
Abstract
Momilactones, which are diterpenoid phytoalexins with antimicrobial and allelopathic functions, have been found only in rice and the moss Hypnum plumaeforme. Although these two evolutionarily distinct plant species are thought to produce momilactones as a chemical defence, the momilactone biosynthetic pathway in H. plumaeforme has been unclear. Here, we identified a gene encoding syn-pimara-7,15-diene synthase (HpDTC1) responsible for the first step of momilactone biosynthesis in the moss. HpDTC1 is a bifunctional diterpene cyclase that catalyses a two-step cyclization reaction of geranylgeranyl diphosphate to syn-pimara-7,15-diene. HpDTC1 transcription was up-regulated in response to abiotic and biotic stress treatments. HpDTC1 promoter-GUS analysis in transgenic Physcomitrella patens showed similar transcriptional responses as H. plumaeforme to the stresses, suggesting that a common response system to stress exists in mosses. Jasmonic acid (JA), a potent signalling molecule for inducing plant defences, could not activate HpDTC1 expression. In contrast, 12-oxo-phytodienoic acid, an oxylipin precursor of JA in vascular plants, enhanced HpDTC1 expression and momilactone accumulation, implying that as-yet-unknown oxylipins could regulate momilactone biosynthesis in H. plumaeforme. These results demonstrate the existence of an evolutionarily conserved chemical defence system utilizing momilactones and suggest the molecular basis of the regulation for inductive production of momilactones in H. plumaeforme.
Similar content being viewed by others

Introduction
The moss Hypnum plumaeforme (Hypnaceae) is widely distributed in East Asia; it prefers sunlit locations and forms large homogeneous colonies on dry ground and rocks. H. plumaeforme has potent allelopathic activity, suppressing the growth of surrounding plants1. Momilactones, originally discovered in rice husks as allelochemicals and phytoalexins, are specialized metabolites that repress growth of neighbouring plants and defend against pathogens2,3. Momilactones accumulate in rice in response to both abiotic and biotic stresses, such as heavy metal treatment, ultra violet (UV) irradiation and infection by the rice blast fungus4,5,6,7,8. Other than rice, the moss H. plumaeforme is the only plant species known to produce momilactones. We previously found that momilactones function as allelochemicals in this moss1. As a defence response to infection by pathogens such as Botrytis cinerea, the moss Physcomitrella patens exhibits fortification of the plant cell walls via incorporation of phenolic compounds9. Momilactones in H. plumaeforme accumulate in response to various stresses, including UV irradiation and heavy metal treatment10. These studies suggest that momilactone production could be a defence reaction in H. plumaeforme, in addition to providing allelopathic activity against other plants.
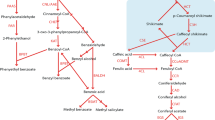
The biosynthetic pathway of momilactones and its corresponding genes have been extensively investigated in rice because of the similarity to the biosynthetic pathway of gibberellins (GAs), tetracyclic diterpene plant hormones (Fig. 1). In angiosperms, the cyclic diterpene skeleton, found in metabolites such as ent-kaurene, a precursor of GA, is derived from geranylgeranyl diphosphate (GGDP) by two different cyclases, ent-coparyl diphosphate synthase (ent-CPS) that belongs to type B cyclase and ent-kaurene synthase (KS) catalysing the type A cyclization of ent-copalyl diphosphate (ent-CDP). Rice also produces the cyclic diterpene skeletons of phytoalexins, momilactones, oryzalexins and phytocassanes from GGDP by two different type B and A cyclases. In rice, syn-copalyl diphospate (syn-CDP) synthase (OsCPS4) and syn-pimara-7,15-diene synthase (OsKSL4) are involved in momilactone biosynthesis11. A key intermediate in the momilactone biosynthetic pathway, syn-pimara-7,15-diene, is further oxidized by cytochrome P450 enzymes and dehydrogenase to yield the active substances, that is, momilactones A and B12,13. The expression of all genes in momilactone biosynthesis is coordinately induced in response to both biotic and abiotic stresses, including blast fungus infection and heavy metal treatment4,14.

The biosynthetic pathway of diterpenoids in plants.
Proposed biosynthetic pathway for momilactones in rice. Both syn-pimara-7,15-dine and ent-kaurene, the key intermediates for momilactone and gibberellin, respectively, are synthesized from geranylgeranyl diphosphate (GGDP) by sequential enzymatic reactions of type B and type A cyclases. In mosses and liverworts, cyclic diterpenes, such as ent-kaurene, are constructed by a bifunctional cyclase with type B and A activity in a single polypeptide.
In contrast to vascular plants, the moss P. patens produces ent-kaurene via a bifunctional ent-kaurene synthase, PpCPS/KS, that is similar to fungal bifunctional ent-kaurene synthases15. PpCPS/KS has both type B and A activity in a single polypeptide and is the sole diterpene cyclase in the P. patens genome (Fig. 1)15. To date, other diterpene cyclase genes have not been identified from the mosses. Additionally, the molecular basis for the biosynthesis and physiological function of phytoalexins and allelochemicals in the moss has not been demonstrated. Therefore, the discovery of biosynthesis genes of momilactones in the moss would shed light on the ancient defence system of early land plants.
To dissect the biosynthetic pathway of momilactone and its regulatory mechanism in response to stress, we cloned the gene for syn-pimaradiene synthase (HpDTC1, H. plumaeforme diterpene cyclase 1), a key first step enzyme in momilactone biosynthesis. The HpDTC1 gene encodes a bifunctional enzyme catalysing two sequential cyclization steps from GGDP to syn-pimara-7-15-diene via syn-CDP as the reaction intermediate. HpDTC1 showed inductive expression under treatments with CuCl2, chitosan and pathogen infection. Similar to HpDTC1transcriptional regulation in H. plumaeforme, the findings for transgenic P. patens, expressing an HpDTC1 promoter-GUS construct (HpDTC1::GUS), demonstrated that the HpDTC1 promoter is up-regulated by both abiotic and biotic stress treatments. An allelopathy activity assay showed that H. plumaeforme is highly resistant to extracts from H. plumaeforme and momilactone B at the active concentration for other types of mosses and liverworts. Additionally, the extract from H. plumaeforme showed phytoalexin activity against B. cinerea and protected the gametophore from infection by the pathogen. Our results aid in understanding the momilactone biosynthetic pathway in the moss and the molecular basis of an evolutionarily conserved chemical defence system between mosses and angiosperms utilizing momilactones.
