
Abstract
Stomatal conductance links plant water use and carbon uptake and is a critical process for the land surface component of climate models. However, stomatal conductance schemes commonly assume that all vegetation with the same photosynthetic pathway use identical plant water use strategies whereas observations indicate otherwise. Here, we implement a new stomatal scheme derived from optimal stomatal theory and constrained by a recent global synthesis of stomatal conductance measurements from 314 species, across 56 field sites. Using this new stomatal scheme, within a global climate model, subtantially increases the intensity of future heatwaves across Northern Eurasia. This indicates that our climate model has previously been under-predicting heatwave intensity. Our results have widespread implications for other climate models, many of which do not account for differences in stomatal water-use across different plant functional types and hence, are also likely under projecting heatwave intensity in the future.
Similar content being viewed by others
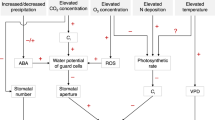


Introduction
Heatwaves are extreme phenomena that have major impacts on environmental, social, health and economic systems1. We define heatwaves as a series of three or more consecutive days during which daily maximum temperatures are higher than the calendar-day 90th percentile2. The frequency, intensity and duration of heatwaves are increasing in many parts of the globe3,4,5. Observations have highlighted an increase in the length of European heatwaves6 and the frequency of heatwave occurrence in China7 and Australia2,8. For example, the 2003 summer heatwave affected much of Western Europe and likely provided a precursor to future extremes across this region9. Many of these observed large-scale heatwaves have been linked to human activity via global warming10,11.
Future warming linked with increases in greenhouse gases is expected to increase the frequency, intensity and duration of heatwaves further5,12, particularly across the mid-latitudes including North America and Europe13,14. Heatwaves are associated with large-scale synoptic states15,16, which are influenced by modes of climate variability17. However, it is now well established from observational18 and modelling studies19,20 that heatwaves are also strongly modulated by the land surface if the synoptic scale weather generates persistent anticyclonic patterns and the planetary boundary-layer strongly couples the land to the atmosphere over consecutive days21. Under these circumstances, heatwaves intensify as desiccated soils and a surface radiation balance dominated by the exchange of sensible heat is coupled with the boundary-layer to lead to events such as the “mega-heatwaves” experienced in Europe during 2003 and 201019,21. Although the detailed role of the land surface on the exchange of water and energy during heatwaves remains uncertain22, there is evidence that capturing the detailed connection between the land and the atmosphere and how soil moisture impacts the surface energy balance to moderate or intensify heat waves, is necessary to produce realistic simulations of these phenomena19.
Within climate models, land surface models (LSMs) simulate soil moisture and partition available radiation at the surface between sensible and latent heat fluxes23. For vegetated surfaces, in particular over forests, the latent heat flux is principally controlled by stomata, as plants exchange water for carbon. Our ability to accurately simulate how soil moisture states and soil moisture variability affects heatwaves, therefore relies at least in part, on accurately modelling stomatal conductance (gs) under current and future CO2 concentrations. At the leaf scale, experiments commonly find that increasing CO2 results in increased photosynthesis24,25 and reduced water loss via lower gs26,27,28. However, there is increasing evidence29,30 that we cannot easily transfer our leaf/canopy level understanding of the response of transpiration due to CO2, to ecosystem scales. Nevertheless, any CO2-induced change in transpiration and/or soil “water-savings”, has the potential to alter future soil moisture state, soil moisture variability and transpiration, which may then feedback on the development of heatwaves over several days21. Given that heatwaves are associated with synoptic state and persistent anticyclonic conditions or so-called “blocking/persistent highs”31,32, these feedbacks are more likely to affect heatwave intensity than duration or frequency.
