for α-A1, β-E1, ikaros, and hbae1 were labeled with digoxigenin (Roche, Basel, Switzerland). The embryos were treated with RNase and washed to a stringency of 0.2× saline-sodium citrate buffer/hybridization buffer (SSC/HYB) at 60 °C. Finally, the embryos were exposed to Nitro tetrazolium blue/5-bromo-4-chloro-3-indoyl phosphate (Roche) in detection solution and incubated in the dark for 15–30 min51.
Treatment of mice with LPA GPCR pharmacons
14-day-old male BALB/c mice (LASCO, Taipei, Taiwan) were dosed with a 0.5 mg/kg 2S-OMPT or 1 mg/kg GRI. Both chemicals were diluted in PBS with 3% BSA to a final volume of 300 μL and were injected peritoneally. Facial vein blood samples were taken on the seventh day and mixed with an equal volume of 2 mg/mL EDTA. The number of RBC, hemoglobin and hematocrit was analyzed using a Sysmex XT-2000i Automated Hematology Analyzer (Sysmex, Taiwan). All animal procedures were authorized and carried out in accordance with the approved guidelines and regulations of the Institutional Animal Care and Use Committee of the National Taiwan University.
Statistical analysis
All the experiments were repeated at least three times. Significant differences were calculated using one-way analysis of variance. The statistical analyses were performed using StatView software (Abacus Concept). Results are expressed as the mean ± standard deviation based on at least three independent experiments. p < 0.05 was considered statistically significant in all the tests.
Additional Information
How to cite this article: Lin, K.-H. et al. Pharmacological activation of lysophosphatidic acid receptors regulates erythropoiesis. Sci. Rep. 6, 27050; doi: 10.1038/srep27050 (2016).
References
Hecht, J. H., Weiner, J. A., Post, S. R. & Chun, J. Ventricular zone gene-1 (vzg-1) encodes a lysophosphatidic acid receptor expressed in neurogenic regions of the developing cerebral cortex. J Cell Biol 135, 1071–1083 (1996).
Contos, J. J., Ishii, I. & Chun, J. Lysophosphatidic acid receptors. Molecular pharmacology 58, 1188–1196 (2000).
Janssens, R., Boeynaems, J. M., Godart, M. & Communi, D. Cloning of a human heptahelical receptor closely related to the P2Y5 receptor. Biochemical and biophysical research communications 236, 106–112, 10.1006/bbrc.1997.6895 (1997).
Patil, R. et al. Combined Mitigation of the Gastrointestinal and Hematopoietic Acute Radiation Syndromes by an LPA2 Receptor-Specific Nonlipid Agonist. Chemistry & biology 22, 206–216, 10.1016/j.chembiol.2014.12.009 (2015).
Deng, W. et al. Mitigation of the Hematopoietic and Gastrointestinal Acute Radiation Syndrome by Octadecenyl Thiophosphate, a Small Molecule Mimic of Lysophosphatidic Acid. Radiation research, 10.1667/rr13830.1 (2015).
Ermakov, A. et al. A role for intracellular calcium downstream of G-protein signaling in undifferentiated human embryonic stem cell culture. Stem cell research 9, 171–184, 10.1016/j.scr.2012.06.007 (2012).
Pebay, A., Bonder, C. S. & Pitson, S. M. Stem cell regulation by lysophospholipids. Prostaglandins & other lipid mediators 84, 83–97, 10.1016/j.prostaglandins.2007.08.004 (2007).
Frisca, F., Crombie, D. E., Dottori, M., Goldshmit, Y. & Pebay, A. Rho/ROCK pathway is essential to the expansion, differentiation and morphological rearrangements of human neural stem/progenitor cells induced by lysophosphatidic acid. Journal of lipid research 54, 1192–1206, 10.1194/jlr.M032284 (2013).
Dzierzak, E. The emergence of definitive hematopoietic stem cells in the mammal. Current opinion in hematology 12, 197–202 (2005).
Cumano, A. & Godin, I. Ontogeny of the hematopoietic system. Annual review of immunology 25, 745–785, 10.1146/annurev.immunol.25.022106.141538 (2007).
Weinstein, B. M. et al. Hematopoietic mutations in the zebrafish. Development (Cambridge, England) 123, 303–309 (1996).
Golub, R. & Cumano, A. Embryonic hematopoiesis. Blood cells, molecules & diseases 51, 226–231, 10.1016/j.bcmd.2013.08.004 (2013).
Frame, J. M., McGrath, K. E. & Palis, J. Erythro-myeloid progenitors: “definitive” hematopoiesis in the conceptus prior to the emergence of hematopoietic stem cells. Blood cells, molecules & diseases 51, 220–225, 10.1016/j.bcmd.2013.09.006 (2013).