Results
Identification of H. plumaeforme diterpene cyclase that is responsible for momilactone synthesis
Production of momilactones in H. plumaeforme has been reported to be highly induced by abiotic stresses, such as UV irradiation and heavy metals10. A key intermediate for momilactone biosynthesis in rice is syn-pimara-7,15-diene produced from GGDP. OsKSL4, a syn-pimara-7,15-diene synthase, is also induced by biotic and abiotic stresses in rice12. To identify the syn-pimaradiene synthase in H. plumaeforme, we screened for homologous genes of P. patens CPS/KS using next-generation sequencing of the cDNA library from sterile cultures of H. plumaeforme gametophores treated with CuCl2. We identified several CuCl2-upregulated contigs having homology to PpCPS/KS and gymnosperm bifunctional diterpene cyclases; one gene, designated HpDTC1, was the only diterpene cyclase gene in H. plumaeforme that was induced by CuCl2. Therefore, HpDTC1 cDNA was amplified with RACE-PCR from a cDNA library from H. plumaeforme gametophores. The full-length HpDTC1 cDNA was determined to be 2786 bp, encoding an 856 amino acid polypeptide (DDBJ ID: LC128408) as shown in Fig. 2. The N-terminal region (about 85 amino acids) was putatively assigned as a transit peptide region. The deduced HpDTC1 contained the three characteristic motifs for diterpene cyclases, SAYDTAWVA, DIDD and DDLFD motifs (Fig. 2). Both active site motifs for type B (DxDD) and type A (DDxxD) cyclases indicated that HpDTC1 would function as a bifunctional diterpene cyclase similar to PpCPS⁄KS, gymnosperm diterpene synthase and fungal CPS⁄ KS (see Supplementary Fig. S1). HpDTC1 showed identity to PpCPS⁄KS (43%), JsCPS/KS (41%), Selaginella moellendorffii diterpene cyclases (DTCs) (38–41%), gymnosperm bifunctional DTCs (40–43%), plant CPSs (43–45%) and angiosperm KS and KS-like DTCs (30–32%). Phylogenetic analysis indicated that HpDTC1 was most closely related to the gymnosperm bifunctional diterpene cyclase (see Supplementary Fig. S2).

Amino acid sequence of HpDTC1 and active site motifs, DIDD and DDLFD.
Three diterpene-specific motifs (SxYDTAWVA, DxDD and DDxxD) are underlined. The DxDD motif in type B cyclase is responsible for the cyclization activity of GGDP to form CDP as a product. The DDxxD in type A cyclase is responsible for the cyclization of CDP to yield a cyclic diterpene hydrocarbon. Type B-A cyclase, HpDTC1, has two cyclization motifs in a single polypeptide and functions as a bifunctional cyclase with both type B and A cyclization activity.
Functional characterization of HpDTC1 diterpene cyclase
HpDTC1 showed two aspartate-rich motifs (DxDD and DDxxD) for active sites responsible for the sequential cyclization reaction of bifunctional diterpene cyclase16,17. ent-Kaurene synthase, PpCPS/KS, was the sole diterpene synthase gene identified from the mosses. PpCPS/KS is a bifunctional diterpene cyclase that catalyses the direct cyclization reaction of GGDP to yield ent-kaurene15. HpDTC1 is predicted to catalyse the cyclization of GGDP to give a tricyclic diterpene product via syn-CDP as the reaction intermediate. The recombinant HpDTC1 enzyme was expressed in E. coli as a His6-tag fusion protein and used for the enzyme assay using GGDP as a substrate. GC-MS analysis of the reaction products of HpDTC1 showed a single product peak on a GC-MS chromatogram and the MS spectrum and retention time (Rt = 12.52 min) of the HpDTC1 product were identical to those of an authentic sample of syn-pimara-7,15-diene (Fig. 3a,b). This result demonstrated that HpDTC1 functions as bifunctional syn-pimaradiene synthase. In rice, diterpene phytoalexins, that is, momilactones, are biosynthesized from syn-CDP; in contrast, similar diterpene phytoalexins, that is, oryzalexins and phytocassanes, are synthesized from ent-CDP produced by OsCPS1/211. To confirm the reaction intermediate of HpDTC1, we performed a mutational study on the HpDTC1 enzyme. The first aspartate-rich motif, DIDD, is required for the cyclization of GGDP to yield CDP (type B activity), whereas the second motif, DDLFD, is essential for the cyclization of CDP (type A activity)18. The loss-of-function mutation related to type A activity of HpDTC1 (DDLFD to GGLFD) resulted in the accumulation of an intermediate syn-CDP that was identical to syn-CDP produced by OsCPS4, a syn-CDP synthase (Fig. 3c,d; see Supplementary Fig. S3). The mutation in the first motif, DIDD (to DIGG), for type B activity disrupted the initial cyclase activity for GGDP in HpDTC1. Co-incubation with OsCPS4 and the type B mutant of HpDTC1 (DIGG) complemented the impaired type B activity of the DIGG mutant to produce syn-pimara-7,15-diene as the product, similar to the HpDTC1 wild type enzyme (Fig. 3e,f; see Supplementary Fig. S3). These findings clearly demonstrate that HpDTC1 is a bifunctional diterpene cyclase utilizing syn-CDP as a reaction intermediate.

HpDTC1 is a bifunctional syn-pimaradiene synthase.
GC-MS chromatograms of enzymatic products. (a) HpDTC1 product and (b) authentic sample of syn-pimara-7,15-diene. (c) syn-copalol derived from syn-CDP, product of OsCPS4. (d) The product of the HpDTC1-GGLFD mutant enzyme was identical to syn-copalol. (e) syn-pimara-7,15-diene, HpDTC1 product. (f) The product from the co-reaction of OsCPS4 and HpDTC1-DIGG enzyme was identical to syn-pimara-7,15-diene. The selected ion was monitored at m⁄ z 272 (a,b,e,f) or m/z 275 (c,d) to detect cyclized diterpenes. The reaction product, syn-CDP in (c,d) was detected as alcohol after dephosphorylation by calf intestinal alkaline phosphatase.
HpDTC1 expression induced by both abiotic and biotic stresses correlates with the regulation of momilactone biosynthesis
In rice, momilactone biosynthesis is elevated in response to various biotic and abiotic stresses. Consistent with the induction of momilactone biosynthesis, the expression of syn-pimaradiene synthase (OsKSL4) was highly induced by these stresses11,19. H. plumaeforme also accumulated momilactones in response to abiotic stresses, such as UV irradiation and heavy metals. To examine the abiotic stress-induced expression of HpDTC1, we assessed HpDTC1 transcriptional regulation and accumulation of momilactones in the gametophore under CuCl2 treatment. The gametophore culture was incubated in 500 μM CuCl2 and then the HpDTC1 transcript level was determined at regular intervals by quantitative RT-PCR (qRT-PCR). HpDTC1 transcript level was normalized to the transcript level of HpACT3, a housekeeping actin gene. Momilactones A and B in the gametophore were quantified by liquid chromatography with electrospray ionization tandem mass spectrometry (LC-MS/MS).
