
Abstract
Psittacosauridae (parrot-beaked dinosaurs) represents the first major radiation of ceratopsians (horned dinosaurs). However, psittacosaurids are divergent from the general morphology found in other ceratopsians and this has resulted in their uncertain systematic position among ceratopsians. Here we describe a new basal neoceratopsian dinosaur, Mosaiceratops azumai gen. et sp. nov. based on a partial semi-articulated skeleton recovered from the Upper Cretaceous Xiaguan Formation of Neixiang County, Henan Province, China. Although our phylogenetic analysis supports this taxon as the most basal neoceratopsian, Mosaiceratops exhibits many features previously considered unique to the Psittacosauridae among the basal Ceratopsia. These include a relatively highly positioned external naris, a proportionally large premaxilla, the nasal extending ventral to the external naris, slender postorbital and temporal bars, a large notch between the basal tubera and the edentulous premaxilla. Thus, the discovery of Mosaiceratops reduces the morphological disparity between the Psittacosauridae and other basal ceratopsians. Character optimization suggests that basal neoceratopsians have re-evolved premaxillary teeth; a major reversal previously unknown in any dinosaur clade. The new specimen also highlights the mosaic nature of evolution among early ceratopsians and supports the phylogenetic hypothesis that the Psittacosauridae is a relatively derived clade, rather than the most basal group of the Ceratopsia.
Similar content being viewed by others

Introduction
Ceratopsians are a group of ornithischian dinosaurs characterized by a number of features including a large head, a skull with narrow beak and flaring jugals, a rostral bone, a bony frill and other characters1,2,3. Among the known ceratopsians, Psittacosauridae is the most unusual clade because it differs from other ceratopsians in many features, such as its bipedal posture, high and short snout, highly positioned external naris and reduced lateral manual digits4,5. The significant morphological differences between the psittacosaurids and other ceratopsians hampered the recognition of the ceratopsian affinity of the Psittacosauridae when they were first discovered6,7,8. The recent discovery of Jurassic ceratopsians has helped to reduce the morphological gap between the Psittacosauridae and other ceratopsians9,10,11,12. However, significant morphological differences still remain. As a result, the systematic position of the Psittacosauridae remains controversial with some studies considering them as the most basal ceratopsian clade5,13,14 and others placing the group as intermediate between Jurassic forms such as Chaoyangsauridae and other Cretaceous taxa2,12.
Here we report a new basal neoceratopsian based on a specimen recovered from the Upper Cretaceous Xiaguan Formation of Neixiang County, Henan Province, China. The locality is on the west bank of Tuanhe River lies in the center of the Xiaguan-Gaoqiu Basin, one of several Cretaceous terrestrial basins in southwestern Henan Province. Xiaguan-Gaoqiu Basin previously has yielded a large number of fossil eggs and two dinosaur taxa, the basal hadrosauroid Nanyangosaurus15 and the titanosauriform Baotianmansaurus16. The fossil-bearing horizon in the Xiaguan-Gaoqiu Basin has been assigned variously to the Sangping15, Gaogou16 and Majiachun formations17. Despite those previous assessments, the horizon is now placed in the Xiaguan Formation17,18. The horizon lacks radiometric dates, but is believed to fall between the early-middle Turonian and the middle Campanian of the Late Cretaceous, based on the plant and invertebrate fossils17. This new specimen displays a mosaic combination of features, some of which were previously considered diagnostic of a certain ceratopsian clade, notably the Psittacosauridae. This specimen represents a significant discovery for understanding the early evolution of ceratopsian dinosaurs. The new information provided by this specimen reshapes our hypotheses of the evolution of several features in ceratopsian evolution.
Results
Systematic palaeontology
Dinosauria Owen, 1842
Ornithischia Seeley, 1887
Ceratopsia Marsh, 1890
Neoceratopsia Sereno, 1986
Mosaiceratops azumai gen. et sp. nov.
Etymology
The generic name Mosaiceratops (“mosaic ceratopsian”) is a contraction of the Latin terms “mosaicus” and “ceratops” in reference to the specimen’s unique (mosaic) combination of characters that were previously considered diagnostic of basal ceratopsians, psittacosaurids, or basal neoceratopsians. The specific name honors Dr. Yoichi Azuma from Fukui Prefectural Dinosaur Museum, who co-organized and participated in several dinosaur expeditions in China. One of those expeditions led to the discovery of the basal neoceratopsian Archaeoceratops19.
