
Abstract
The LC8 family of small ~8 kD proteins are highly conserved and interact with multiple protein partners in eukaryotic cells. LC8-binding modulates target protein activity, often through induced dimerization via LC8:LC8 homodimers. Although many LC8-interactors have roles in signaling cascades, LC8’s role in developing epithelia is poorly understood. Using the Drosophila wing as a developmental model, we find that the LC8 family member Cut up (Ctp) is primarily required to promote epithelial growth, which correlates with effects on the pro-growth factor dMyc and two genes, diap1 and bantam, that are classic targets of the Hippo pathway coactivator Yorkie. Genetic tests confirm that Ctp supports Yorkie-driven tissue overgrowth and indicate that Ctp acts through Yorkie to control bantam (ban) and diap1 transcription. Quite unexpectedly however, Ctp loss has inverse effects on ban and diap1: it elevates ban expression but reduces diap1 expression. In both cases these transcriptional changes map to small segments of these promoters that recruit Yorkie. Although LC8 complexes with Yap1, a Yorkie homolog, in human cells, an orthologous interaction was not detected in Drosophila cells. Collectively these findings reveal that that Drosophila Ctp is a required regulator of Yorkie-target genes in vivo and suggest that Ctp may interact with a Hippo pathway protein(s) to exert inverse transcriptional effects on Yorkie-target genes.
Similar content being viewed by others


Introduction
The LC8 family of cytoplasmic dynein light-chains, which includes vertebrate LC8 (aka DYNLL1/DYNLL2) and Drosophila Cut-up (Ctp), are small highly conserved proteins that are ubiquitously expressed and essential for viability1,2,3,4. The LC8 protein is 8 kilodaltons (kD) in size and was first identified as an accessory subunit in the dynein motor complex, within which an LC8 homodimer binds to and stabilizes a pair of dynein intermediate chains (DIC)1,5,6. However, the LC8 protein has since emerged as a general interaction hub with multiple dynein/motor-independent roles and binding partners3,7,8. In fact the majority of LC8 protein in mammalian cells is not associated with either dynein or microtubules1 and LC8 orthologs are encoded in the genomes of flowering plants that otherwise lack genes encoding heavy-chain dynein motors9.
Accumulating evidence has reinforced the idea that the primary role of LC8 in mammalian cells is to facilitate dimerization of its binding partners via LC8 self-association, a mechanism that has been termed ‘molecular velcro’7. LC8 can be found in association with over 40 proteins that function in diverse cellular processes, including intracellular transport, nuclear translocation, cell cycle progression, apoptosis, autophagy and gene expression8,10. LC8 is found in both the nucleus and cytoplasm and can interact with partners in either compartment. For example the mammalian kinase Pak1 binds and phosphorylates LC8 in the cytoplasm, which in turn enhances the ability of LC8 to interact with the BH3-only protein Bim and inhibit its pro-apoptotic activity11,12. Accordingly, overexpression of LC8 or the phosphorylation of LC8 by Pak1 enhances survival and proliferation of breast cancer cells in culture12. LC8 also binds estrogen receptor-α (ERα) and facilitates ERα nuclear translocation, which in turn recruits LC8 to the chromatin of ERα-target genes13,14,15. In the cytoplasm, LC8 is also found in association with the kidney and brain expressed protein (KIBRA), which is an upstream regulator of the Hippo tumor suppressor pathway16. KIBRA binding potentiates the effect of LC8 on nuclear translocation of ERα, suggesting crosstalk may occur between LC8-regulated pathways15. The KIBRA-LC8 complex also interacts with Sorting Nexin-4 (Snx4) to promote dynein-driven traffic of cargo between the early and recycling endosomal compartments17. Thus, LC8 has been linked to a variety of proteins in both the cytoplasm and nucleus that play important roles in signaling, membrane dynamics and gene expression.
Drosophila Ctp differs from vertebrate LC8/DYNLL by only four conservative amino acid substitutions across its 89 amino acid length. Similar to mammalian LC8, phenotypes produced by Ctp loss in flies imply roles in multiple developmental mechanisms. Drosophila completely lacking Ctp die during embryogenesis due to excessive and widespread apoptosis2,18. Partial loss of Ctp function causes thinned wing bristles and morphogenetic defects in wing development, as well as ovarian disorganization and female sterility2. Within salivary gland cells, Ctp promotes autophagy during pupation19, while in neuronal stem cells it localizes to centrosomes and influences mitotic spindle orientation and the symmetry of cell division20. Testes mutant for ctp have motor-dependent defects in spermatagonial divisions as well as motor-independent defects in cyst cell differentiation21. A recent study linked ctp mRNA expression to the zinc-finger transcription factor dASCIZ and showed that knockdown of either Ctp or dASCIZ reduces wing size22. In sum, this diversity of effects produced by Ctp loss in different Drosophila cell types suggest that Ctp plays important yet context specific roles in vivo. However our knowledge of molecular pathways that require Ctp and in turn underlie these developmental phenotypes associated with Ctp loss, remain poorly characterized.
Here we use a genomic null allele of ctp and a validated ctp RNAi transgene to assess the role of the Ctp/LC8/DYNLL protein family in pathways that act within the developing Drosophila wing epithelium. We find that clones of ctp null cells are quite small relative to controls and that RNAi depletion of Ctp shrinks the size of the corresponding segment of the adult wing without clear defects in mitotic progression or tissue patterning. The effect of Ctp depletion on adult wing size is primarily associated with a reduction in cell size, rather than cell division or cell number, implying a role for Ctp in supporting mechanisms that enable developmental growth. In assessing the effect of Ctp loss on multiple pathways that control wing growth, we detect robust effects on one–the Hippo pathway. The Hippo pathway is a conserved growth suppressor pathway that acts via its core kinase Warts to inhibit nuclear translocation of the coactivator Yorkie (Yki), which otherwise enters the nucleus, complexes with the DNA-binding factor Scalloped (Sd) and activates transcription of growth and survival genes23,24,25,26. In parallel to the effect of Ctp loss on clone and wing size, Ctp loss alters expression of the classic Yki target genes bantam and thread(th)/diap-1 in wing pouch cells. Parallel genetic tests confirm a requirement for ctp in Yki-driven tissue growth in the wing or eye. Quite unexpectedly however, Ctp loss has opposing effects on bantam and diap1 transcription in wing pouch cells: bantam transcription is strongly elevated while diap1 expression is strongly decreased in cells lacking Ctp. In each case, these effects map to small segments of DNA in the ban and diap1 promoters that recruit Yki transcriptional complexes24,25,27. Epistasis experiments confirm that Yki is required to activate the bantam promoter in Ctp-depleted cells and that transgenic expression of Yki can overcome the block to diap1 transcription. In sum these data argue that Ctp supports physiologic Hippo signaling in wing disc epithelial cells and that Ctp likely interacts with an as yet unidentified Hippo pathway protein(s) to exert inverse transcriptional effects on Yorkie-target genes. These types of inverse effects have not previously been described within the Hippo pathway and imply that distinct subsets of genes within the Yorkie transcriptome can be simultaneously activated and repressed in developing tissues via a mechanism that involves Ctp.