As shown in Fig. 4a, abiotic stress by CuCl2 treatment dramatically induced HpDTC1 expression within 6 h and the expression remained at a high level for 48 h after the treatment. Consistent with this HpDTC1 induction, momilactones A and B also accumulated to a high level with CuCl2 treatment (Fig. 4e). Similar to the induction of momilactone synthesis in rice, H. plumaeforme was reported to induce momilactone production in response to jasmonic acid (JA), a signalling molecule involved in the defence response of vascular plants10. However, under our experimental conditions for the cultured gametophores, enhancement of accumulation of momilactones was barely detected with JA treatment in the sterile culture of H. plumaeforme gametophores (Fig. 4f). Consistent with the lack of response of momilactone production to JA, HpDTC1 expression showed almost negligible change with JA treatment (Fig. 4b). 12-Oxo-phytodienoic acid (OPDA), a biosynthetic precursor of JA, functions in activation of the defence response in the moss9,20. Thus, we also examined the effect of OPDA on the expression of HpDTC1 and production of momilactones. As shown in Fig. 4b, we found that OPDA clearly induced HpDTC1 gene expression at 6 h after the treatment and increased accumulation of momilactones was also detected 72 h after the treatment. Taken together, the response of HpDTC1 expression and momilactone production to CuCl2 and OPDA treatment suggests that HpDTC1 expression, triggered by these abiotic stresses, correlates well with momilactone accumulation.

HpDTC1 expression and momilactone production is induced by abiotic and biotic stresses.
(a–d) qRT-PCR was performed using HpDTC1 specific primers on total RNA from H. plumaeforme gametophores treated with (a) CuCl2 (500 μM), (b) OPDA, 12-oxo phytodienoic acid, (100 μM) and jasmonic acid (100 μM), (c) chitosan (500 μg/mL) for the indicated times and (d) the pathogenic fungus, Botrytis cinerea. The fungus B. cinerea was inoculated onto BCDATG agar plates of H. plumaeforme gametophores and then incubated for the indicated number of days. As an internal standard, the H. plumaeforme actin gene, HpACT3, was amplified by qRT-PCR and then the HpDTC1 transcription level was normalized against the HpACT3 level. (e–h) Momilactones A and B in H. plumaeforme gametophores treated with stresses were measured by LC-MS/MS. Data are presented as mean ± standard error. (a–c,e–h) n = 3, (d) n = 4, *p < 0.05, **p < 0.01.
We next examined the effects of biotic stress on HpDTC1 expression. Biotic elicitors activate the defence system of rice and induce the expression of diterpene cyclase genes for phytoalexins, leading to the accumulation of diterpene phytoalexins, including momilactones21,22. Chitosan is also reported to induce defence-related responses in the moss23. Therefore, H. plumaeforme gametophores were incubated with chitosan. In addition, B. cinerea, a necrotrophic plant pathogenic fungus, infects the gametophores of moss plants and causes disease symptoms9. B. cinerea was inoculated onto H. plumaeforme gametophores and then incubated for 6 days (see Supplementary Fig. S4). HpDTC1 expression levels in the chitosan-treated or B. cinerea-infected gametophores were analysed by qRT-PCR. Both chitosan treatment and B. cinerea infection up-regulated HpDTC1 expression (Fig. 4c,d) and accumulation of momilactones was significantly induced by these biotic stresses (Fig. 4g,h). These results suggest that the HpDTC1 gene is stress-responsive not only to abiotic stress, but also to biotic stress and the expression of HpDTC1 is part of the defence response through the production of phytoalexin momilactones in H. plumaeforme.
HpDTC1 expression is regulated by a common stress signalling pathway conserved in moss plants
To assess whether the stress signalling pathway regulating HpDTC1 expression is commonly conserved in other mosses, a GUS reporter assay for the HpDTC1 promoter was performed in the model moss P. patens. The promoter region of HpDTC1 was amplified by TAIL-PCR from genomic DNA and 3.4 kbp of the HpDTC1 promoter was inserted into the GUS reporter plasmid, pPIG1bNGGII. The HpDTC1promoter::GUS construct (HpDTC1::GUS) was used to transform P. patens protoplasts by a conventional PEG-mediated method. The transgenic HpDTC1::GUS line showed very faint GUS basal expression in gametophores, similar to the control line (transformed with a promoter-less pPIG1b-NGGII vector) (Fig. 5a). In contrast, the protonema cells showed significant basal GUS expression compared to the control line (see Supplementary Fig. S5). The treatment of protonema cells with CuCl2 and chitosan did not strongly induce HpDTC1::GUS reporter expression (see Supplementary Fig. S5). However, consistent with HpDTC1 expression in H. plumaeforme, the treatment of gametophores with CuCl2 dramatically induced HpDTC1::GUS expression in P. patens (Fig. 5a,b). Chitosan and its oligomers, functioning as elicitors and the chitosan-inducible peroxidase, Prx34, play a crucial role in the basal defence system against fungal invasion in P. patens23. Consistent with the elicitation of the P. patens defence response with chitosan, the treatment with chitosan and its oligomer greatly upregulated GUS reporter expression in gametophores (Fig. 5a,b). HpDTC1::GUS induction reached its maximum at 24 h after exposure to these stress conditions (Fig. 5c). However, jasmonic acid had little effect on HpDTC1::GUS expression in P. patens, similar to H. plumaeforme (Fig. 5a,b). In contrast, OPDA treatment weakly but clearly induced HpDTC1 promoter activity, as seen in the expression of HpDTC1 in OPDA-treated H. plumaeforme (see Supplementary Fig. S6), suggesting that OPDA plays a role in common defence signalling in the mosses.

The HpDTC1 promoter is regulated by abiotic and biotic stress responses in the moss Physcomitrella patens.
(a) HpDTC1 promoter::GUS (HpDTC1::GUS) expression was histochemically visualized after 24 h stress treatment. The HpDTC1::GUS reporter was highly induced by CuCl2 (500 μM), chitosan (250 μg/ml) and chitosan oligomers (Average Mw = 4000–6000, 250 μg/ml), but not by jasmonate, JA, (100 μM) on leafy gametophores (upper panel). The vector control line did not respond to the stress treatments (lower panel). Scale bars: 1 mm. The quantitative data for HpDTC1::GUS reporter induction at 24 h treatment (b) by CuCl2 (500 μM), chitosan (250 μg/ml), chitosan oligomers (250 μg/ml) and JA (100 μM) and at the indicated time (c) by CuCl2 (500 μM) and chitosan (250 μg/ml). The GUS activity of HpDTC1::GUS gametophores was fluorometrically determined. Data are presented as mean ± standard deviation of biological replicates (n = 5 for (b), n = 4 for (c); *p < 0.001).