To date, the representation of gs in LSMs has been largely based on empirical models33,34,35. These models typically assume that differences in plant water use strategy are only tied to the photosynthetic pathway (C3 vs. C4). This assumption is not supported by experimental evidence; instead leaf level measurements suggest that plant water use strategies vary among species (or plant functional types, PFTs)36. Ignoring these differences among PFTs will likely result in errors in the simulated flux of moisture to the atmosphere. A recent collation of a global database of leaf-level gs measurements36 from 319 species across 56 field studies was used to parameterise differences in plant water use strategy among PFTs within the Community Atmosphere Biosphere Land Exchange (CABLE) model37. Parameters were estimated for each of the models PFTs by fitting Eq. 1 (see Methods) to this leaf-level dataset using a non-linear mixed effects model38.
This new gs model27,36 is similar in functional form to the previous empirical model35 used in CABLE and many other LSMs but is derived following optimal stomal theory. Consequently, model parameters carry biological meaning and can be hypothesised to vary with climate and plant water use strategy27. Such variations are supported by experimental data36. Offline CABLE simulations38 and coupled land-atmosphere simulations39 performed using the Australian Community Climate and Earth Systems Simulator (ACCESS1.3b)40, showed that this parameterisation led to a reduction in transpiration (up to 1 mm day−1) across boreal regions, which resulted in an increase in daily minimum and maximum temperatures (by up to 1 °C). These changes in contemporary simulations of water fluxes and daily warm temperature extremes were an improvement in the model’s climatology in comparison to observations during the boreal summer, especially over Eurasia39.
We extend our previous work38,39 to examine how an alterantive gs model, constrained by a global synthesis of leaf-level measurements, impacts upon future simulations of the likely incidence of heatwaves. We use the “business as usual” emission scenario (Representative Concentration Pathway 8.5 (RCP8.5))41 with the ACCESSv1.3 climate model. To the best of our knowledge, this is the first paper to implement a gs model within a global climate model focussing on future climate simulations, where the gs model parameters vary per PFT and are derived from best available data. We focus specifically on Eurasia for several reasons. Firstly, this is the region where the new gs scheme improved ACCESS’s climatology of evaporation and warm extremes39. Secondly, a previous evaluation of ACCESS’s simulations of extremes has shown large biases in extreme temperatures linked to clouds over North America42 and hence we avoid analysing this continent. Thirdly, Eurasia was shown to be sensitive to the parameterization of gs in earlier ACCESS experiments39 and finally, work by many researchers19,21,43 hints at this region being susceptible to large changes in warm extremes and heatwaves in the future.
Results
We first examine changes in warm extremes and surface moisture fluxes as illustrated in Fig. 1 showing the difference in mean Boreal summer (June-July-August) daily maximum temperature (TMAX, Fig. 1a), warmest yearly maximum temperature (TXx, Fig. 1b) and evapotranspiration (ET, Fig. 1c), averaged over 20 year intervals (2020–2099), between the new and the default gs scheme (i.e., Experiment minus Control). TMAX increases commonly by ∼1 °C but by more than 1.5 °C over Western Europe and 2 °C in some regions. The impact of the new gs scheme on TXx is larger, reaching 5 °C over widespread regions. Not surprisingly, there is a strong similarity between the patterns of temperature increases, decreases in ET (Fig. 1c) and subsequent decrease in precipitation (Fig. 1d), consistent with our previous work39. We note that the difference in both TMAX and TXx between models is largest during the period 2040–2059 and decreases towards the end of the century. One possible explanation for this decrease is that at high leaf temperatures (ca. 30 °C), photosynthesis and stomatal conductance (and thus transpiration) are reduced due to photosynthetic inhibition (Fig. S2). This response to high temperature minimises the differences in transpiration between the models that originally resulted from the more conservative water use paramaterisation in the new scheme.

Difference (Experiment minus Control) in mean Boreal summer (June-July-August) (a) daily maximum temperature (TMAX, top row), (b) warmest maximum temperature (TXx, middle row) and (c) evapotranspiration (ET, bottom row) and (d) precipitation (mm day−1), averaged over 20 year intervals between 2020–2099. Stippling shows regions where differences are statistically significant at the 95% level using the student’s t-test and the false discovery method for field significance. This figure was created using NCLV6.2.1 (http://www.ncl.ucar.edu/).