Materials and Methods
Fly Strains
All crosses were maintained at 25 °C unless otherwise noted. Alleles used in these studies (Bloomington stock number indicated) are as follows: thread-lacZ (#12093), ex697 (ex-lacZ, #44248), e2f1rM729 (e2f1-lacZ, #34054), UAS-yki-V5 (#28819), ykiB5 (#36290), mirrDE-Gal4 (#29650), EcRE-lacZ (#4517), Stat92E-10XGFP (#26198) (courtesy of R.Read), UAS-diap1 (#6657), UAS-p35 (#5073), UAS-dcr2;enGal4,UAS-GFP (#25752) obtained from the Bloomington Drosophila Stock Center. UAS-yki-IR (v104523) and UAS-ctp-IR (v43116) were obtained from the Vienna Drosophila Resource Center (VDRC). Other alleles used were ctpex3 (Gift of Bill Chia), enGal4/CyO, HRE-lacZ, HREmut6-lacZ, 2B2-lacZ, 2B2C-lacZ (all gift of D.J. Pan), brC12-lacZ (K. Irvine), GMR-Gal4, UAS-ykiS168A:GFP (K. Harvey), E-spl(m)ß-CD2, Su(H)-lacZ, PCNA-GFP, ban-3xGFP and UAS-yki (D.J. Pan).
Immunofluorescence Microscopy
Immunostaining and confocal microscopy performed as described previously on a Zeiss 510 inverted confocal microscope28. Primary antibodies include mouse anti-β-Gal 1:1000 (Promega); rabbit anti-LC8 (1:100) (W. Sale), mouse anti-Ci (1:50), mouse-α-Notch 1:10, mouse anti-Wg (1:800), mouse anti-Cut (1:100) and mouse anti-DIAP1 (1:50) (DSHB); rabbit α-yki 1:1000 (K. Irvine); mouse α-rat CD2 1:100 (Research Diagnostics, Inc.); mouse anti-V5 (1:200, Invitrogen); mouse anti-dpERK (1:10,000, Sigma); rabbit anti-phospho histone H3 (1:100), rabbit anti-phospho-Smad1/5 (1:100), rabbit anti-DCP1 (1:150), rabbit anti-Caspase3 (1:100) (Cell Signaling); mouse anti-dMyc (monoclonal P4C4-b10); mouse anti-Cyclin E (H. Richardson). BrdU assays performed as described previously (Robinson et al. 2010) with mouse anti-BrdU (1:100; Becton Dickinson).
Wing/Eye Measurements
Eyes/wings were imaged on a Leica DFC500 CCD camera and quantified with Image J. Posterior compartment ratio (PCR) = posterior compartment size/total wing size.
Results
Ctp is required for imaginal-disc derived adult tissues to grow to normal size
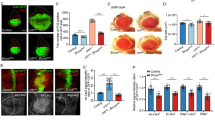
To test the role of ctp in the developing wing disc, a UAS transgene encoding a ctp RNA interference (RNAi) cassette (Vienna line #43116) was expressed in the posterior compartment of the larval wing disc at 25 °C using the engrailed-Gal4 driver (enGal4). Depletion of Ctp protein in posterior cells of en > ctp-IR discs was confirmed by immunostaining with an antibody raised against the Chlamydomonas reinhardtii homolog of Ctp that also cross-reacts with metazoan LC8/Ctp proteins1 (Figure S1). Ctp-depleted larval wing discs develop into adult wings with a fully penetrant phenotype of a shrunken posterior blade (Fig. 1A–C). This effect occurs without obvious scarring of the adult wing or major disruption in its pattern of venation, with the exception of occasional truncated cross veins (e.g. wing in Fig. 1B). Ctp-depletion with a second larval driver, mirrDE-Gal4, which is active in the dorsal half of developing larval discs29, shrinks the dorsal half of the adult eye (more darkly pigmented region above the dotted lines in Fig. 1D,E), contracts the dorsal surface of the adult head and leads to small, thin thoracic bristles (Figure S2). The Ctp-RNAi small-bristle phenotype matches a thin-bristle phenotype observed in adult flies carrying the hypomorphic ctp mutation, ctpins1 and further confirms the validity of the RNAi cassette2. As in the wing, Ctp depletion did not obviously scar the dorsal domain of the adult eye or strongly disrupt its surface organization, suggesting that Ctp depletion impairs growth of these organs but less so the mechanisms that drive their morphological patterning.

ctp loss reduces compartment size and clonal growth.
Paired images of control (A,D) and Ctp-depleted (B,E) adult wings and eyes generated using the en-Gal4 (A,B) or mirrDE-Gal4 (AKA dorsal eye (DE)-Gal4) (D,E) drivers in combination with the ctpIR transgene. en is expressed in posterior wing cells (below dotted line in (A,B)). DE is expressed in dorsal eye cells (above dotted line in (D,E)). (C) Box-plot quantitation of the wing posterior compartment ratio (PCR = P area/P + A areas) of the genotypes in (A) (n = 22) and (B) (n = 31). Standard error of the mean (SEM) is indicated. *p < 0.0001. (F) Confocal image of a ctp mosaic L3 wing disc (ctpex3,FRT19A/ubi-GFP,FRT19A,hsFlp) containing GFP-negative ctpex3 clones (arrows) and their paired GFP-positive twinspots. (G,H) Histogram plots of the two-dimensional area of multiple ctpex3 clone:twinspot pairs in the hinge/notum (G) (n = 29) or pouch (H) (n = 47) of L3 wing discs. Each pair of bars on the X-axis represents a distinct clone:twinspot pair. The clones are significantly smaller than their paired twinspots, *p = 0.0002 for (G), *p < 0.0001 for (H). (I) Bar graph plot of average ratios of ctpex3 clone:twinspot sizes in the hinge/notum versus the pouch. SEMs are indicated. *p = 0.0034.