The response of HpDTC1 promoter expression under biotic stress was also examined with fungal infection; B. cinerea24 or Pythium irregulare25 was inoculated onto a H. plumaeforme colony and then cultured for 4 days. Consistent with the induction of HpDTC1 in H. plumaeforme, HpDTC1::GUS expression was highly induced by fungal infection of both B. cinerea (Fig. 6a–c) and P. irregulare (see Supplementary Fig. S7), while GUS activity was not observed in the vector control line. Additionally, sterilized mycelia of B. cinerea also activated HpDTC1::GUS expression, suggesting an elicitor component from the mycelia would activate the defence response in P. patens (Fig. 6d). This evidence suggests that a common defence signalling pathway, conserved among mosses, would function in defence-related gene expression, including HpDTC1 in H. plumaeforme.

The HpDTC1 promoter is induced by Botrytis cinerea infection in the moss Physcomitrella patens.
(a,b) The pathogenic fungus, B. cinerea, was inoculated onto colonies of HpDTC1::GUS and vector control lines and then the culture plate was incubated for 4 days at 24 °C. The HpDTC1::GUS plants were histochemically stained. Scale bars: 1 mm. (c) The quantitative data for HpDTC1::GUS reporter induction by B. cinerea infection after 4 days incubation. The GUS activity was fluorometrically determined. Data are presented as mean ± standard deviation of biological replicates. (n = 5–8, *p < 0.01.). (d) Application of sterilized mycelia of B. cinerea induced HpDTC1::GUS reporter expression. The mycelia were placed on a HpDTC1::GUS colony and then incubated for 4 days at 24 °C.
Physiological role of momilactones in defence response of H. plumaeforme
Momilactones are exuded as allelopathic substances from the roots of rice3. The moss H. plumaeforme forms large homogeneous colonies in dry areas. Momilactones could be accumulated as allelopathic substances in the gametophores to prevent the invasion of the moss colony by competing plant species. To confirm the allelopathic role of momilactones in H. plumaeforme, the plant growth inhibitory activity of the extract from the gametophore was measured (Fig. 7a–f; see Supplementary Fig. S8). The extract from stress-induced gametophores (Fig. 7d,e) showed more potent growth inhibition on P. patens and M. polymorpha than the control extract (Fig. 7a,b). In contrast, H. plumaeforme was highly resistant to both extracts at 200 μg/ml (Fig. 7c,f). Additionally, pure momilactone B showed no effects on the growth of H. plumaeforme at 100 μM, but showed potent inhibition on the growth of M. polymorpha at 5 μM (see Supplementary Fig. S9). This evidence supports the allelopathic role of momilactones in the formation of large homogeneous colonies of H. plumaeforme.

Physiological function of momilactone produced by H. plumaeforme.
(a–f) The allelopathic effects of inductively produced momilactones on the moss P. patens (a,d), liverwort Marchantia polymorpha (b,e) and H. plumaeforme (c,f). The acetone extract was prepared from H. plumaeforme gametophores treated with or without CuCl2. The plants were cultured on BCDATG agar plates containing the extract at the indicated concentration and growth recorded at the indicated times. Data are presented as mean ± standard deviation of biological replicates (n = 15 for (a,d), n > 18 for (b,e), n = 8 for c,f). (g) Effects of H. plumaeforme extract on the growth of the pathogenic fungus, B. cinerea. The mycelia were inoculated onto the BCDATG plate containing H. plumaeforme acetone extract and the gametophores. The inoculated plate was incubated for 4 days. (h) Effects of H. plumaeforme gametophores on the growth of B. cinerea. H. plumaeforme gametophores were dried at 70 °C and then agar plates were prepared with the dead gametophores suspended in BCDATG agar (10% w/v). Fresh H. plumaeforme gametophores were placed on the agar plates; then, the mycelia were inoculated and incubated for 7 days. Arrowheads indicate the cells damaged by the pathogenic infection.
HpDTC1 expression in the moss was induced by the chitosan elicitor and B. cinerea infection, suggesting the production of momilactones is a defence response to pathogenic attack. To investigate the role of momilactone as a phytoalexin in H. plumaeforme, we examined the protective activity against B. cinerea infection of momilactone produced by H. plumaeforme. The acetone extract from the stress-induced gametophytes showed potent inhibitory activity on the growth of B. cinerea, implying momilactone plays a role as a phytoalexin of H. plumaeforme to protect from the pathogenic attack of B. cinerea (Fig. 7g). In the natural world, H. plumaeforme forms large and dense colonies composed of stacked gametophytes1. Momilactones were found in the soil covered by H. plumaeforme colonies, suggesting H. plumaeforme accumulates momilactones in response to stresses and releases them as phytoalexins to protect from pathogenic attack from the soil. The momilactones released from wounds or dead cells in the stacked gametophyte colony could play a role in the defence system of H. plumaeforme. In our experiment, living gametophytes were inoculated with B. cinerea on a medium containing stress-treated dead gametophytes (Fig. 7h; see Supplementary Fig. S10). The application of stress-treated dead gametophytes to the medium repressed the growth of B. cinerea and reduced the damage of H. plumaeforme by the infection (Fig. 7h), implying that the stacked gametophyte colony could play a role in the chemical defence system of moss against the pathogenic fungus. These results support the physiological function of momilactones as both phytoalexins and allelopathic substances in the defence response of H. plumaeforme.
Discussion
In this study, we identified a stress-inducible H. plumaeforme diterpene cyclase gene, HpDTC1, encoding syn-pimaradiene synthase, which catalyses key steps of momilactone biosynthesis to yield syn-pimara-7,15-diene from GGDP via syn-CDP as the reaction intermediate. HpDTC1 is a bifunctional enzyme consisting of type B cyclase that is responsible for syn-CDP synthesis and type A activity for syn-pimaradiene synthesis. In angiosperms, cyclic diterpenes such as ent-kaurene are key intermediates of gibberellin biosynthetic pathways and syn-pimaradiene is a precursor for momilactone. These precursors are synthesized by the successive reactions of monofunctional type B and type A cyclases11,19,26. In gymnosperms and lycophytes, ent-kaurene is synthesized by two monofunctional diterpene cyclases, ent-CPS (type B) and KS (type A)27, whereas other cyclic diterpenes for secondary metabolites, such as miltiradiene, pimaradiene, levopimaradiene and abietadiene, are produced by bifunctional type B-A diterpene cyclases28,29. In contrast, the ent-kaurene synthase, PpCPS/KS, in the moss has been identified as a bifunctional type B-A cyclase that is involved in the biosynthesis of diterpene-type growth regulators for protonema development in P. patens14. PpCPS/KS is the sole diterpene synthase in the moss P. patens and other DTCs have not been identified from other mosses. HpDTC1, therefore, is the first diterpene synthase identified that is responsible for the biosynthesis of specialized metabolites in the mosses. In the defence response of gymnosperms, bifunctional abietadiene synthase was induced by wound stress to produce diterpene resin acids as protective substances17. Consistent with the role of the bifunctional DTC in the gymnosperm defence system, phylogenetic analysis showed HpDTC1 is more closely related to the gymnosperm bifunctional DTCs than to moss (PpCPS/KS) and liverwort (JsCPS/KS) ent-kaurene synthases, implying HpDTC1 could be the prototype of cyclases for specialized diterpenes for defence responses in the land plants (see Supplementary Fig. S2).