Furthermore, the two gs schemes have different sensitivities to vapor pressure deficit (VPD), with the default model showing stronger sensitivity at high VPD (>3 kPa)38. Thus, as dryland expansion accelerates under climate change44 and the air temperature and VPD increase towards the end of the 21st century, the difference in predicted transpiration between the two models becomes smaller (Fig. S2 and related text), which potentially accounts for the smaller effect on TMAX and TXx compared to earlier in the century. Nevertheless, there are still large differences between the models across most of Eurasia at the end of the century (2 to 4 °C for TXx).
The increases in TMAX and TXx and a decrease in ET can be clearly seen in the probability density functions (PDFs, Fig. 2). There is a clear shift to the right for the PDF of TMAX and TXx, but the limits of the lower and upper tails are mostly unchanged. The new gs scheme does not lead to the emergence of temperatures not previously experienced across the region; rather, it leads to a much more frequent occurrence of hot temperatures. Clearly, this change is linked to a shift in the PDF of ET to the left, such that ET exceeding 4 mm day−1 is rare with the new gs scheme, but common using the old scheme.

Probability distribution function (PDF, %) of monthly mean Boreal summer (June-July-August) daily maximum temperature (TMAX, left plot) warmest maximum temperature (TXx, middle plot) and evapotranspiration (ET, right plot) over the period 2020–2099. Results using the new gs are shown in blue (i.e., experiment) and the default gs scheme is in black (i.e., the control). This figure was created using NCLV6.2.1 (http://www.ncl.ucar.edu/).
We next examined the influence of the change in gs on heatwave duration, frequency and intensity (see Methods for definition). The changes in heatwave duration and frequency were very small, but changes in heatwave intensity (HWI) were large (Fig. 3). During the earlier part of the century (2020–2039), there are regions of both increases and reductions in HWI indicating that the forcing associated with the change in gs is commonly smaller than internal model variability. However, by 2040–2059, the new scheme results in an increase in HWI everywhere, with particularly large increases over western Europe, western Russia and eastern China, where HWI increases by 6–7 °C. Similarly to the changes in TMAX and TXx, the magnitude of the increase in HWI decreases towards the end of the century, but remains higher than 5 °C in many regions.

Same as in Fig. 1 except showing the change in heatwave intensity (HWI). This figure was created using NCLV6.2.1 (http://www.ncl.ucar.edu/).
Discussion and Conclusion
The increase in future (2020–2099) simulated TXx resulting from changing the representation of gs is approximately 4–5 °C over Western Europe. This sensitivity to gs can be put into context by recognising that this change is equivalent to more than half the increase projected under RCP8.541 (>1370 ppm CO2 equivalent in 2100) by an ensemble of climate models for 2081–210045. The change is similar to estimates reported for RCP4.541 (~650 ppm CO2 equivalent at stabilization after 2100) and higher than those reported for RCP2.641 (~490 equivalent before 2100 and declining) by 2081–210046. It is also similar in magnitude to the estimate reported for the change in heatwave intensity under RCP8.547. The increases in TXx due to the change in the gs model and parameterisation are therefore of the size reported for large increases in greenhouse gases.
Over western and northern Europe, the changes in TXx and heatwave intensity due to a change in the representation of gs as reported here are similar in terms of both pattern and intensity when compared to studies which have linked these changes to projected increases in greenhouse gases46,47. There are regions where the improved parameterization of gs led to increases in temperature and improved simulations39, particularly between around 45–60°N. The increases predominately occurred across regions defined as evergreen needleleaf forest, Tundra and crop PFTs.