Baron, M. H., Vacaru, A. & Nieves, J. Erythroid development in the mammalian embryo. Blood cells, molecules & diseases 51, 213–219, 10.1016/j.bcmd.2013.07.006 (2013).
Li, H. et al. Lysophosphatidic acid acts as a nutrient-derived developmental cue to regulate early hematopoiesis. The EMBO journal 33, 1383–1396, 10.15252/embj.201387594 (2014).
Evseenko, D. et al. Lysophosphatidic acid mediates myeloid differentiation within the human bone marrow microenvironment. PloS one 8, e63718, 10.1371/journal.pone.0063718 (2013).
Chiang, C. L. et al. Lysophosphatidic acid induces erythropoiesis through activating lysophosphatidic acid receptor 3. Stem cells (Dayton, Ohio) 29, 1763–1773, 10.1002/stem.733 (2011).
Tonkonow, B. L. et al. Differing responses of globin and glycophorin gene expression to hemin in the human leukemia cell line K562. Blood 59, 738–746 (1982).
Testa, U. et al. Hemoglobin expression in clones of K562 cell line. European journal of biochemistry/FEBS 121, 649–655 (1982).
Yao, C. L. et al. Characterization of serum-free ex vivo-expanded hematopoietic stem cells derived from human umbilical cord blood CD133(+) cells. Stem cells and development 15, 70–78, 10.1089/scd.2006.15.70 (2006).
Flygare, J. & Karlsson, S. Diamond-Blackfan anemia: erythropoiesis lost in translation. Blood 109, 3152–3154, 10.1182/blood-2006-09-001222 (2007).
Lai, S. L. et al. Autotaxin/Lpar3 signaling regulates Kupffer’s vesicle formation and left-right asymmetry in zebrafish. Development (Cambridge, England) 139, 4439–4448, 10.1242/dev.081745 (2012).
Ganis, J. J. et al. Zebrafish globin switching occurs in two developmental stages and is controlled by the LCR. Developmental biology 366, 185–194, 10.1016/j.ydbio.2012.03.021 (2012).
Willett, C. E., Kawasaki, H., Amemiya, C. T., Lin, S. & Steiner, L. A. Ikaros expression as a marker for lymphoid progenitors during zebrafish development. Developmental dynamics: an official publication of the American Association of Anatomists 222, 694–698, 10.1002/dvdy.1223 (2001).
Chen, A. T. & Zon, L. I. Zebrafish blood stem cells. Journal of cellular biochemistry 108, 35–42, 10.1002/jcb.22251 (2009).
Burns, C. E. et al. Isolation and characterization of runxa and runxb, zebrafish members of the runt family of transcriptional regulators. Experimental hematology 30, 1381–1389 (2002).
Kalev-Zylinska, M. L. et al. Runx1 is required for zebrafish blood and vessel development and expression of a human RUNX1-CBF2T1 transgene advances a model for studies of leukemogenesis. Development (Cambridge, England) 129, 2015–2030 (2002).
Thompson, M. A. et al. The cloche and spadetail genes differentially affect hematopoiesis and vasculogenesis. Developmental biology 197, 248–269, 10.1006/dbio.1998.8887 (1998).
Jin, H. et al. Definitive hematopoietic stem/progenitor cells manifest distinct differentiation output in the zebrafish VDA and PBI. Development (Cambridge, England) 136, 647–654, 10.1242/dev.029637 (2009).
Binder, B. Y., Genetos, D. C. & Leach, J. K. Lysophosphatidic acid protects human mesenchymal stromal cells from differentiation-dependent vulnerability to apoptosis. Tissue engineering. Part A 20, 1156–1164, 10.1089/ten.TEA.2013.0487 (2014).
Pitson, S. M. & Pebay, A. Regulation of stem cell pluripotency and neural differentiation by lysophospholipids. Neuro-Signals 17, 242–254, 10.1159/000231891 (2009).
Ho, Y. H. et al. Opposing regulation of megakaryopoiesis by LPA receptors 2 and 3 in K562 human erythroleukemia cells. Biochimica et biophysica acta 1851, 172–183, 10.1016/j.bbalip.2014.11.009 (2015).
Okusa, M. D. et al. Selective blockade of lysophosphatidic acid LPA3 receptors reduces murine renal ischemia-reperfusion injury. American journal of physiology. Renal physiology 285, F565–574, 10.1152/ajprenal.00023.2003 (2003).
Furuta, D., Yamane, M., Tsujiuchi, T., Moriyama, R. & Fukushima, N. Lysophosphatidic acid induces neurite branch formation through LPA3. Molecular and cellular neurosciences 50, 21–34, 10.1016/j.mcn.2012.03.006 (2012).