To better assess the effects of Ctp loss on cell proliferation and survival, ctp mutant clones (GFP-) and their wildtype twinspots (GFP+) were generated by combining a heatshock(hs)-induced Flp mitotic recombination system with the ctpex3 null allele2. Within dissected larval wing discs, GFP-deficient ctpex3 clones could be found in the pouch, hinge and notum at 48 hrs post-clone induction (Fig. 1F). In the hinge and notum, these ctp null clones can reach a fairly large sizes (see arrows in Fig. 1F) but are nonetheless ~2-fold smaller than their control twinspots (Fig. 1F,G,I). This trend toward small clones is more pronounced in the pouch where ctp-null clones are occasionally missing and consistently quite small relative to their age-matched twinspots (e.g. see Fig. 1F). A quantitative measurement across multiple pouch clone pairs with extant twinspots shows an approximate 5-fold reduction in ctp mutant clone area (Fig. 1H,I). This apparently unequal growth deficit elicited by ctp loss in hinge/notum cells vs. pouch cells could nonetheless stem from perturbation of an otherwise ubiquitous cellular process, such as the mitotic spindle defects observed in ctp mutant neuronal stem cells20. Alternatively, it could indicate a role for Ctp in a molecular pathway or process that is especially important in pouch cells, similar to the cell-type specific and motor-independent requirement for ctp in germline and somatic cyst cells21.
Ctp loss elevates apoptosis and division of wing disc cells and reduces their size
To better assess the effects ctp loss on division and survival of larval wing disc cells, larval wing cells depleted of Ctp were probed with antibodies to cleaved caspase-3 (cC3), the incorporated nucleotide base analog bromodeoxyuridine (BrdU) and the mitotic marker phospho-histone H3 (pH3) (Fig. 2A–C). cC3 is moderately elevated in the posterior compartment of en > ctp-IR wing discs (Fig. 2A,A′) and an antibody to the cleaved caspase Dcp-1 detects elevated apoptosis in ctpex3 mutant wing pouch cells (Figure S3). This finding in disc cells parallels the widespread apoptosis that occurs in ctp mutant embryos2 and raises the possibility that death of ctp mutant cells may deprive the developing wing of a sufficient pool of cells to achieve normal size. To test this hypothesis, two transgenes encoding either the Drosophila protein Diap1, which blocks caspase cleavage, or the baculoviral protein p35, which inhibits active cleaved caspases30, were co-expressed with ctp-IR from the en driver (Fig. 2D). Neither of these anti-apoptotic transgenes were able to significantly rescue small en > ctp-IR wings, indicating that cell death is unlikely to be the sole cause of wing blade undergrowth induced by chronic Ctp-depletion.

Effects of Ctp loss on the division, survival and size of wing cells.
Confocal images of en > ctpIR L3 wing discs stained for (A′) cleaved Caspase-3 (cC3), (B′) the base analog BrdU, or (C′) phospho-histone H3 (PH3). Anti-Cubitis interruptus (Ci) staining marks the anterior domain (A–C). (D) Quantitative analysis of PCR in the indicated genotypes. en > diap1 (n = 13) is significantly larger than en > diap1 + ctpIR (n = 12) (*p < 0.0001) and en > p35 (n = 10) is significantly larger than en > p35 + ctpIR (n = 17) (*p < 0.0001). (E–E″) Box-plot quantitation and images of wing hair cell density in a fixed area in the posterior wing between the L4 and L5 wing veins in control (en > + ) (n = 10) or Ctp-depleted (en > ctpIR) (n = 9) adult wings. SEMs are indicated. *p < 0.0001. (F) Comparative effects of Ctp-depletion (en > + vs. en > ctpIR) on cell size (data in 2E) and PCR (data in 1C). en > + values are standardized to 100%.
The early mitotic marker phospho-histone H3 (pH3) is increased among Ctp-depleted larval wing pouch cells (Fig. 2C,C′). Taken in isolation, this higher abundance of pH3-positive ctp-IR larval wing cells could be indicative of an accelerated rate of mitotic entry or a slower rate of M-phase transit. Although disrupting LC8 function in cultured mammalian cells does not impede mitotic progression31, a past study in Drosophila attributed an excess-pH3 phenotype in Ctp-depleted cells to dynein-motor defects that block cells in mitosis and thus enrich for M-phase markers22. This mitotic-block model logically predicts that Ctp-depleted cells accumulate in M-phase and thus depopulate other phases of the cell cycle. However, BrdU-incorporation analysis of ctp-IR wing pouch cells does not show a depletion of S-phase cells and in fact seems to show an increase in the frequency of S-phase entry (Fig. 2B,B′). A very similar increase in cell division occurs in Drosophila germ cells lacking ctp21. Intriguingly ctp loss reduces cell division among neural stem cells32, suggesting that Ctp plays distinct proliferative roles in different cell types. In combination, the pH3 and BrdU data presented here argue that Ctp-depleted larval wing disc cells are able to actively transit between the mitotic and DNA synthesis phases of the cell cycle, suggesting that a cell cycle block is not the prime cause of small ctp-IR adult wings.
Genetic manipulations that reduce cell size can also shrink Drosophila adult organs. Thus, the lack of experimental evidence pointing to excess apoptosis or a proliferative deficit among Ctp-depleted cells prompted analysis of the effect of Ctp loss on cell size. Each cell in the adult wing generates a single hair, enabling a quantitative determination of cell density derived from hair counts within a fixed region of the adult wing. Applying this approach to an area of the posterior wing blade between the L4 and L5 veins reveals that en > ctp-IR wings have significantly higher hair density relative to control en > + wings, indicating that Ctp-depleted cells are smaller (Fig. 2E). The measured effect of en > ctp-IR depletion on cell size (~14% smaller; Fig. 2E) is of similar magnitude to its effect on the relative size of the posterior wing blade (~17% smaller; Fig. 1C), indicating that effects on cell size, rather than cell number, are likely be a central cause of small en > ctp-IR wings (Fig. 2F).