The stomatal parameterisation we used in ACCESS accounts for differences in stomatal behaviour between PFTs and is supported by a global synthesis of leaf-level stomatal data36, in line with both predictions from optimal stomatal theory27,48 and the leaf and wood economic spectrum49,50. This empirical basis lends support to the robustness of these model simulations, which highlight the role of stomatal conductance in influencing future heatwaves. Nevertheless, some uncertainties remain. First, the data behind this parameterisation are measured at leaf scale; it has not been confirmed that the differences among PFTs observed at this scale also emerge at canopy/ecosystem scale. In light of our results, there is an urgent need for future work which tests how the stomatal parameterisation (g1, the sensitivity of the conductance to the assimilation rate, see material and methods) scales from the leaf to the canopy/ecosystem. Secondly, we have assumed all vegetation to have the same drought sensitivity. Observations suggest that vegetation adapted to different hydroclimates have different sensitivity51, which has significant consequences for ecosystem-scale water flux during drought periods52. A generic parameterisation for varying drought sensitivity across different vegetation types is another important priority.
We note a further significant caveat to our study: the ACCESS 1.3b climate model, in common with all climate models, has biases in its simulation of extremes42. The new gs parameterisation resolves some of these biases, at both site38 and global scales38,39. We also note that heatwaves are coupled phenomenon linking large-scale synoptic conditions, persistence, boundary layer coupling and land processes21. While ACCESS 1.3b is similar to other models in its representation of land-atmosphere coupling strength53 it remains a limitation to our study that we used a single climate model. We therefore encourage other groups to repeat our experiments to see if they can be generalized. Our results are also influenced by our use of a prescribed monthly climatology of leaf area index (LAI) derived from remote sensing estimates (see Methods). By prescribing the LAI, we are not allowing increases in leaf area due to CO2 to reduce any CO2 induced “water savings”. A model inter-comparison study54 which examined the response to elevated CO2 at two free-Air CO2 enrichment experiments found that even when LAI was not prescribed, the land surface component of ACCESS, i.e., CABLE, predicted modest changes in LAI (~5% increase). This result suggests that the use of prescribed LAI is unlikely to affect the results shown here for ACCESS, but clearly this may vary in other climate models. As both simulations prescribed the same LAI, the result is robust to assumptions of leaf area and CO2 and instead highlights the direct impact of the change in gs scheme and parameterization. Nevertheless, we plan to investigate the influence of prognostic LAI between the two schemes in future work.
The impact of the revised gs scheme on heatwave intensity is confronting, with increases of 5 °C (2040–2059). These increases are additive to those likely caused by increasing greenhouse gases over the same period47. The magnitude of these changes is large when compared to studies which have investigated the influence of soil moisture and vegetation dynamics on heatwaves. For example, lowering soil moisture by 25% for the 2003 European heatwaves is reported to lead to a maximum increase of 2 °C20 and other studies report changes of +0.5 °C by increasing LAI55 and ±1.5 °C due to dynamic phenology56. Our results are inevitably model-specific, but if confirmed by other groups, the current systematic under-estimation of future increases in heatwave intensity will have significant implications for socio-economic and environmental systems. We note that our revised parameterization of gs had no impact on the frequency or duration of heatwaves, since these are primarily driven by larger-scale synoptic-scale processes such as blocking highs57 and changing patterns of circulation58. However, our results do show that gs strongly affects the intensity of heatwaves over Eurasia and is therefore further evidence that land-atmosphere interactions are an important driver of extreme temperature events.
Methods
New representation of stomatal conductance
The default gs model35 used in ACCESSv1.3b40 has been described in detail in the literature37. The new gs scheme27 follows the form:

where A is the net assimilation rate (μmol m−2 s−1), Cs (μmol mol−1) and D (kPa) are the CO2 concentration and the vapour pressure deficit at the leaf surface, respectively and g0 (mol m−2 s−1) and g1 (kPa0.5) are fitted constants representing the residual stomatal conductance as A rate reaches zero and the slope of the sensitivity of gs to A, respectively. g0 is zero, leaving one key model parameter, g1, which theoretically represents the marginal carbon cost of water27.