Holotype
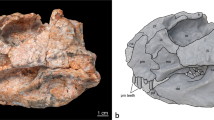
ZMNH M8856 (housed in the collections of the Zhejiang Museum of Natural History, Hangzhou, China) (Figs 1 and 2) is composed of a partial skull and associated partial postcranial skeleton that includes three cervical vertebrae, three dorsal vertebrae, dorsal ribs, 18 caudal vertebrae, chevrons, the right humerus, the radius, both ilia, the left ischium, both femora, both tibiae, the left fibula, the left astragalus, the left calcaneum, metatarsals, phalanges and some undiagnostic remains.

Holotype and skeletal reconstruction of Mosaiceratops azumai, gen. et sp. nov (ZMNH M8856).
(a) photograph and line drawing of ZMNH M8856; (b) skeletal reconstruction showing preserved elements in white. Scale bar 10 cm. Abbreviations: a, astragalus; boc, basioccipital; c, calcaneum; cav, caudal vertebra; ch, chevron; cv, cervical vertebra; dr, dorsal rib; dv, dorsal vertebra; f, frontal; fem, femur; fl, fibula; h, humerus; il, ilium; is, ischium; L, left; mt, metatarsal; ph, phalanx/phalanges; po, postorbital; R, right; sk, skull; sq, squamosal; t, tibia; td, tendon; ?, undiagnostic remains.

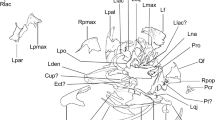
Cranial remains of Mosaiceratops azumai, gen. et sp. nov.
Photograph and interpretive drawing of skull in left lateral (a), right lateral (b) and ventral (c) views; right postorbital in dorsal (d) and anterolateral (e) views; left squamosal in lateral (f) and laterodorsal (g) views; partial braincase in occipital (h) view. Scale bars in a–c, 5 cm, in d–h, 2 cm. Abbreviations: af, antorbital fossa; an, angular; ar, articular; boc, basioccipital; bt, basioccipital tubera; d, dentary; dp, dorsal process of rostral; en, external naris; exo, exoccipital; fm, foramen magnum; g, groove; itf, infratemporal fenestra; j, jugal; jp, jugal process; lp, lateral process; mp, medial process; mx, maxilla; n, notch; na, nasal; o, orbit; oc, occipital condyle; pd, predentary; pmx, premaxilla; pop, postorbital process; prf, prefrontal; prfp, prefrontal process; qj, quadratojugal; r, rostral bone; sa, surangular; spl, splenial; sqp, squamosal process; tb, tubercle; vp, ventral process.
Locality and horizon
Upper Cretaceous (lower-middle Turonian—middle Campanian), Xiaguan Formation, Neixiang County, Henan Province, China.
Diagnosis
Small basal neoceratopsian with the following autapomorphies: curved groove anterior to the contact between the premaxilla and maxilla; elongate anterior process of jugal with sub-parallel dorsal and ventral edges; two tubercles developed in the ventral border infraorbital ramus; and the distal end of the dorsal ilium edge dorsoposteriorly directed. Mosaiceratops also differs from other basal neoceratopsians in the following features: edentulous premaxilla; premaxilla slightly larger than maxilla in lateral view; nasal extended ventral to the external naris; two tubercles close to the ventral border of the infraorbital ramus of the jugal; an inverted T-shape jugal in lateral view; a T-shape postorbital with slender and elongate jugal and squamosal processes; medial process of squamosal anteromedially directed; anterior process of the squamosal simple, rather than bifid; and basal tubera separated by a deep middle notch.
Description and comparisons
The skull has the characteristic ceratopsian triangular morphology in dorsal view, with a narrow beak and strongly flaring jugal (Fig. 2c). The external naris sits relatively high with its ventral border slightly higher than the ventral edge of the orbit. Although incomplete, the external naris seems elongate, as characteristic of other basal neoceratopsians. In contrast whereas the naris is circular in Psittacosaurus5. The antorbital fossa is subtriangular in outline with sharp ventral and posterior rims as in Archaeoceratops20, Leptoceratops and Protoceratops14 and the antorbital fossa is absent in Psittacosaurus5. The ventral border of the infratemporal fenestra sits lower than the ventral border of the orbit, similar to other neoceratopsians such as Liaoceratops and Archaeoceratops, but contrasts with that of Psittacosaurus5.