Markiewicz, W., Kaminska, K., Bogacki, M., Maslanka, T. & Jaroszewski, J. Participation of analogues of lysophosphatidic acid (LPA): oleoyl-sn-glycero-3-phosphate (L-alpha-LPA) and 1-oleoyl-2-O-methyl-rac-glycerophosphothionate (OMPT) in uterine smooth muscle contractility of the pregnant pigs. Polish journal of veterinary sciences 15, 635–643 (2012).
Buckner, F. S., Eschbach, J. W., Haley, N. R., Davidson, R. C. & Adamson, J. W. Hypertension following erythropoietin therapy in anemic hemodialysis patients. American journal of hypertension 3, 947–955 (1990).
Glaspy, J. A. Erythropoietin in cancer patients. Annual review of medicine 60, 181–192, 10.1146/annurev.med.60.050307.110718 (2009).
Blau, C. A. Erythropoietin in cancer: presumption of innocence? Stem cells (Dayton, Ohio) 25, 2094–2097, 10.1634/stemcells.2007-0229 (2007).
Yasuda, Y. et al. Erythropoietin regulates tumour growth of human malignancies. Carcinogenesis 24, 1021–1029, 10.1093/carcin/bgg060 (2003).
Dicato, M. & Plawny, L. Erythropoietin in cancer patients: pros and cons. Current opinion in oncology 22, 307–311, 10.1097/CCO.0b013e32833aa9de (2010).
de Lacerda, J. F., Oliveira, S. N. & Ferro, J. M. Chronic myeloproliferative diseases. Handbook of clinical neurology 120, 1073–1081, 10.1016/b978-0-7020-4087-0.00072-3 (2014).
Yamaguchi, H., Ishii, E., Tashiro, K. & Miyazaki, S. Role of umbilical vein endothelial cells in hematopoiesis. Leukemia & lymphoma 31, 61–69, 10.3109/10428199809057585 (1998).
Bendall, L. J. & Bradstock, K. F. G-CSF: From granulopoietic stimulant to bone marrow stem cell mobilizing agent. Cytokine & growth factor reviews 25, 355–367, 10.1016/j.cytogfr.2014.07.011 (2014).
Lin, C. I., Chen, C. N., Chen, J. H. & Lee, H. Lysophospholipids increase IL-8 and MCP-1 expressions in human umbilical cord vein endothelial cells through an IL-1-dependent mechanism. Journal of cellular biochemistry 99, 1216–1232, 10.1002/jcb.20963 (2006).
Wang, H. et al. Effect of the LPA-mediated CXCL12-CXCR4 axis in the tumor proliferation, migration and invasion of ovarian cancer cell lines. Oncology letters 7, 1581–1585, 10.3892/ol.2014.1926 (2014).
Kiss, G. N. et al. Virtual screening for LPA2-specific agonists identifies a nonlipid compound with antiapoptotic actions. Molecular pharmacology 82, 1162–1173, 10.1124/mol.112.079699 (2012).
Wang, B. J. et al. Establishment of a cell-free bioassay for detecting dioxin-like compounds. Toxicology mechanisms and methods 23, 464–470, 10.3109/15376516.2013.781254 (2013).
Metscher, B. D. & Ahlberg, P. E. Zebrafish in context: uses of a laboratory model in comparative studies. Developmental biology 210, 1–14, 10.1006/dbio.1999.9230 (1999).
Iuchi, I. & Yamamoto, M. Erythropoiesis in the developing rainbow trout, Salmo gairdneri irideus: histochemical and immunochemical detection of erythropoietic organs. The Journal of experimental zoology 226, 409–417, 10.1002/jez.1402260311 (1983).
Detrich, H. W., 3rd et al. Intraembryonic hematopoietic cell migration during vertebrate development. Proceedings of the National Academy of Sciences of the United States of America 92, 10713–10717 (1995).
Leung, A. Y. et al. Characterization of expanded intermediate cell mass in zebrafish chordin morphant embryos. Developmental biology 277, 235–254, 10.1016/j.ydbio.2004.09.032 (2005).
Acknowledgements
This research was supported by grants (NSC 100-2325-B-002-045, NHRI 101-EX102-10130BI, NTU 102R76263A) to H. Lee and grants (MOST 101-2221-E-155 -044 -MY3) to CL. Yao from the National Science Council/Ministry of Science and Technology, National Health Research Institutes and National Taiwan University of the Republic of China and grants from NIAID AI080405 (GT), the NCI CA092160 (GT).