The model was parameterised for the different PFTs (Fig. S1) using a global synthesis of stomatal measurements compiled from 314 species, across 56 field sites, covering the Arctic tundra, boreal regions, temperate forests and tropical rainforest biomes36. Values are shown in Table S1. The default gs scheme in CABLE has two fitted parameters which only vary by photosynthetic pathway (C3 versus C4) but not by PFT. More details on the differences between the default and new scheme and the implementation of the new scheme in CABLE can be found in our earlier work38,39.
Simulations
The ACCESS model setup is identical to our previous work in evaluating the new gs model under current climate39, except that simulations use sea surface temperatures from a previous fully-coupled simulation with ACCESS1.3 driven by the RCP8.5 emission scenario41 (official CMIP5 submission). Five ensembles were run; each initialised a year apart, with the default gs scheme (i.e., the control) and the new scheme (i.e., the experiment). All results shown are for the ensemble mean. We performed statistical significance testing of the differences between the experiment and the control using the student’s- t-test at 95% confidence interval and tested for field significance using the false discovery rate method59. Similar to our previous work39, nutrient-limited carbon pool dynamics and dynamic phenology were not activated, as the focus was on biophysical effects of the new gs scheme. Leaf area index (LAI) was prescribed as a monthly climatology derived from MODIS estimates. Results are also only shown between 30°W-150°E longitude and 30°N-80°N latitude, corresponding to the region where the new gs scheme improved ACCESS’s climatology of ET and warm extremes when compared to observations39.
Heatwave definition
Following the literature on heatwaves (HWs)2, we use thresholds based on percentiles rather than absolute values, with an event defined as temperatures exceeding the 90% percentile of daily maximum temperatures for at least 3 consecutive days. The percentiles are computed for each calendar day over a moving window over a user-defined base period, which is a commonly adopted approach5,43,46,60. To account for seasonality, we use a 15-day moving window over a 30-year base period during the first 30 years of the simulation (2020–2049) to provide the baseline. We note that most studies use 1961–199046,60; however, although we do have data over this period, these were generated using observed prescribed sea surface temperatures39. So, for consistency, we use the first 30 years of our simulation as baseline. The percentiles are computed for each of the 5 ensembles of the control and experiment separately. The HW-duration is the mean length (in days) of heatwave events during summer; the HW-frequency is the number of HW events; the HW-intensity is the mean temperature during the HW events; and the HW-max-intensity is the maximum temperature during the HW events. Indices are averaged across the 5 ensembles for the control and experiment and the ensemble mean difference is shown between the experiment and the control.
Additional Information
How to cite this article: Kala, J. et al. Impact of the representation of stomatal conductance on model projections of heatwave intensity. Sci. Rep. 6, 23418; doi: 10.1038/srep23418 (2016).
References
IPCC. Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation. A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change [ Field, C. B. et al. (eds)]. Cambridge University Press, Cambridge, UK and New York, NY, USA, 582 pp (2012).
Perkins, S. E. & Alexander, L. V. On the Measurement of Heat Waves. J Clim 26, 4500–4517 (2013).
Alexander, L. V. et al. Global observed changes in daily climate extremes of temperature and precipitation. J Geophys Res 111, D05109 (2006).
Seneviratne, S. I. et al. Changes in climate extremes and their impacts on the natural physical environment. In: Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation [ Field, C. B. et al. (eds)]. A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change (IPCC). Cambridge University Press, Cambridge, UK and New York, NY, USA, pp. 109–230 (2012).
Perkins, S. E., Alexander, L. V. & Nairn, J. R. Increasing frequency, intensity and duration of observed global heatwaves and warm spells. Geophys Res Lett 39, L20714 (2012).
Della-Marta, P. M., Haylock, M. R., Luterbacher, J. & Wanner, H. Doubled length of western European summer heat waves since 1880. J Geophys Res 112, D15103 (2007).
Ding, T., Qian, W. & Yan, Z. Changes in hot days and heat waves in China during 1961–2007. Int J Climatol 30, 1452–1462 (2010).
Alexander, L. V. & Arblaster, J. M. Assessing trends in observed and modelled climate extremes over Australia in relation to future projections. Int J Climatol 29, 417–435 (2009).