The rostral extends anteriorly beyond the anterior tip of the lower jaw and curves strongly ventrally in lateral view (Fig. 2a). It appears round and unkeeled (Fig. 2c), similar to Chaoyangsaurus10, Psittacosaurus5, Liaoceratops12 and Yamaceratops21. In contrast, the rostral is keeled in most neoceratopsians, e.g. Archaeoceratops20. As in other neoceratopsians, the rostral bears lateral processes that extend posteriorly along the ventral margin of the premaxilla21.
The premaxilla appears subrectangular with a deeper posterior portion more closely resembling Archaeoceratops than those of other basal neoceratopsians (Fig. 2a)20. However, it is proportionally longer and lower than Archaeoceratops. As in psittacosaurids, the premaxilla is larger than the maxilla in lateral view. This differs from the condition in other ceratopsians such as Liaoceratops12, Archaeoceratops19 and Auroraceratops22, in which the premaxilla is smaller. The premaxillae and rostral bones form sharp, edentulous cutting margins (Fig. 2a–c), a feature similar to Psittacosaurus but not other basal ceratopsians5. A curved groove lies anterior to the premaxillary−maxillary suture, with its dorsal and ventral ends bending toward the posterior end of the premaxilla dorsally and ventrally, respectively. The maxilla widens posteriorly beneath the antorbital fossa (Fig. 2a). The dentigerous margin of the maxilla is inset more strongly than that of basal ceratopsians, such as Yinlong9 and Chaoyangsaurus10. More than ten teeth are present in the left maxilla, in addition to two disarticulated teeth preserved adjacent to the maxilla.
The external surface of the nasal has a roughened texture with no indication of a nasal horn, similar to other basal ceratopsians5. The nasal extends anteroventrally below the external naris, as in Psittacosaurus5.
The shape of the jugal resembles an inverted T in lateral view (Fig. 2a), similar to basal ceratopsians and Psittacosaurus. The infraorbital ramus of the jugal is shallow, elongate and its dorsal and ventral edges are sub-parallel to each other in lateral view (Fig. 2a). In other basal neoceratopsians, e.g. Archaeoceratops20 and Yamaceratops21, the infraorbital ramus is deep near the base of the dorsal process and tapers anteriorly with a sub-triangular morphology in lateral view. The lateral surface bears a distinct bumpy ornamentation. There are two tubercles close to the ventral border of the infraorbital ramus that are not present in other neoceratopsians (Fig. 2a). The jugal bears a low horn that lacks a separate epijugal, as in Liaoceratops12. As in the neoceratopsians more derived than Liaoceratops, the quadratojugal appears more prominent in occipital than lateral view21.
The triradiate postorbital retains elongate jugal and squamosal processes and a shortened prefrontal process (Fig. 2d,e), as in Psittacosaurus. In contrast, the postorbital in basal neoceratopsians is a triangular plate of bone, the jugal process is shorter and the squamosal process is more stout5. The postorbital process of the squamosal does not bifurcate (Fig. 2f,g), like the condition in Yinlong9 and Psittacosaurus5. However, the postorbital process is bifid in the other basal neoceratopsians, such as Liaoceratops, Archaeoceratops and the leptoceratopsids21. Although broken, the medial ramus is anteromedially directed (Fig. 2g), similar to that in Yinlong9 and Psittacosaurus5.
The foramen magnum is slightly larger than the occipital condyle, in contrast to the condition in Montanoceratops (Fig. 2h)23. The occipital condyle shares the spherical shape and distinct neck with other neoceratopsians5. The basioccipital flares laterally below the condyle to form the large basal tubera that is characteristic of ceratopsians23. A shallow groove occurs in the midline of the tubera and is also present in Montanoceratops23. In contrast, a longitudinal ridge is present below the condyle in Yinlong9 and other basal neoceratopsians, such as Liaoceratops24 and Archaeoceratops19. The basal tubera are widely separated by a middle notch, similar to Psittacosaurus25,26,27,28. This notch is absent or incipient in other neoceratopsians including Archaeoceratops20 and Auroraceratops25.