The smaller cell size of Ctp-depleted wing cells combined with the shorter, thinner thoracic bristles seen with mirrDE-Gal4 driven Ctp (Figure S2) is reminiscent of phenotypes induced by mutations in genes that support metabolic process of growth, such as the diminutive gene or the Minute ribosomal RNA genes33 (and reviewed in ref. 34). The diminutive (dm) locus encodes the Drosophila Myc protein (dMyc), a well-established pro-growth transcription factor that promotes cell and tissue growth35. Notably, ctpex3 null clones generated in the eye disc show reduced levels of dMyc protein as detected by immunofluorescence (Fig. 3A,A′). Within the wing disc, dMyc protein is normally detected throughout the dorsal and ventral halves of the pouch, with a region of cells along the dorsoventral boundary that express less dMyc36. In en > ctp-IR discs, dMyc staining is intact in the corresponding anterior regions but decreased in these areas that express the ctp-IR transgene (Figure S4). Consistent with this link between Ctp and dMyc protein levels, heterozygosity for the null allele dMycPL35 further shrinks en > ctp-IR wings (Fig. 3B). In sum these data suggest that reduced cell size contributes significantly to the undergrowth of en > ctp-IR adult wings and potentially link ctp to one or more of the transcriptional, translational and post-translational mechanisms that control dMyc levels in disc cells28,37,38.

Effect of Ctp loss on a panel of disc cell proliferation and/or growth pathways.
(A) Confocal image of an L3 eye disc (posterior to the right) mosaic for ctpex3 clones marked by the absence of β-galactosidase (βgal) (A) and stained for dMyc (A′). Note the drop in anti-dMyc fluorescence in ctpex3 clones in (A′). (B) Quantitation of PCR in the indicated adult genotypes shows that heterozygosity for the dMycPL35 allele (n = 6) has a dominant enhancing effect on the en > ctpIR phenotype (n = 31). SEMs are indicated. *p = 0.0037. (C–K, M) Confocal images of L3 wing discs with Ctp depleted in the posterior domain (right-side in all images, with Ci marking the anterior domain (C–E,H,J,K) or transgenic UAS-GFP marking the posterior domain (F,G,I,M)) and analyzed for each of the indicated factors: (C′) anti-Notch (N) to detect the C-terminal fragment of the Notch receptor; (D′) the E2F-activity reporter PCNA-GFP; (E′) the Notch-activity reporter E(spl)mβ-CD2; (F′) the E2f1-lacZ enhancer trap; (G′) the Notch-activity reporter Su(H)-lacZ; (H′) the Jak-Stat activity reporter Stat10x-GFP; (I′) anti-Wingless (Wg) to detect the ligand of the Wg/Wnt pathway; (J′) anti-phospho-Mothers against decapentaplegic (Mad) to detect signaling through the Dpp pathway; (K′) anti-Cut, a target of both Wg and Notch; (M′) the ecdysone receptor (EcR) reporter activity EcRE-lacZ. (L,N) L3 eye discs mosaic for ctpex3 clones marked by the absence of GFP (L,N): (L′) anti-diphospho-Erk to detect signaling downstream of the Drosophila epidermal growth factor receptor (DER); (N′) anti-CyclinE (CycE), which is rate-limiting for progression into S-phase.
Ctp is dispensable for multiple signaling pathways but genetically interacts with Yorkie
Developing imaginal discs are exposed to signals from an array of conserved developmental signaling pathways, some of which are proposed to affect dMyc levels or activity (e.g. Notch and Wg)39. Hence, a panel of reagents that detect activity changes in a variety of major cell pathways known to be active in the larval wing were assayed in en > ctp-IR discs (Figs 3 and S5). These included Su(H)-lacZ and E(spl)-mβ-CD2 (Notch pathway)40,41, PCNA-GFP and E2f1-lacZ (E2F/Rb pathway)42,43, EcRE-lacZ (EcR pathway)44, Stat-10xGFP (Jak-Stat pathway)45 and antibodies to the intracellular domain of Notch (N-icd), phospho-Mad (pMad), Wingless (Wg), the Wg/Notch regulated transcription factor Cut46, the EGFR pathway component diphospho-Erk (dpErk) and the G1/S regulator CyclinE47. None of these markers were evidently altered in the posterior domain of en > ctp-IR discs (Fig. 3C–K,M) or ctpex3 clones in the eye disc (Fig. 3L,N). In parallel genetic experiments, a loss-of-function allele of the pro-growth gene yorkie (yki) was found to dominantly enhance the en > ctp-IR small-wing phenotype (Fig. 4A). The Yki protein is the main target of the Hippo pathway and acts as nuclear co-activator for Scalloped (Sd)-dependent induction of Hippo target genes, which include dMyc48. An enlarged-wing phenotype produced by expression of a UAS-yki-V5 transgene from the en > Gal4 driver is significantly suppressed by co-depletion of Ctp (Fig. 4B). Likewise ctp knockdown can suppress eye overgrowth induced by GMR-Gal4 driven expression of YkiS168A, a hyperactive phospho-mutant form of Yki (Fig. 4C). Together, this evidence points towards a functional interaction between ctp and yki in disc cells destined to form the wing blade and eye.

Genetic interactions between ctp and yki in control of wing and eye size.
Box-plot representation of the effect of (A) the ykiB5 allele (n = 13), *p < 0.0001 or (B) a UAS-yki-V5 transgene (n = 22, with GFP n = 12, with ctpIR + GFP), on PCR in control (en > GFP, n = 12) or Ctp-depleted (en > ctpIR + GFP, n = 18) adult wings. All *p values < 0.0001. (C) Box-plot showing the effect of GMR-Gal4 driven Ctp-depletion at 29 °C among eye cells on final adult eye area (2-dimensional en face circumference) and the ability of the ctpIR transgene to suppress this metric in the background of the GMR-ykiS168A hyperactive allele. SEMs are indicated; (GMR, n = 24; GMR > ctpIR, n = 22; GMR > ykiS168A, n = 41; GMR > ykiS168A + ctpIR, n = 24), both *p values < 0.0001.