Schär, C. et al. The role of increasing temperature variability in European summer heatwaves. Nature 427, 332–336 (2004).
Stott, P. A., Stone, D. A. & Allen, M. R. Human contribution to the European heatwave of 2003. Nature 432, 610–614 (2004).
Lewis, S. C. & Karoly, D. J. Anthropogenic contributions to Australia’s record summer temperatures of 2013. Geophys Res Lett 40, 3705–3709 (2013).
Orlowsky, B. & Seneviratne, S. I. On the spatial representativeness of temporal dynamics at European weather stations. Int J Climatol 34, 3154–3160 (2014).
Lau, N.-C. & Nath, M. J. Model Simulation and Projection of European Heat Waves in Present-Day and Future Climates. J Clim 27, 3713–3730 (2014).
Andrade, C., Fraga, H. & Santos, J. A. Climate change multi-model projections for temperature extremes in Portugal: Multi-model ensemble projections for temperature in Portugal. Atmos Sci Lett 15, 149–156 (2014).
Pezza, A. B., Rensch, P. & Cai, W. Severe heat waves in Southern Australia: synoptic climatology and large scale connections. Clim Dyn 38, 209–224 (2011).
Parker, T. J., Berry, G. J., Reeder, M. J. & Nicholls, N. Modes of climate variability and heat waves in Victoria, southeastern Australia. Geophys Res Lett 41 (2014).
Perkins, S. E., Argüeso, D. & White, C. J. Relationships between climate variability, soil moisture and Australian heatwaves: Australian Climate and Heatwaves. J Geophys Res 120, 8144–8164 (2015).
Hirschi, M. et al. Observational evidence for soil-moisture impact on hot extremes in southeastern Europe. Nature Geosci 4, 17–21 (2010).
Seneviratne, S. I., Lüthi, D., Litschi, M. & Schär, C. Land-atmosphere coupling and climate change in Europe. Nature 443, 205–209 (2006).
Fischer, E. M., Seneviratne, S. I., Vidale, P. L., Lüthi, D. & Schär, C. Soil Moisture-Atmosphere Interactions during the 2003 European Summer Heat Wave. J Clim 20, 5081–5099 (2007).
Miralles, D. G., Teuling, A. J., van Heerwaarden, C. C. & Vilà-Guerau de Arellano, J. Mega-heatwave temperatures due to combined soil desiccation and atmospheric heat accumulation. Nature Geosci 7, 345–349 (2014).
Teuling, A. J. et al. Contrasting response of European forest and grassland energy exchange to heatwaves. Nature Geosci 3, 722–727 (2010).
Pitman, A. J. The evolution of and revolution in, land surface schemes designed for climate models. Int J Climatol 23, 479–510 (2003).
Kimball, B. A., Mauney, J. R., Nakayama, F. S. & Idso, S. W. Effects of increasing atmospheric CO2 on vegetation. Vegetatio 104/105, 65–75 (1993).
Mooney, H. A. et al. The terrestrial biosphere and global change: Ecosystem physiology responses to global change. In Implications of Global Change for Natural and Managed Ecosystems: A Synthesis of GCTE and Related Research. Eds Walker, B. H., Canadell, J. & Ingram, J. S. I. Cambridge University Press, Cambridge (1999).
Morison, J. I. L. Sensitivity of stomata and water use efficiency to high CO2 . Plant, Cell Environ 8, 467–474 (1985).
Medlyn, B. E. et al. Reconciling the optimal and empirical approaches to modelling stomatal conductance. Global Change Biol 17, 2134–2144 (2011).
Ainsworth, E. A. & Rogers, A. The response of photosynthesis and stomatal conductance to rising CO2: Mechanisms and environmental interactions. Plant, Cell Environ 30, 258–270 (2007).
Wullschleger, S. D., Tschaplinski, T. J. & Norby, R. J. Plant water relations at elevated CO2 - Implications for water-limited environments. Plant, Cell Environ 25, 319–331 (2002).