Momilactone production was reported to be induced by various stresses, including UV irradiation and some chemical elicitors, such as JA, CuCl2 and phosphatase inhibitors10. We confirmed significant induction of momilactone production and HpDTC1 expression in our study in response to abiotic stress treatments, except for the JA treatment. The lack of responsiveness of momilactone synthesis to JA contradicts a previous study by Kato-Noguchi et al.10. However, it has been reported that endogenous JA is not consistently detected at physiologically active levels in mosses, including P. patens and M. polymorpha and no obvious physiological effects on plant development could be observed with JA treatment30,31. In fact, JA accumulation in wound/CuCl2-treated H. plumaeforme was undetectable by LC-MS/MS (see Supplementary Fig. S11). In contrast, we found that OPDA, a precursor of JA, has the ability to activate HpDTC1 expression and momilactone production in H. plumaeforme. Notably, OPDA has been reported to act as a signalling molecule to induce defence responses in P. patens9. These observations suggest that the active signalling molecules in moss might be OPDA or its metabolites and differ from those in vascular plants. Further evidence is needed to demonstrate how OPDA or its metabolites can activate momilactone production in H. plumaeforme.
Currently, the molecular basis of defence responses in the moss is not well understood. However, there have been some reports of defence responses in mosses. For example, Lehtonen et al. reported that type III peroxidase (Prx34), which is known as a PR protein involved in plant defence, is rapidly induced in response to chitosan treatment. Further, the knock-out of the Prx34 gene in P. patens confers the hyper-sensitive phenotype to pathogenic fungus infection23. In addition, the PR genes in P. patens, that is, those encoding phenylalanine ammonia-lyase, chalcone synthase and lipoxygenase, were up-regulated by the pathogenic attack of B. cinerea and P. irregulare30. Our results also showed that biotic stresses, such as chitosan treatment and B. cinerea infection, clearly induced both momilactone accumulation and HpDTC1 expression, suggesting that momilactone biosynthesis in H. plumaeforme is part of the defence response to pathogen attack. Our findings here in H. plumaeforme, together with previous reports in P. patens, strongly support that defence responses leading to the expression of PR genes, HpDTC1 and Prx34, are a common strategy in mosses to resist attacks by pathogens. H. plumaeforme could have evolved the ability to use momilactone synthesis as a chemical weapon against pathogens. In addition, momilactones function as allelochemicals, preventing growth of neighbouring plants. Thus, H. plumaeforme has a great growth advantage when facing biotic environmental stresses like pathogens and competition with other plants.
In our histochemical GUS staining analysis, the transgenic HpDTC1::GUS line showed GUS reporter gene induction when the gametophore was treated with CuCl2, chitosan and infection by B. cinerea. We also found that the expression of HpDTC1 has different responses in different developmental stages. Protonema cells are an early developmental stage of moss that forms a highly branched filamentous structure. When we tested GUS expression in protonema cells of HpDTC1::GUS reporter lines with treatments of CuCl2 and chitosan, in contrast to the gametophore, high basal expression and no obvious induction of the GUS reporter gene were observed in whole protonema cells (see Supplementary Fig. S12). This result is consistent with HpDTC1 expression and momilactone production in H. plumaeforme protonema cells, showing uniform momilactone production and HpDTC1 expression, even in the treatment with the chitosan elicitor (see Supplementary Fig. S5). This finding also suggests that defence-related transcriptional regulation would only be active in the gametophore. Moreover, our results indicate that the HpDTC1 promoter is active in the model moss plant, P. patens. This result indicates that regulatory mechanisms of HpDTC1 expression could be conserved among mosses, i.e., similar transcriptional regulation might function in HpDTC1 expression in both mosses.
In P. patens, oxylipins could function in the defence response and regulate plant development, but the signalling component(s), specifically transcriptional regulators in oxylipin-regulated pathways, are still not understood. For momilactone synthesis and its transcriptional regulation of biosynthetic gene expression in rice, several transcription factors regulating momilactone synthesis have been identified. A bZIP-type transcription factor, OsTGAP132,33, a bHLH-type transcription factor, DPF14 and WRKY-type transcription factors, OsWRKY53 and OsWRKY4534,35, have been shown to function as positive regulators of momilactone biosynthetic gene expression, whereas OsbZIP79 and OsWRKY76 have potential activity as negative regulators36,37. In H. plumaeforme, many types of transcription factors (ERF, bHLH and WRKY-type) were induced by CuCl2 treatment in our RNA-seq result (see Supplementary Fig. S13). However, these transcription factors did not have high similarity to rice orthologs of the regulatory factors for momilactone synthesis. It seems that different factors are involved in defence signalling leading to momilactone production in rice and H. plumaeforme (both momilactone producers), although mosses would likely possess a common system for transcriptional regulation of defence-related gene expression. Further investigation of the transactivation activity of these factors to the HpDTC1 promoter will provide possible candidates for modulating stress-responsive expression of the momilactone biosynthetic HpDTC1 gene.
Momilactones, which are inductively produced in H. plumaeforme and rice, are typical specialized metabolites for chemical defence strategies in these plants. When and how these two evolutionarily distinct species acquired the ability to produce the identical specialized diterpenes and to regulate their biosynthesis comprise one of the great mysteries of science. Further, how rice and H. plumaeforme resist allelopathic substances, such as momilactones, when the growth of many other plants is inhibited by momilactones, is an interesting question. The mode of action and the candidates for factors enabling resistance to momilactones are not known at present. Further exploration of candidate genes responsible for such resistance mechanisms will provide insight into allelopathic activity of and resistance mechanisms to momilactones. On-going whole genome analysis of H. plumaeforme might reveal a broad picture of the evolutionary steps towards the momilactone biosynthetic pathway in plants.