Ctp loss reduces thread/diap1 transcription
The proposed dynein-independent role of mammalian LC8/Ctp as a dimerization hub for cytoplasmic and nuclear complexes (reviewed in refs. 3,7) suggests that Ctp could be involved in modulating activity of the Hippo pathway in vivo. This hypothesis was tested by assessing expression of the thread(th)/diap1 gene, a key anti-apoptosis factor and canonical Yki transcriptional target23,49,50 in Ctp-depleted wing pouch cells. The steady-state level of Diap1 protein is reduced but not eliminated in the posterior compartment of en > ctp-IR larval discs (Fig. 5A,B), which could explain the mild increase in cC3 signal observed in ctp knockdown discs (see Fig. 2A,A′). The th-lacZ reporter, which is a Yki-responsive ‘trap’ of the bacterial β-galactosidase gene inserted into the endogenous th/diap1 locus51, also shows reduced expression in Ctp-depleted and ctpex3 mutant cells in the wing pouch (Fig. 5C,D,K,L and Figure S6E,F). A series of successively smaller promoter fragments of the th/diap1 promoter driving lacZ have been used to define a minimal Hippo response element (HRE) that responds to Yki hyperactivation in larval disc cells25. Expression of two of these reporters, 2b2-lacZ and 2b2c-lacZ, is strongly reduced in response to ctp knockdown (Fig. 5E–H). Baseline expression of the minimal HRE-lacZ is fairly low, but its expression is also reduced in Ctp-depleted cells (Fig. 5I,J with co-expression of dicer2 to enhance ctp knockdown and Figure S6A,B, without dicer2). Furthermore, a mutant version of the minimal HRE lacking the Sd-binding site necessary for Yki dependent transcription (HREmut6-lacZ)25 has lowered expression and no longer responds to loss of ctp (Figure S6C,D). In sum, these data provide evidence that Ctp is required to support transcription of the Yki target gene thread/diap1 in larval wing pouch cells.

Ctp supports expression of the diap-1/thread locus.
Confocal images of control (en > + in (A,C,E,G,I)) or Ctp-depleted (en > ctpIR in (B,D,F,H,J)) L3 wing discs stained for Ci (anterior domain) (A,B) and Diap1 proteins (A’,B’), or with anti-ßgal to detect expression of endogenous th-lacZ (C’,D’) as well as sequentially smaller Yki-responsive promoter elements 2B2-lacZ (E’,F’), 2B2C-lacZ (G’,H’) and Hippo response element (HRE)-lacZ (I’,J’). Co-expressed UAS-GFP transgene marks the posterior compartment (C–J) and the addition of dicer2 (+dcr) was used to enhance ctp knockdown in (I,J). Note decreased expression of all three Diap1 markers in cells depleted of Ctp in the posterior (right-side) compartment. (K,L) Heat-shock induced GFP-negative ctpex3 null clones in an L3 wing disc in the background of the th-lacZ reporter. Area in L is a magnified view of the anterior clone in K. Note the drop in th-lacZ expression in ctpex3 cells.
Ctp loss elevates transcription of the bantam microRNA locus
Analysis of a second well-validated Yki target, the pro-growth bantam (ban) microRNA, was carried out to determine whether the requirement for Ctp is unique to th/diap1 transcription, or can be extended to other well-validated Yki-target genes as well. Two ban transcriptional reporter transgenes were used for these studies: ban3-GFP, which contains a large proximal fragment of the ban promoter driving GFP and brC12-lacZ, a Yki-responsive 410-bp promoter fragment that lies within the ban3 region and contains two Yki-association sites27,52. Surprisingly and in polar contrast to th/diap1, expression of both ban reporters was strongly elevated in Ctp-depleted wing pouch cells (Fig. 6A–D). The brC12 reporter is also strongly induced in ctpex3 null wing pouch clones (Fig. 6E,F; two independent discs are shown with multiple ctpex3 clones), confirming the link between endogenous Ctp and ban promoter activity. Thus, while Ctp normally supports expression of the Yki-response gene th/diap1 in wing pouch cells, it has the inverse role of repressing transcription of the Yki-responsive locus ban. A third classic Yki reporter, expanded-lacZ, did not respond to Ctp-loss in otherwise wildtype larval wing cells or in those overexpressing Yki (Figure S7).

Ctp represses expression of the bantam locus.
Confocal images of control (en > + in (A,C)) or Ctp-depleted (en > ctpIR in (B,D)) L3 wing discs in the background of the bantam reporters ban3-GFP (A’,B’) or brC12-lacZ (C’,D’). Anti-Ci marks anterior cells in (A,B) while a co-expressed UAS-GFP transgene marks posterior cells in (C,D). Note increased expression of both ban3 and brC12 upon depletion of Ctp in the posterior (right-side) compartment. (E,F) Heat-shock induced GFP-negative ctpex3 null clones in an L3 wing disc in the background of the brC12-lacZ reporter. Two different discs are shown. Note consistently elevated brC12-lacZ expression in ctpex3 cells, particularly within the pouch.
Epistatic relationships between yki and ctp in control of th/diap1 and ban reporters
The opposing effects of Ctp loss on the minimally Hippo-responsive th/diap1 and ban promoter fragments imply an Yki-dependent mechanism links Ctp to expression of these genes. To test these relationships, the effect of Ctp loss on brC12-lacZ and th-lacZ were reassessed either in the presence of overexpressed Yki, or RNAi depletion of endogenous Yki. Elevated brC12-lacZ expression upon Ctp-depletion is suppressed by concurrent RNAi depletion of Yki, indicating that Yki is required for maximal induction of ban following Ctp loss (Fig. 7A). Furthermore, expression of a UAS-yki transgene does not obviously elevate induction brC12-lacZ above the level observed in ctpex3 null clones (Figure S8A,A′). Indeed within the center of the L3 wing pouch, brC12-lacZ is more highly expressed in ctp null cells than in those expressing a yki transgene (Figure S8B; compare centrally located ctpex3 clone vs. surrounding Yki-expressing cells in the posterior domain). Thus, Ctp-depletion robustly activates brC12-lacZ expression in pouch cells via a mechanism that requires Yki. Because overexpression of Ctp in wing pouch cells via UAS-ctp has no effect on wing growth or Yki readouts (data not shown), the epistatic relationship between ctp and yki on the diap-1 promoter is more challenging to assess. However, transgenic expression of UAS-yki remains capable of inducing th-lacZ expression in wing disc cells depleted of Ctp (Fig. 7I–L), suggesting that ctp acts either upstream or parallel to yki. Thus, manipulating Yki levels modifies the effects of Ctp depletion on both th/diap1 and ban reporters, which provides some evidence that ctp inversely modulates Yki activity toward each of these target genes.

Epistatic relationship between Yki and Ctp-regulated growth.