Author information
Authors and Affiliations
Contributions
K.-H.L., C.-L.Y. and H.L. convinced the study. K.-H.L. and Y.-H.H. wrote the main manuscript. Y.-H.H. and J.-C.C. prepared Figures 1, 2 and 7. K.-H.L. and Y.-J.Y. prepared Figure 3. M.-W.L. prepared Figures 4 and 5. S.-H.L. prepared Figure 6. W.-M.C., C.-L.C., Y.-N.L., C.-N.C. and G.T. provided the GRI compound helped with the experimental design and writing of the manuscript. J.L. provide the stem cell source and C.-J.H. provide zebrafish for experiments. All authors read and approved the final manuscript.

The role of LPA GPCR in RBC differentiation of the K562 cell line.
(a–c) Knockdown of LPAR2 using lentiviral shRNA and (d–f) knockdown of LPAR3 by siRNA. CD71-FITC-conjugated and GlyA-PE-conjugated antibodies were used to stain and analyze for the expression of CD71 and GlyA. The geometric mean of fluorescence was used to quantify the results. The quantitative data presented as the mean ± SD of at least three independent experiments. *p < 0.05 and **p < 0.01 indicate significant differences compared with vehicle control.

Pharmacological treatment with different LPA receptor agonists and antagonists regulates erythropoiesis in K562 cells.
K562 cells were treated, either in the presence (indicated as black bars) or absence of hemin (indicated as white bars), plus (a) LPA2 agonist DMP at 5 μM and 10 μM or (f) LPA3 agonist 2S-OMPT at 80 nM and 100 nM. The mRNA expression levels of γ-globin were analyzed by qPCR. K562 cells were treated with (b–e) LPA2 agonist DMP at 5 μM and 10 μM and (g–j) LPA3 agonist 2S-OMPT at 80 nM and 100 nM. Fluorescence intensity of CD71 (b,c,g,h) and GlyA (d,e,i,j) staining was quantified by Flow cytometry. Data are represent the mean ± SD of three independent experiments. *p < 0.05, **p < 0.01 and ***p < 0.005 indicate significant differences compared with vehicle control.

Pharmacological treatment with LPA2 and LPA3 agonists of CD34+ hHSC.
Umbilical blood-derived CD34+ hHSC were cultured with GRI and 2S-OMPT separately for six days under EPO induction. (a,b) The cells were harvested and the expression of CD71 and GlyA RNA was analyzed using qPCR. (c,d) The CD71 protein expression was analyzed by flow cytometry. Data are the mean ± SD of at least three independent experiments. *p < 0.05, *p < 0.01 and ***p < 0.005 indicate significant differences compared with vehicle control.

Characterization of the function of LPA3 during primitive erythropoiesis in zebrafish embryos.
Zebrafish embryos were immersed in (a) the LPA1/3 antagonist Ki16425 or (b) the LPA3 agonist 2S-OMPT for 48 h. O-dianisidine staining was classified as normal, moderate, or severe based on the expression level of embryonic blood in the CHT region. The right-hand panel shows the quantitative results. (c) Whole-mount in situ hybridization showed the mRNA expression of α-A1, β-E1 globin and ikaros in zLPAR3 tMO-injected or wild type embryos.

Analysis of the roles of LPA3 during definitive erythropoiesis in zebrafish.
(a) O-dianisidine staining showed the effects of LPAR3 mRNA, 2S-OMPT and LPAR3-MO during definitive erythropoiesis at 96 hpf in the CHT region. (b) The quantitative results for (a). (c) A whole-mount in situ hybridization showing the mRNA expression levels of α-E1 and β-E1 in the wild-type, LPAR3-MO and 2S-OMPT treatment groups.

Characterization of the effects of LPA2 during definitive erythropoiesis.
The effects of LPAR2 MO revealed defective erythropoiesis in the CHT region based on o-dianisidine staining. (b) The quantitative results for (a). (c) The effects of the LPA2 agonist GRI after treatment for 96 h. O-dianisidine staining levels were classified as normal, moderate, or defective based on the expression levels in the embryonic blood. (d) The quantification hemoglobin expression after treatment with different concentrations of GRI for four days.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Lin, KH., Ho, YH., Chiang, JC. et al. Pharmacological activation of lysophosphatidic acid receptors regulates erythropoiesis. Sci Rep 6, 27050 (2016). https://doi.org/10.1038/srep27050
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep27050
This article is cited by
-
The synergistic and enhancive effects of IL-6 and M-CSF to expand and differentiate functional dendritic cells from human monocytes under serum-free condition
Journal of Biological Engineering (2023)
-
An Aligned Poly(3-hydroxybutyrate-co-3-hydroxyvalerate) Scaffold Fixed with Fibronectin to Enhance the Attachment and Growth of Human Endothelial Progenitor Cells
Biotechnology and Bioprocess Engineering (2023)
-
Co-stimulation of LPAR1 and S1PR1/3 increases the transplantation efficacy of human mesenchymal stem cells in drug-induced and alcoholic liver diseases
Stem Cell Research & Therapy (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