Methods
Plant Materials and Growth Conditions
The wild-type strain of Hypnum plumaeforme was collected at Okayama, Japan and sterilized with benzethonium chloride (0.1 w/v% aqueous solution). The sterilized gametophores were cultured on BCDATG agar medium38 under continuous white light at 24 °C. The wild-type strain of Physcomitrella patens subsp patens was cultured on BCDAT agar medium under continuous white light at 24 °C. Marchantia polymorpha was cultured on half-strength Gamborg’s B5 medium containing 8 g/L agar under continuous white light at 24 °C. For the stress treatment, media containing 8 g/L agar under continuous white light at 24 °C was used. For the stress treatment, the H. plumaeforme culture was incubated with BCDATG liquid media containing the chemicals and elicitors (jasmonic acid (Tokyo Kasei, Japan), low molecular weight chitosan (Aldrich, Japan) and chitosan oligosaccharide lactate (Aldrich, Japan)) or aqueous solution of copper (II) chloride dihydrate (Nacalai Tesque, Japan). Chitosan was dissolved in water by the addition of acetic acid and final pH of the solution was adjusted to pH 6.0. The acetone extract of H. plumaeforme was prepared from the culture of gametophores incubated with or without 500 μM CuCl2 for 96 h. The gametophore was soaked in acetone for 24 h at 20 °C and then the organic layer was evaporated in vacuo. For the growth inhibitory assay, the plants were inoculated on the corresponding agar medium containing the acetone extract in a 6-well plate and cultured under continuous white light at 24 °C. Photographs of the plant were taken at regular intervals and the growth of the plant was analysed using ImageJ software. For the stress treatment of the HpDTC1::GUS reporter line, P. patens gametophores grown on BCDAT agar media for 3 weeks were incubated in BCDATG liquid media containing the chemicals and elicitors for 24 h under continuous white light at 24 °C. For the infection experiment, the necrotrophic plant fungus, B. cinerea MAFF237501 and Pythium irregulare MAFF237249, were obtained from the GeneBank of the National Institute of Agrobiological Sciences (Japan) and grown on potato dextrose agar medium. Agar blocks (1 mm) containing the mycelia were placed on a P. patens colony or H. plumaeforme gametophore on BCDATG agar plates. The plates were incubated under continuous white light at 24 °C.
RNA-Seq
H. plumaeforme gametophores were treated with or without 500 μM CuCl2 for 8 h. Total RNA was extracted from the gametophore (500 mg) using the RNeasy Plant Mini Kit (QIAGEN, USA) according to the manufacturer’s instructions. The concentrations and qualities of RNA were determined with the Agilent Bioanalyser 2100 (Agilent Technologies, Inc., Santa Clara, CA, USA). RNA concentration was adjusted to 1 μg/μL and ratios of 28S/18S were estimated to be higher than 2.10. Total RNA (9 μg per sample) was used for the following library generation and sequencing conducted by BGI Japan.
Identification of HpDTC1 gene
The BLAST search of diterpene synthase against RNA-seq data revealed several contigs of diterpene cyclase homologous to the plant bifunctional terpene synthases PpCPS/KS (AB302933.1), JsCPS/KS (BAJ39816) and AgAs (AAK83563). Primers used for PCR and plasmid construction are shown in Supplementary Table S1. Based on the sequence information, HpDTC1 cDNA fragments were amplified by PCR using internal primers, HpDTC1-FR1 and then both 5′ and 3′ ends of HpDTC1 cDNA were amplified by RACE-PCR (Gene Racer, Invitrogen, Japan). The full length of HpDTC1 cDNA was amplified by PCR (Phusion High-Fidelity DNA Polymerase) using HpDTC1_cDNA primers. The genomic sequence of HpDTC1 was confirmed by PCR and the HpDTC1 promoter region was determined by four rounds of TAIL-PCR with arbitrary degenerate primers and gene specific primers. The 4.5 kbp HpDTC1 promoter region was amplified by PCR with PR-InF-HpDTC1 primers and subcloned into the KpnI and HindIII sites of pUC18 using the In-Fusion Cloning Kit (Clontech, Japan).
Recombinant protein expression and functional analysis
The open reading frame lacking the region encoding the plastid transit peptide of HpDTC1 was ligated into the KpnI and EcoRI sites of the pCold I vector (Takara Bio, Japan) using two sets of primers, HpDTC1-ORF. The recombinant HpDTC1 protein was produced in Escherichia coli strain M-15 as a His6-tagged protein according to the manufacture’s protocol and our previous work27. Enzyme products were identified using a GC-MS system (Bu-25, Jeol, Tokyo, Japan) equipped with a DB-WAX capillary column (0.25 mm ϕ, length 30 m and film thickness 0.25 μm; J&W Scientific).
Enzyme reactions and preparation of samples for analysis were performed as reported previously27. To identify an intermediate during the bifunctional cyclization reaction from GGDP to syn-pimara-7,15-diene, an amino acid-substituted mutant of HpDTC1 was prepared by PCR using two sets of primers (double-mutation-A and double-mutation-B in Supplementary Table S1) as described previously27,39. The products obtained from wild type HpDTC1 and the HpDTC1 mutant were determined by using a GC-MS system equipped with a DB-1 capillary column (0.25 mm ϕ, length 15 m and film thickness 0.25 μm; J&W Scientific).
RNA extraction and quantitative RT-PCR
Total RNA was extracted from H. plumaeforme gametophores treated with various elicitors at regular time intervals using a Sepasol-RNA I Super G (Nacalai Tesque, Japan) and subjected to cDNA synthesis using a PrimeScript RT reagent Kit with gDNA Eraser (Takara Bio, Japan). Quantitative RT-PCR (qRT-PCR) was performed using a Power SYBR Green PCR Master Mix (Applied Biosystems, CA, USA) for the HpDTC1 and HpACT3 genes on an ABI 7500 Fast Real-Time PCR System (Applied Biosystems) with standard mode. To calculate the transcript levels of characterized genes, the copy number of their mRNAs was determined by generating standard curves using a series of known concentrations of the target sequence. HpACT3 (DDBJ ID: LC129863, homologous to P. patens Actin3 gene (AAQ88110.1) found in RNA-seq) was used as an internal control to normalize the amount of mRNA. For each sample, the mean value from triplicate amplifications was used to calculate the transcript abundance. Sequences of PCR primers used for qRT-PCR analysis are provided in Supplementary Table S1.