(A–D) Anti-βgal staining of L3 wing discs to detect brC12-lacZ expression in the background of (A’) control en > GFP; (B’) Ctp-depleted, en > GFP + ctpIR; (C’) Yki-depleted, en > GFP + ykiIR; or (D’) simultaneous depletion of Yki and Ctp, en > GFP + ctpIR + ykiIR. Co-expressed UAS-GFP transgene marks the posterior compartment (A–D). (E,F) Quantitation of the effect of Ctp and/or Yki depletion on brC12-lacZ expression determined by measuring anti-βgal pixel fluorescence intensity across the anterior:posterior (A:P) boundary (left-to-right) in the white boxes in (A–D). GFP-expression domain is indicated. Average fluorescence of the anterior domain was set to zero (0). (E) ■ en > GFP, Δ en > GFP + ctpIR. (F) ○ en > GFP + ykiIR, ▼ en > GFP + ykiIR + ctpIR. (G–J) Anti-ßgal staining of L3 wing discs to detect th-lacZ expression in the background of (G’) control en > GFP; (H’) en > GFP + ctpIR Ctp-depleted; (I’) en > GFP + ykiWT Yki-overexpression; or (J’) simultaneous expression of Yki and depletion of Ctp, en > GFP + ykiWT + ctpIR. Co-expressed UAS-GFP transgene marks the posterior compartment (G–J). (K,L) Quantitation of the effect of Ctp depletion and/or Yki overexpression on th-lacZ expression determined by measuring anti-ßgal pixel fluorescence intensity across the anterior:posterior (A:P) boundary (left-to-right) in the white boxes in (G–J) ⋄ en > GFP, ▲ en > GFP + ctpIR. (L) ● en > GFP + ykiWT, □ en > GFP + ykiWT + ctpIR.
Wing disc cells lacking the Yki-binding protein Myopic also show selective effects on Yki-target genes53. Myopic tethers Yki to endosomes for eventual degradation in lysosomes and its loss causes Yki to accumulate, leading to induction of ex and ban but not thread/diap153. To test whether Ctp is required for Yki cytoplasmic trafficking in a manner similar to Myopic, endogenous Yki and a V5-epitope tagged form of Yki were visualized in Ctp-depleted pouch cells (Figure S9). Yki steady state levels and nuclear:cytoplasmic distribution are unaltered in Ctp-depleted cells, indicating that wholesale changes in Yki protein dynamics and trafficking in ctp mutant wing pouch cells are unlikely to drive downstream effects on th/diap1 and ban expression.
Discussion
Here we define a role for the Drosophila protein Ctp, a member of the LC8 protein family, in regulating expression of two Hippo target genes, th/diap1 and bantam, in larval disc cells. Ctp is a member of a highly conserved family (Ctp, Dlc1 and DYNNL1/DYNNL2) of small proteins that were first identified as components of cytoplasmic dynein motors1,5,6 but are now recognized to also act as interaction hubs for many proteins with roles in diverse processes such as autophagy, signal transduction, cell:cell adhesion and transcription3,7,8. Largely because of this diverse set of potential effector pathways, the role of LC8 proteins in specific cellular and developmental contexts is not particularly well defined. We find that larval wing compartments depleted of Ctp by RNAi give rise to smaller compartments in the adult wing that are populated by smaller cells. Importantly, the Ctp-depleted larval precursors of these adult wing cells express markers of both M and S-phase and thus do not appear to undergo cell cycle arrest as reported elsewhere. Parallel analysis with a ctp genomic allele confirms that Ctp supports clonal growth in the larval wing disc, particularly in the pouch and that levels of the pro-growth transcription factor dMyc are reduced in ctp mutant epithelial cells. Testing candidate growth regulatory pathways active in the wing pouch uncovers a specific role for Ctp in regulating expression of genes regulated by the Hippo pathway target Yorkie (Yki), which is can positively regulate dMyc transcription. Activity reporters of two Yki-responsive genes, thread/diap1 and bantam, each respond to ctp loss in pouch cells and these effects map to smaller regions of the thread/diap1 and bantam promoters that contain Yki-responsive elements. Reduction of Yki activity enhances the Ctp-small wing phenotype and depletion of Ctp in turn blunts eye and wing growth driven by yki transgenes. These data are consistent with Ctp normally regulating Yki activity and Yki-dependent growth in imaginal disc epithelia.
The effects of Ctp on growth and Yki-target gene transcription are complex and intriguing. While Ctp supports Yki activity on the thread/diap1 promoter, it restricts activity of the bantam promoter and has no effect on a third Yki reporter, ex-lacZ. Moreover transgenic expression of ban drives tissue overgrowth54,55 but upregulation of ban transcription in ctp mutant cells is paradoxically associated with tissue undergrowth. This apparent ban paradox may be explained by the finding that ban promotes tissue growth through dMyc36,56, so that Ctp-depleted cells with reduced dMyc levels may be resistant to ban-induced growth.
The opposing effects of Ctp on thread/diap1 and ban transcription differ from core Hippo components, which affect these targets in a uniform way. The mechanism through which Ctp achieves its unique effects on thread/diap1 and bantam is not known. Intriguingly human LC8 interacts with the Kibra protein, a conserved element of the Hippo pathway15. Drosophila Kibra forms a complex with Merlin and Expanded and together these proteins promote phosphorylation of Warts57. However Warts uniformly represses expression of Yki-target genes, making it unlikely that Ctp acts via a Kibra-Warts axis to exert inverse effects on thread/diap1 and ban. The Kibra-LC8 complex interacts with Sorting Nexin-4 (Snx4) to promote dynein-driven traffic of cargo between the early and recycling endosomal compartments17. Yki associates with endosomes53 and endosomal traffic modulates levels of Yki protein and the transcription of its nuclear targets53,58. However, Ctp loss does not obviously alter steady state levels of Yki or its distribution in wing disc cells (see Fig. 7) and although vertebrate LC8 (aka DYNLL1) is as a high-confidence interactor of the Yki vertebrate homolog Yap159,60, an equivalent association was not detectable in cultured Drosophila S2 cells (data not shown).
In addition to cytoplasmic effects, Ctp/LC8 proteins also have nuclear roles in transcriptional control3. For example, LC8 interacts with the estrogen receptor (ER), can promotes ER activity and is found with ER on the promoters of induced genes14. A recent study confirmed that ER dimerizes when bound to DNA and can activate target gene transcription either as a monomer or dimer61. Drosophila Ctp may thus interact with Yki, or alternatively with a nuclear effector in a distinct pathway that converges on Yki, to bias Yki promoter selectivity and /or to modulate formation of higher order transcriptional complexes with promoter-specific roles in vivo. Future studies will be required to identify Ctp interacting proteins within the Hippo pathway and to resolve the precise link between Yki and the multifunctional and highly conserved Ctp/LC8 protein.