The mandibular ramus is shallow as in other basal neoceratopsians (Fig. 2a). In ventral view, the mandibular ramus expands posteriorly (Fig. 2c), resembling other basal neoceratopsians. The mandibular ramus appears nearly straight in dorsal view in Yinlong, Psittacosaurus and Chaoyangsaurus29. The ratio of the length posterior to the apex of the coronoid process to total mandibular length is approximately 0.35, similar to other basal neoceratopsians. This ratio is smaller than that in more basal ceratopsians and Psittacosaurus29.
The relatively long predentary possesses a strongly upturned and acutely pointed anterior tip that fits inside the upper beak (Fig. 2a). That feature is present in most other neoceratopsians including Archaeoceratops20 and Auroraceratops25. In contrast, the predentary is short and blunt in the basal neoceratopsian Liaoceratops24 and the more basal ceratopsians Yinlong9, Chaoyangsaurus10 and Psittacosaurus5. The dorsal edges are beveled, a feature present in many ceratopsians. In ventral view, the predentary displays a flattened ventromedial process that bifurcates posteriorly (Fig. 2c). The anteroventral surface accommodates a sharp keel at the midline. The dentary bears an everted flange extending laterally (Fig. 2a), as in Liaoceratops12. This feature is well developed and directed ventrally in Psittacosaurus29. The coronoid process sits low but still relatively higher than that of basal ceratopsians and Psittacosaurus. The process is well developed and separated from the tooth row by a wide sulcus, unlike Chaoyangsaurus, Psittacosaurus and Liaoceratops. There are approximately eleven tooth positions in the dentary.
The splenial is restricted to the medial margin of the mandible, with little ventral exposure. The anterior end of the splenial reaches the posteroventral end of the mandibular symphysis (bifurcate point of the predentary). That condition is similar to Liaoceratops30 and Leptoceratops29, but contrasts with the state in Archaeoceratops29.
The surangular is shorter than the dentary (Fig. 2a) and their dorsal and lateral surfaces are almost perpendicular to each other. The surangular forms part of the glenoid fossa lateral to the articular and comprises more than half of the lateral cotyle of the glenoid, a condition common in basal neoceratopsians31. The retroarticular process is short. The medial and lateral cotyles of the glenoid are well separated by a ridge. The elliptical and gently dorsally concave, elevated glenoid surface contrasts with the flat articular surface of the mandibular condyle in Psittacosaurus5,32. Two small tubercles are present near the posterior end of the angular. The larger one occurs on the suture with the surangular and the smaller one occurs anteroventrally to the previous tubercle. A weak ridge is developed along these two tubercles and is confluent with the dentary flange.
The maxillary tooth crowns have a low ridge distal to the middle line labially, similar to Archaeoceratops33. The primary ridge is narrow and the width remains constant, like that in Psittacosaurus lujiatunensis, Psittacosaurus sinensis and Archaeoceratops33. In contrast, the primary ridges taper toward apex in other species of Psittacosaurus, Yinlong and Auroraceratops33. Two or three subsidiary ridges are present medial to the prominent primary ridge that are parallel to each other, but oblique to the primary ridge (as in Archaeoceratops33). The teeth are single rooted as in other non-ceratopsid ceratopsians like Chaoyangsaurus, Psittacosaurus33 and Leptoceratops34,35. The dentary tooth crowns have a primary ridge lingually with three to four distal secondary ridges. The primary ridge is narrow like other basal neoceratopsians (Archaeoceratops and Auroraceratops33). In contrast, the primary ridge is more prominent and bulbous in Psittacosaurus5,33. In one dentary tooth, distal secondary ridges are parallel with the primary ridge, but in other dentary teeth, the distal secondary ridges are directed obliquely to the primary ridge.
The neurocentral suture is closed in all three preserved cervical vertebrae. The anterior cervical displays a well-developed ventral keel that becomes weaker in the posterior cervical. In the caudal vertebrae, the transverse processes are not fully fused with the centrum. The longest neural spines are approximately 1.5 times that of the corresponding centrum length.