Measurements of momilactone accumulation
Moss samples were soaked in 2 ml extraction solvent (MeOH/H2O, 80:20 [v/v]) and incubated at 4 °C overnight. Then, 5 μL of the extract was subjected to phytoalexin measurement by LC-MS/MS. An Agilent 1200 separation module (Agilent Technologies, Palo Alto, CA, USA) equipped with a CAPCELL CORE C18 column (50 mm long, 2.1 mm in diameter; Shiseido, Tokyo, Japan) was used for HPLC analysis. The mobile phase consisted of 0.05% AcOH in H2O (solvent A) and 0.05% AcOH in MeCN (solvent B). Elution was conducted using a linear gradient from 40% to 60% solvent B over 10 min at a flow rate of 0.2 mL/min and the eluate was monitored by an Agilent 6460 Triple Quadrupole mass spectrometer (Agilent Technologies, Palo Alto, CA, USA) using the positive ion mode. Electrospray conditions were as follows: capillary voltage, 3500 V; drying gas flow, 5 L/min nitrogen; drying gas temperature, 300 °C; nebulizer pressure, 45 psi; sheath gas temperature, 350 °C; and sheath gas flow, 11 L/min. Dwell time and fragmentor voltage were 200 ms and 135 V, respectively. The multiple reaction monitoring mode (MRM) was used in the MS/MS. MRM transitions were as follows: m/z 315.2/271.1 for momilactone A; m/z 331.2/269.1 for momilactone B.
Plasmid construction and moss transformation
A 3.4 kbp fragment of the HpDTC1 promoter (DDBJ ID: LC128407) was amplified by PrimeSTAR Max DNA polymerase (Takara Bio, Japan) using the primer pair, PR-DTC1-GUS (Supplementary Table S1). The pPIG1b-NGGII vector (accession number AB537478), the GUS reporter plasmid for P. patens, was digested with XbaI and BamHI. The 3.4 kbp fragment of the HpDTC1 promoter was inserted into the XbaI and BamHI sites of pPIG1b-NGGII using the In-Fusion Cloning Kit (Clontech, Japan) to construct the proHpDTC1::GUS vector. The proHpDTC1::GUS vector and original pPIG1b-NGGII vector were linearized by sse83871 (Takara Bio, Japan). Transformation of P. patens with the polyethylene glycol method was performed as described previously38. Blasticidin-resistant plants were cultured for an additional week without antibiotics and blasticidin resistance of the moss on the selection medium was confirmed. The appropriate integration of the proHpDTC1::GUS reporter and promoter-less pPIG1b-NGGII at the pPIG1b genome region was confirmed by PCR using appropriate primer sets and Southern gel-blot hybridization40.
Histochemical and Quantitative GUS Measurements
For GUS histochemical analysis, P. patens plants were transferred to a GUS-staining buffer (100 mM sodium phosphate, pH 7.0, 10 mM EDTA, 0.5 mM K4Fe(CN)6, 0.5 mM K3Fe(CN)6 and 0.1% Triton X-100) containing 1 mM 5-bromo-4-chloro-3-indolyl-β-d-glucuronide (X-Gluc) and then incubated at 37 °C until sufficient staining was achieved. For quantitative measurement for GUS activity, P. patens gametophores (n = 10–15) were homogenized in an extraction buffer as previously described41. After centrifugation, GUS activity was fluorophotometrically measured with 1 mM 4-methyl umbelliferyl-β-D-glucuronide as a fluorogenic substrate at 37 °C.
Additional Information
How to cite this article: Okada, K. et al. HpDTC1, a Stress-Inducible Bifunctional Diterpene Cyclase Involved in Momilactone Biosynthesis, Functions in Chemical Defence in the Moss Hypnum plumaeforme. Sci. Rep. 6, 25316; doi: 10.1038/srep25316 (2016).
References
Nozaki, H. et al. Momilactone A and B as allelochemicals from moss Hypnum plumaeforme: first occurrence in bryophytes. Biosci Biotechnol Biochem 71, 3127–3130 (2007).
Kato, T. et al. Momilactones, growth inhibitors from rice, Oryza sativa. Tetrahedron Lett. 3861–3864 (1973).
Cartwright, D. W., Langcake, P., Pryce, R. J., Leworthy, D. P. & Ride, J. P. Isolation and characterization of two phytoalexins from rice as momilactones A and B. Phytochemistry 20, 535–537 (1981).
Okada, A. et al. Elicitor induced activation of the methylerythritol phosphate pathway toward phytoalexins biosynthesis in rice. Plant Mol Biol 65, 177–187 (2007).
Hasegawa, M. et al. Phytoalexin Accumulation in the Interaction Between Rice and the Blast Fungus. Molecular Plant-Microbe Interactions 23, 1000–1011 (2010).
Kodama, O., Suzuki, T., Miyakawa, J. & Akatsuka, T. Ultraviolet-induced accumulation of phytoalexins in rice leaves. Agric. Biol. Chem. 52, 2469–2473 (1988).
Kodama, O., Yamada, A., Yamamoto, A., Takemoto, T. & Akatsuka, T. Induction of phytoalexins with heavy metal ions in rice leaves. J. Pestic. Sci. 13, 615–617 (1988).
Shimizu, T. et al. OsJAR1 Contributes Mainly to Biosynthesis of the Stress-Induced Jasmonoyl-Isoleucine Involved in Defense Responses in Rice. Bioscience Biotechnology and Biochemistry 77, 1556–1564 (2013).
De Leon, I. P. et al. Physcomitrella patens activates reinforcement of the cell wall, programmed cell death and accumulation of evolutionary conserved defence signals, such as salicylic acid and 12-oxo-phytodienoic acid, but not jasmonic acid, upon Botrytis cinerea infection. Molecular Plant Pathology 13, 960–974 (2012).
Kato-Noguchi, H. & Kobayashi, K. Jasmonic acid, protein phosphatase inhibitor, metals and UV-irradiation increased momilactone A and B concentrations in the moss Hypnum plumaeforme. J Plant Physiol 166, 1118–1122 (2009).
Otomo, K. et al. Diterpene cyclases responsible for the biosynthesis of phytoalexins, momilactones A, B and oryzalexins A-F in rice. Biosci Biotechnol Biochem 68, 2001–2006 (2004).
Shimura, K. et al. Identification of a biosynthetic gene cluster in rice for momilactones. J Biol Chem 282, 34013–34018 (2007).
Wang, Q., Hillwig, M. L. & Peters, R. J. CYP99A3: functional identification of a diterpene oxidase from the momilactone biosynthetic gene cluster in rice. Plant J 65, 87–95 (2011).
Yamamura, C. et al. Diterpenoid phytoalexin factor, a bHLH transcription factor, plays a central role in the biosynthesis of diterpenoid phytoalexins in rice. Plant J 84, 1100–1113 (2015).
Hayashi, K. et al. Identification and functional analysis of bifunctional ent-kaurene synthase from the moss Physcomitrella patens. FEBS Lett 580, 6175–6181 (2006).
Kawaide, H., Imai, R., Sassa, T. & Kamiya, Y. Ent-kaurene synthase from the fungus Phaeosphaeria sp. L487. cDNA isolation, characterization and bacterial expression of a bifunctional diterpene cyclase in fungal gibberellin biosynthesis. J Biol Chem 272, 21706–21712 (1997).