Additional Information
How to cite this article: Barron, D. A. and Moberg, K. Inverse regulation of two classic Hippo pathway target genes in Drosophila by the dimerization hub protein Ctp. Sci. Rep. 6, 22726; doi: 10.1038/srep22726 (2016).
References
King, S. M. et al. Brain cytoplasmic and flagellar outer arm dyneins share a highly conserved Mr 8,000 light chain. J Biol Chem 271, 19358–66 (1996).
Dick, T., Ray, K., Salz, H. K. & Chia, W. Cytoplasmic dynein (ddlc1) mutations cause morphogenetic defects and apoptotic cell death in Drosophila melanogaster. Mol Cell Biol 16, 1966–77 (1996).
Rapali, P. et al. DYNLL/LC8: a light chain subunit of the dynein motor complex and beyond. FEBS J 278, 2980–96 (2011).
Kamath, R. S. et al. Systematic functional analysis of the Caenorhabditis elegans genome using RNAi. Nature 421, 231–7 (2003).
Stuchell-Brereton, M. D. et al. Functional interaction between dynein light chain and intermediate chain is required for mitotic spindle positioning. Mol Biol Cell 22, 2690–701 (2011).
Williams, J. C. et al. Structural and thermodynamic characterization of a cytoplasmic dynein light chain-intermediate chain complex. Proc Natl Acad Sci USA 104, 10028–33 (2007).
Barbar, E. Dynein light chain LC8 is a dimerization hub essential in diverse protein networks. Biochemistry 47, 503–8 (2008).
Rapali, P. et al. Directed evolution reveals the binding motif preference of the LC8/DYNLL hub protein and predicts large numbers of novel binders in the human proteome. PLoS One 6, e18818 (2011).
Wickstead, B. & Gull, K. Dyneins across eukaryotes: a comparative genomic analysis. Traffic 8, 1708–21 (2007).
Rapali, P. et al. LC8 dynein light chain (DYNLL1) binds to the C-terminal domain of ATM-interacting protein (ATMIN/ASCIZ) and regulates its subcellular localization. Biochem Biophys Res Commun 414, 493–8 (2011).
Puthalakath, H., Huang, D. C., O’Reilly, L. A., King, S. M. & Strasser, A. The proapoptotic activity of the Bcl-2 family member Bim is regulated by interaction with the dynein motor complex. Mol Cell 3, 287–96 (1999).
Vadlamudi, R. K. et al. Dynein light chain 1, a p21-activated kinase 1-interacting substrate, promotes cancerous phenotypes. Cancer Cell 5, 575–85 (2004).
den Hollander, P. & Kumar, R. Dynein light chain 1 contributes to cell cycle progression by increasing cyclin-dependent kinase 2 activity in estrogen-stimulated cells. Cancer Res 66, 5941–9 (2006).
Rayala, S. K. et al. Functional regulation of oestrogen receptor pathway by the dynein light chain 1. EMBO Rep 6, 538–44 (2005).
Rayala, S. K. et al. Essential role of KIBRA in co-activator function of dynein light chain 1 in mammalian cells. J Biol Chem 281, 19092–9 (2006).
Xiao, L., Chen, Y., Ji, M. & Dong, J. KIBRA regulates Hippo signaling activity via interactions with large tumor suppressor kinases. J Biol Chem 286, 7788–96 (2011).
Traer, C. J. et al. SNX4 coordinates endosomal sorting of TfnR with dynein-mediated transport into the endocytic recycling compartment. Nat Cell Biol 9, 1370–80 (2007).
Phillis, R., Statton, D., Caruccio, P. & Murphey, R. K. Mutations in the 8 kDa dynein light chain gene disrupt sensory axon projections in the Drosophila imaginal CNS. Development 122, 2955–63 (1996).
Batlevi, Y. et al. Dynein light chain 1 is required for autophagy, protein clearance and cell death in Drosophila. Proc Natl Acad Sci USA 107, 742–7 (2010).
Wang, C. et al. An ana2/ctp/mud complex regulates spindle orientation in Drosophila neuroblasts. Dev Cell 21, 520–33 (2011).
Joti, P., Ghosh-Roy, A. & Ray, K. Dynein light chain 1 functions in somatic cyst cells regulate spermatogonial divisions in Drosophila. Sci Rep 1, 173 (2011).
Zaytseva, O. et al. The novel zinc finger protein dASCIZ regulates mitosis in Drosophila via an essential role in dynein light-chain expression. Genetics 196, 443–53 (2014).
Wu, S., Huang, J., Dong, J. & Pan, D. hippo encodes a Ste-20 family protein kinase that restricts cell proliferation and promotes apoptosis in conjunction with salvador and warts. Cell 114, 445–56 (2003).
Zhang, L. et al. The TEAD/TEF family of transcription factor Scalloped mediates Hippo signaling in organ size control. Dev Cell 14, 377–87 (2008).
Wu, S., Liu, Y., Zheng, Y., Dong, J. & Pan, D. The TEAD/TEF family protein Scalloped mediates transcriptional output of the Hippo growth-regulatory pathway. Dev Cell 14, 388–98 (2008).
Oh, H. & Irvine, K. D. In vivo regulation of Yorkie phosphorylation and localization. Development 135, 1081–8 (2008).
Oh, H. & Irvine, K. D. Cooperative regulation of growth by Yorkie and Mad through bantam. Dev Cell 20, 109–22 (2011).
Moberg, K. H., Mukherjee, A., Veraksa, A., Artavanis-Tsakonas, S. & Hariharan, I. K. The Drosophila F box protein archipelago regulates dMyc protein levels in vivo. Curr Biol 14, 965–74 (2004).
Morrison, C. M. & Halder, G. Characterization of a dorsal-eye Gal4 Line in Drosophila. Genesis 48, 3–7 (2010).
Xue, D. & Horvitz, H. R. Inhibition of the Caenorhabditis elegans cell-death protease CED-3 by a CED-3 cleavage site in baculovirus p35 protein. Nature 377, 248–51 (1995).
Varma, D. et al. Development and application of in vivo molecular traps reveals that dynein light chain occupancy differentially affects dynein-mediated processes. Proc Natl Acad Sci USA 107, 3493–8 (2010).
Neumuller, R. A. et al. Genome-wide analysis of self-renewal in Drosophila neural stem cells by transgenic RNAi. Cell Stem Cell 8, 580–93 (2011).