De Kauwe, M. G. et al. Forest water use and water use efficiency at elevated CO2: A model-data intercomparison at two contrasting temperate forest FACE sites. Global Change Biol 19, 1759–1779 (2013).
Marshall, A. G. et al. Intra-seasonal drivers of extreme heat over Australia in observations and POAMA-2. Clim Dyn 43, 1915–1937 (2014).
Perkins, S. E. A review on the scientific understanding of heatwaves? Their measurement, driving mechanisms and changes at the global scale. Atmos Res 164–165, 242–267 (2015).
Jarvis, P. G. The Interpretation of the Variations in Leaf Water Potential and Stomatal Conductance Found in Canopies in the Field. Philos Trans R Soc 273, 593–610 (1976).
Ball, J. T., Woodrow, I. E. & Berry, J. A. A model predicting stomatal conductance and its contribution to the control of photosynthesis. In: Progress in photosynthesis research, proceedings of the VIIth International Congress on Photosynthesis, 221–224 (1987).
Leuning, R. A critical appraisal of a combined stomatal-photosynthesis model for C3 plants. Plant, Cell Environ 18, 339–355 (1995).
Lin, Y.-S. et al. Optimal stomatal behaviour around the world. Nature Clim Change 5, 459–464 (2015).
Wang, Y. P. et al. Diagnosing errors in a land surface model (CABLE) in the time and frequency domains. J Geophys Res 116, G01034 (2011).
De Kauwe, M. G. et al. A test of an optimal stomatal conductance scheme within the CABLE land surface model. Geosci Mod Dev 8, 431–452 (2015).
Kala, J. et al. Implementation of an optimal stomatal conductance scheme in the Australian Community Climate Earth Systems Simulator (ACCESS1.3b). Geosci Model Dev 8, 3877–3889 (2015).
Bi, D. et al. The ACCESS coupled model: Description, control climate and evaluation. Aust Meteorol Ocean Soc J 63, 9–32 (2013).
Moss, R. H. et al. The next generation of scenarios for climate change research and assessment. Nature 463, 747–756 (2010).
Lorenz, R. et al. Representation of climate extreme indices in the ACCESS1.3b coupled atmosphere-land surface model. Geosci Mod Dev 7, 545–567 (2014).
Fischer, E. M., Rajczak, J. & Schär, C. Changes in European summer temperature variability revisited. Geophys Res Lett 39, L19702 (2012).
Huang, J., Yu, H., Guan, X., Wang, G. & Guo, R. Accelerated dryland expansion under climate change. Nature Clim Change 6, 166–171 (2016).
Collins, M. et al. Long-term Climate Change: Projections, Commitments and Irreversibility. In: Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [ Stocker, T. F. et al. (eds)]. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA (2013).
Sillmann, J., Kharin, V. V., Zwiers, F. W., Zhang, X. & Bronaugh, D. Climate extremes indices in the CMIP5 multimodel ensemble: Part 2. Future climate projections. J Geophys Res 118, 2473–2493 (2013).
Schoetter, R., Cattiaux, J. & Douville, H. Changes of western European heat wave characteristics projected by the CMIP5 ensemble. Clim Dyn 45, 1601–1616 (2015).
Cowan, I. R. & Farquhar, G. D. Stomatal function in relation to leaf metabolism and environment. Symp Soc Exp Biol 31, 471–505 (1977).
Wright, I. J., Falster, D. S., Pickup, M. & Westoby, M. Cross-species patterns in the coordination between leaf and stem traits and their implications for plant hydraulics. Physiol Plant 127, 445–456 (2006).
Chave, J., Coomes, D., Jansen, S., Lewis, S. L., Swenson, N. G. & Zanne, A. E. Towards a worldwide wood economics spectrum. Ecol Lett 12, 351–366 (2009).
Zhou, S., Medlyn, B., Sabaté, S., Sperlich, D. & Prentice, I. C. Short-term water stress impacts on stomatal, mesophyll and biochemical limitations to photosynthesis differ consistently among tree species from contrasting climates. Tree Physiol 10, 1035–1046 (2014).