Vogel, B. S., Wildung, M. R., Vogel, G. & Croteau, R. Abietadiene synthase from grand fir (Abies grandis). cDNA isolation, characterization and bacterial expression of a bifunctional diterpene cyclase involved in resin acid biosynthesis. J Biol Chem 271, 23262–23268 (1996).
Kawaide, H., Sassa, T. & Kamiya, Y. Functional analysis of the two interacting cyclase domains in ent-kaurene synthase from the fungus Phaeosphaeria sp. L487 and a comparison with cyclases from higher plants. J Biol Chem 275, 2276–2280 (2000).
Wilderman, P. R., Xu, M., Jin, Y., Coates, R. M. & Peters, R. J. Identification of syn-pimara-7,15-diene synthase reveals functional clustering of terpene synthases involved in rice phytoalexin/allelochemical biosynthesis. Plant Physiol 135, 2098–2105 (2004).
Ponce de Leon, I., Hamberg, M. & Castresana, C. Oxylipins in moss development and defense. Front Plant Sci 6, 483 (2015).
Yamada, A., Shibuya, N., Kodama, O. & Akatsuka, T. Induction of Phytoalexin Formation in Suspension-Cultured Rice Cells by N-Acetyl-Chitooligosaccharides. Bioscience Biotechnology and Biochemistry 57, 405–409 (1993).
Cho, E. M. et al. Molecular cloning and characterization of a cDNA encoding ent-cassa-12,15-diene synthase, a putative diterpenoid phytoalexin biosynthetic enzyme, from suspension-cultured rice cells treated with a chitin elicitor. Plant J 37, 1–8 (2004).
Lehtonen, M. T. et al. Quickly-released peroxidase of moss in defense against fungal invaders. New Phytol 183, 432–443 (2009).
Ponce de Leon, I. et al. Erwinia carotovora elicitors and Botrytis cinerea activate defense responses in Physcomitrella patens. BMC Plant Biol 7, 52 (2007).
Oliver, J. P. et al. Pythium infection activates conserved plant defense responses in mosses. Planta 230, 569–579 (2009).
Yamaguchi, S., Sun, T., Kawaide, H. & Kamiya, Y. The GA2 locus of Arabidopsis thaliana encodes ent-kaurene synthase of gibberellin biosynthesis. Plant Physiol 116, 1271–1278 (1998).
Shimane, M. et al. Molecular evolution of the substrate specificity of ent-kaurene synthases to adapt to gibberellin biosynthesis in land plants. Biochem J 462, 539–546 (2014).
Sugai, Y. et al. Enzymatic (13)C labeling and multidimensional NMR analysis of miltiradiene synthesized by bifunctional diterpene cyclase in Selaginella moellendorffii. J Biol Chem 286, 42840–42847 (2011).
Hall, D. E. et al. Evolution of conifer diterpene synthases: diterpene resin acid biosynthesis in lodgepole pine and jack pine involves monofunctional and bifunctional diterpene synthases. Plant Physiol 161, 600–616 (2013).
Ponce, d. L. I. The Moss Physcomitrella patens as a Model System to Study Interactions between Plants and Phytopathogenic Fungi and Oomycetes. J Pathog 2011, 719873 (2011).
Stumpe, M. et al. The moss Physcomitrella patens contains cyclopentenones but no jasmonates: mutations in allene oxide cyclase lead to reduced fertility and altered sporophyte morphology. New Phytol 188, 740–749 (2010).
Okada, A. et al. OsTGAP1, a bZIP transcription factor, coordinately regulates the inductive production of diterpenoid phytoalexins in rice. J Biol Chem 284, 26510–26518 (2009).
Miyamoto, K. et al. Identification of target genes of the bZIP transcription factor OsTGAP1, whose overexpression causes elicitor-induced hyperaccumulation of diterpenoid phytoalexins in rice cells. PLoS One 9, e105823 (2014).
Chujo, T. et al. Overexpression of phosphomimic mutated OsWRKY53 leads to enhanced blast resistance in rice. PLoS One 9, e98737 (2014).
Akagi, A. et al. WRKY45-dependent priming of diterpenoid phytoalexin biosynthesis in rice and the role of cytokinin in triggering the reaction. Plant Mol Biol 86, 171–183 (2014).
Miyamoto, K. et al. Overexpression of the bZIP transcription factor OsbZIP79 suppresses the production of diterpenoid phytoalexin in rice cells. J Plant Physiol 173C, 19–27 (2014).
Yokotani, N. et al. WRKY76 is a rice transcriptional repressor playing opposite roles in blast disease resistance and cold stress tolerance. Journal of Experimental Botany 64, 5085–5097 (2013).
Nishiyama, T., Hiwatashi, Y., Sakakibara, I., Kato, M. & Hasebe, M. Tagged mutagenesis and gene-trap in the moss, Physcomitrella patens by shuttle mutagenesis. DNA Res 7, 9–17 (2000).
Kawaide, H. et al. Identification of the single amino acid involved in quenching the ent-kauranyl cation by a water molecule in ent-kaurene synthase of Physcomitrella patens. FEBS J 278, 123–133 (2011).
Ishikawa, M. et al. Physcomitrella Cyclin-Dependent Kinase A Links Cell Cycle Reactivation to Other Cellular Changes during Reprogramming of Leaf Cells. Plant Cell 23, 2924–2938 (2011).
Yamazoe, A., Hayashi, K., Kepinski, S., Leyser, O. & Nozaki, H. Characterization of terfestatin A, a new specific inhibitor for auxin signaling. Plant Physiol 139, 779–789 (2005).
Acknowledgements
This work was supported by JSPS Grant-in-Aid for Scientific Research (C) 25350972. We thank Prof. Naoki Nishimura (Okayama University of Science, Japan) for the collection and identification of Hypnum plumaeforme. We thank Prof. Mitsuyasu Hasebe (NIBB, Japan) for kindly providing the pPIG1b-NGG vector and for useful suggestions on moss transformation, Prof. Hideyuki Matsuura (Hokkaido University, Japan) for providing OPDA and Prof. Yoshiteru Noutoshi (Okayama University, Japan) for the helpful comments.
Author information
Authors and Affiliations
Contributions
K.O., H.K., H.Y., H. Nozaki and K.H. participated in the design of the study. K.O., K.M., S.M., R.K., H.K., K.F., Y.H., M. Nakajima, H. Nozaki, H. Nojiri, K.M., M. Natsume and K.H. performed the experiments and analyzed data. K.O., S.M. and H.K. interpreted the data and wrote the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Okada, K., Kawaide, H., Miyamoto, K. et al. HpDTC1, a Stress-Inducible Bifunctional Diterpene Cyclase Involved in Momilactone Biosynthesis, Functions in Chemical Defence in the Moss Hypnum plumaeforme. Sci Rep 6, 25316 (2016). https://doi.org/10.1038/srep25316
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep25316
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