As in other basal neoceratopsians, the humeral head inclines medially in posterior view. It is hemispherical in proximal view and supported by a thickened tubercle that projects to the posterior surface. In contrast, the head is poorly defined in Psittacosaurus5. The relatively long deltopectoral crest broadly expands and faces anteriorly. The crest is limited to the proximal half of shaft in Mosaiceratops, whereas it extends to the distal half in ceratopsids35. As in other neoceratopsians31,34,36, the medial condyle extends slightly further than the lateral condyle.
The gently arched dorsal margin of the ilium has its apex over the acetabulum and the dorsal margin of the postacetabular process curves slightly dorsally at the posterior end (Fig. 1a). The postacetabular process in basal neoceratopsians normally curves ventrally towards the posterior end, as in Archaeoceratops20, Yamaceratops21 and Auroraceratops31. The narrow pubic peduncle projects anteroventrally and the much more robust ischial peduncle contains a large convex antitrochanter. The ischial shaft appears oval in cross section like Auroraceratops, but not laterally compressed as in Psittacosaurus lujiatunensis, Leptoceratops, Montanoceratops and Protoceratops31. The distal end of the ischium downturns slightly and swells with a rugose surface, similar to other basal neoceratopsians such as Auroraceratops37 and Protoceratops36. However, the ischium is robust and decurved more strongly in ceratopsids35.
The femur gently bows anteriorly in lateral view (Fig. 1a). The fourth trochanter is large and only slightly pendent, as in other non-ceratopsid ceratopsians5,35. The shaft of the tibia is straight and longer than the femur (Fig. 1a) and comparable to other basal ceratopsians. The opposite condition is present in ceratopsids35,38. The foot is gracile and elongate with a constricted metatarsus and elongate phalanges (Fig. 1a). The ungual phalanges are slender and sharply pointed, as in other basal ceratopsians14 such as Archaeoceratops20 and Leptoceratops34.
Methods
Phylogenetic analysis
In order to determine the systematic position of Mosaiceratops azumai, we conducted a phylogenetic analysis with a dataset modified from a recently published dataset for ceratopsian phylogeny39. The major modifications include 12 new characters, four additional taxa [Mosaiceratops, Auroraceratops22,31, Zhuchengceratops40 and Aquilops41] and the revision of some character coding (see Supplementary Information). The modified matrix is comprised of 159 characters and 26 taxa.
The analysis was carried out using a traditional search with the tree bisection reconnection algorithm in TNT version 1.142. The parameters were left at their default settings with the following exception: maximum trees in memory set to 10000, 1000 replicates were used with 10 trees saved per replication. The analysis resulted in one parsimonious tree (tree length = 339 steps, CI = 0.54 and RI = 0.71). Mosaiceratops is recovered as the most basal neoceratopsian. The Jurassic forms, such as Yinlong and the Chaoyangsauridae, are placed at the base of the Ceratopsia and the Psittacosauridae are positioned intermediate between the Jurassic forms and the Neoceratopsia (Fig. 3).

Temporal calibration of the single most parsimonious tree produced by phylogenetic analysis.
Numbers beneath nodes are Bremer support values. The geologic numerical ages and coloring follow International Chronostratigraphic Chart 2014/1048.
Stratigraphic fit analysis
Measures of stratigraphic fit to phylogeny were analyzed by calculating modified Manhattan stratigraphic measure (MSM*) in TNT version 1.142,43,44. Temporal calibrations used for fit analysis are provided in Supplementary Information.
Discussion
Mosaiceratops clearly is a member of the Ceratopsia based on the presence of a rostral bone, the narial fossa being separated by a flat margin from the ventral margin of the premaxilla, the laterally projected and crested jugal and the predentary with a wide ventral process. Furthermore, this new species can be placed in the Neoceratopsia based on the presence of several derived features known only in neoceratopsians: a rostral with a well-developed lateral process and a strongly ventrally curved ventral edge; a quadratojugal more prominent in occipital view than lateral view; a spherical occipital condyle; a predentary with a beveled dorsal margin; a predentary that is relatively long and keeled and has an acutely pointed and strongly upturned anterior tip; shortened post-dentary mandibular elements; a reduced retroarticular process; and the hemispherical humeral head extending onto the posterior surface of the humerus.