Schreiber-Agus, N. et al. Drosophila Myc is oncogenic in mammalian cells and plays a role in the diminutive phenotype. Proc Natl Acad Sci USA 94, 1235–40 (1997).
Marygold, S. J. et al. The ribosomal protein genes and Minute loci of Drosophila melanogaster. Genome Biol 8, R216 (2007).
Johnston, L. A., Prober, D. A., Edgar, B. A., Eisenman, R. N. & Gallant, P. Drosophila myc regulates cellular growth during development. Cell 98, 779–90 (1999).
Herranz, H. et al. The miRNA machinery targets Mei-P26 and regulates Myc protein levels in the Drosophila wing. EMBO J 29, 1688–98 (2010).
Quinn, L. M. et al. Drosophila Hfp negatively regulates dmyc and stg to inhibit cell proliferation. Development 131, 1411–23 (2004).
Antonucci, L. et al. CNBP regulates wing development in Drosophila melanogaster by promoting IRES-dependent translation of dMyc. Cell Cycle 13, 434–9 (2014).
Herranz, H., Perez, L., Martin, F. A. & Milan, M. A Wingless and Notch double-repression mechanism regulates G1-S transition in the Drosophila wing. EMBO J 27, 1633–45 (2008).
Barolo, S. et al. A notch-independent activity of suppressor of hairless is required for normal mechanoreceptor physiology. Cell 103, 957–69 (2000).
de Celis, J. F., Tyler, D. M., de Celis, J. & Bray, S. J. Notch signalling mediates segmentation of the Drosophila leg. Development 125, 4617–26 (1998).
Thacker, S. A., Bonnette, P. C. & Duronio, R. J. The contribution of E2F-regulated transcription to Drosophila PCNA gene function. Curr Biol 13, 53–8 (2003).
Duronio, R. J., O’Farrell, P. H., Xie, J. E., Brook, A. & Dyson, N. The transcription factor E2F is required for S phase during Drosophila embryogenesis. Genes Dev 9, 1445–55 (1995).
Brennan, C. A., Ashburner, M. & Moses, K. Ecdysone pathway is required for furrow progression in the developing Drosophila eye. Development 125, 2653–64 (1998).
Bach, E. A. et al. GFP reporters detect the activation of the Drosophila JAK/STAT pathway in vivo. Gene Expr Patterns 7, 323–31 (2007).
Nepveu, A. Role of the multifunctional CDP/Cut/Cux homeodomain transcription factor in regulating differentiation, cell growth and development. Gene 270, 1–15 (2001).
Duronio, R. J. & O’Farrell, P. H. Developmental control of a G1-S transcriptional program in Drosophila. Development 120, 1503–15 (1994).
Ziosi, M. et al. dMyc functions downstream of Yorkie to promote the supercompetitive behavior of hippo pathway mutant cells. PLoS Genet 6, e1001140 (2010).
Huang, J., Wu, S., Barrera, J., Matthews, K. & Pan, D. The Hippo signaling pathway coordinately regulates cell proliferation and apoptosis by inactivating Yorkie, the Drosophila Homolog of YAP. Cell 122, 421–34 (2005).
Jia, J., Zhang, W., Wang, B., Trinko, R. & Jiang, J. The Drosophila Ste20 family kinase dMST functions as a tumor suppressor by restricting cell proliferation and promoting apoptosis. Genes Dev 17, 2514–9 (2003).
Hay, B. A., Wassarman, D. A. & Rubin, G. M. Drosophila homologs of baculovirus inhibitor of apoptosis proteins function to block cell death. Cell 83, 1253–62 (1995).
Matakatsu, H. & Blair, S. S. Separating planar cell polarity and Hippo pathway activities of the protocadherins Fat and Dachsous. Development 139, 1498–508 (2012).
Gilbert, M. M., Tipping, M., Veraksa, A. & Moberg, K. H. A Screen for Conditional Growth Suppressor Genes Identifies the Drosophila Homolog of HD-PTP as a Regulator of the Oncoprotein Yorkie. Dev Cell 20, 700–12 (2011).
Hipfner, D. R., Weigmann, K. & Cohen, S. M. The bantam gene regulates Drosophila growth. Genetics 161, 1527–37 (2002).
Brennecke, J., Hipfner, D. R., Stark, A., Russell, R. B. & Cohen, S. M. bantam encodes a developmentally regulated microRNA that controls cell proliferation and regulates the proapoptotic gene hid in Drosophila. Cell 113, 25–36 (2003).
Reddy, B. V. & Irvine, K. D. Regulation of Drosophila glial cell proliferation by Merlin-Hippo signaling. Development 138, 5201–12 (2011).
Yu, J. et al. Kibra functions as a tumor suppressor protein that regulates Hippo signaling in conjunction with Merlin and Expanded. Dev Cell 18, 288–99 (2010).
Kwon, Y. et al. The Hippo signaling pathway interactome. Science 342, 737–40 (2013).
Couzens, A. L. et al. Protein interaction network of the mammalian Hippo pathway reveals mechanisms of kinase-phosphatase interactions. Sci Signal 6, rs15 (2013).
Wang, W. et al. Defining the protein-protein interaction network of the human hippo pathway. Mol Cell Proteomics 13, 119–31 (2014).
Yi, P. et al. Structure of a biologically active estrogen receptor-coactivator complex on DNA. Mol Cell 57, 1047–58 (2015).
Acknowledgements
We thank the Bloomington Drosophila Stock Center (BDSC) and the Developmental Studies Hybridoma Bank (DSHB) for fly stocks and antibodies respectively. We thank W. Sale for the anti-LC8 antibody, W. Chia and E. for the ctpex3 allele, K. Irvine for the bantam promoter reporter lines, D.J. Pan for the thread/diap1 promoter reporter lines and E. Bach for the 10xStat-GFP line. We are especially grateful to H. Zhao (U. Mass./Boston) for help with protein interaction experiments. We also thank members of the Moberg, Veraksa and Read labs for helpful discussions and reagents. Finally, we apologize to those we could not cite due to space constraints. This work was funded by NIH grants CA123368 and GM105813 to KHM and by F30-CA177156 to D.A.B.
Author information
Authors and Affiliations
Contributions
K.M. and D.B. wrote and reviewed the main manuscript and D.B. carried out all experiments and prepared all figures.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Barron, D., Moberg, K. Inverse regulation of two classic Hippo pathway target genes in Drosophila by the dimerization hub protein Ctp. Sci Rep 6, 22726 (2016). https://doi.org/10.1038/srep22726
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep22726
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