De Kauwe, M. G. et al. Do land surface models need to include differential plant species responses to drought? Examining model predictions across a mesic-xeric gradient in Europe. Biogeosci 12, 7503–7518 (2015).
Lorenz, R., Pitman, A. J., Hirsch, A. L. & Srbinovsky, J. Intraseasonal versus Interannual Measures of Land-Atmosphere Coupling Strength in a Global Climate Model: GLACE-1 versus GLACE-CMIP5 Experiments in ACCESS1.3b. J Hydrometeorol 16, 2276–2295 (2015).
De Kauwe, M. G. et al. Where does the carbon go? A model–data intercomparison of vegetation carbon allocation and turnover processes at two temperate forest free-air CO2 enrichment sites. New Phytol 203, 883–899 (2014).
Lorenz, R., Davin, E. L., Lawrence, D. M., Stöckli, R. & Seneviratne, S. I. How Important is Vegetation Phenology for European Climate and Heat Waves? J Clim 26, 10077–10100 (2013).
Stéfanon, M., Drobinski, P., D’Andrea, F. & de Noblet-Ducoudré, N. Effects of interactive vegetation phenology on the 2003 summer heat waves. J Geophys Res 117, D24103 (2012).
He, Y., Huang, J. & Ji, M. Impact of land–sea thermal contrast on interdecadal variation in circulation and blocking. Clim Dyn 43, 3267–3279 (2014).
Purich, A. et al. Atmospheric and Oceanic Conditions Associated with Southern Australian Heat Waves: A CMIP5 Analysis. J Clim 27, 7807–7829 (2014).
Wilks, D. S. On “Field Significance” and the False Discovery Rate. Journal of Applied Meteorology and Climatology 45, 1181–1189 (2006).
Sillmann, J., Kharin, V. V., Zhang, X., Zwiers, F. W. & Bronaugh, D. Climate extremes indices in the CMIP5 multimodel ensemble: Part 1. Model evaluation in the present climate. J Geophys Res 118, 1716–1733 (2013).
Acknowledgements
This work was supported by the Australian Research Council Centre of Excellence for Climate System Science (CE110001028). S. E. P.-K. is supported by an Australian Research Council Discovery Early Career Researcher Award (DE140100952). We thank the National Computational Infrastructure at the Australian National University, an initiative of the Australian Government, for access to supercomputer resources; the Commonwealth Scientific and Industrial Research Organisation and the Bureau of Meteorology for their support in the use of the CABLE and ACCESS models; and, David Fuchs for assisting with Python scripting. All this assistance is gratefully acknowleged.
Author information
Authors and Affiliations
Contributions
A.J.P. conceptualized the project. J.K. carried out all the analysis. M.D.K. and J.K. implemented the new stomatal conductance scheme in CABLE. J.K. and R.L. ran all ACCESS simulations. B.E.M. and Y.-P.W. developed the new and default stomatal conductance schemes respectively. S.E.P.-K. assisted in implementing the heatwave indices. J.K., M.D.K. and A.J.P. lead the writing and all other co-authors made substantial contributions towards several draft manuscripts.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Kala, J., De Kauwe, M., Pitman, A. et al. Impact of the representation of stomatal conductance on model projections of heatwave intensity. Sci Rep 6, 23418 (2016). https://doi.org/10.1038/srep23418
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep23418
This article is cited by
-
Radiative forcing geoengineering causes higher risk of wildfires and permafrost thawing over the Arctic regions
Communications Earth & Environment (2024)
-
Stomatal responses of terrestrial plants to global change
Nature Communications (2023)
-
Diminishing CO2-driven gains in water-use efficiency of global forests
Nature Climate Change (2020)
-
The intensification of Arctic warming as a result of CO2 physiological forcing
Nature Communications (2020)
-
Amplification of heat extremes by plant CO2 physiological forcing
Nature Communications (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