Mosaiceratops possesses several plesiomorphic features that are previously unknown in other basal neoceratopsians. For example, the postorbital is T-shape with an elongation of the jugal and squamosal processes (similar to Yinlong and Psittacosaurus). However, the postorbital is a triangular plate of bone with stout jugal and squamosal processes in other basal neoceratopsians. The jugal is an inverted T-shape in lateral view, like that in Yinlong and Psittacosaurus. However, the jugal appears sub-triangular in other basal neoceratopsians. The simple postorbital process of the squamosal in Mosaiceratops appears similar to Yinlong and Psittacosaurus and the postorbital inserts into the bifid anterior end of the squamosal in other basal neoceratopsians. The medial process of the squamosal angles anteromedially, as in Yinlong and Psittacosaurus. However, the medial process of the squamosal is directed posteromedially to form the lateral corner of the frill.
Mosaiceratops possesses several features previously only known in the Psittacosauridae among basal ceratopsians and other basal neoceratopsians. As in the Psittacosauridae, the external naris is positioned relatively high, the nasal extends ventral to the external naris and reaches the rostral and there is a large notch between the basal tubera. The most striking feature is the edentulous premaxilla in Mosaiceratops. Although derived ceratopsians such as most Leptoceratopsids (Montanoceratops, Prenoceratops, Leptoceratops and Udanoceratops)39, Bagaceratops6 and Ceratopsids35 lack premaxillary teeth, all other basal ceratopsians have premaxillary teeth. For example, Yinlong has three premaxillary teeth9, Archaeoceratops has three19 and Protoceratops has two36. Among the basal ceratopsians and basal neoceratopsians, only the psittacosaurids lack premaxillary teeth. Mosaiceratops represents the first known non-psittacosaurid taxon among basal ceratopsians that lacks premaxillary teeth. Character optimization indicates that the absence of premaxillary teeth is a diagnostic feature for a clade composed of the Psittacosauridae and the Neoceratopsia and the occurrence of premaxillary teeth in other basal neoceratopsians, more derived than Mosaiceratops, represents a reversal to the more primitive condition present in basal ceratopsians. The potential reappearance of premaxillary teeth among more derived neoceratopsians would be a major character reversal previously unknown in any dinosaur clade and the absence of premaxillary teeth in Mosaiceratops helps to indicate another example of the occurrence of major homoplasy in dinosaur evolution. One other example is the re-evolving of a robust, functional hallux in derived therizinosaurs45. Major reversals also are known in several other vertebrate taxa, for example, the re-evolution of lost digits in lizard Bachia46 and re-evolution of lost mandibular teeth in frogs47. However, the recovered systematic position of Mosaiceratops as the most basal neoceratopsian is not strongly supported (Bremer index of 3) in our phylogenetic analysis and the possibility that Mosaiceratops represents the sister taxon to Psittacosaurus exists. In that less parsimonious case, the loss of premaxillary teeth would characterize a much less inclusive clade and no major reversal would have occurred in ceratopsian evolution.
It should be noted that some features in Mosaiceratops display an intermediate condition between those of psittacosaurids and neoceratopsians. For example, the premaxilla is larger than the maxilla in lateral view, but not proportionally as large as in psittacosaurids. In the latter, the premaxilla occupies the majority of the lateral surface of the snout. In other neoceratopsians, the maxilla overshadows the premaxilla in size in lateral view. The external naris sits relatively high on the face, its ventral border slightly higher than the ventral edge of the orbit. The external naris sits high in psittacosaurids and lies just below the upper portion of orbit. However, in other neoceratopsians, the external naris is positioned lower than the orbit. In our specimen, the basal tubera are separated by a deep middle notch. In psittacosaurids, the basal tubera also are separated by a deep middle notch, but the notch is deeper and narrower. In some neoceratopsians, the ventral border of basal tubera is slightly concave or almost flat. Mosaiceratops exhibits a laterally directed jaw flange that also is present in Liaoceratops. However, the jaw flange is well developed in psittacosaurids and directed ventrally and it is absent among other neoceratopsians12.
The discovery of Mosaiceratops highlights the mosaic evolution of basal ceratopsians that was also demonstrated by the previous discovery of Liaoceratops12. Mosaiceratops combines several derived features seen across different ceratopsian clades while combining those traits with many psittacosaurid-like features. These mingled characters in this basal neoceratopsian reduce the morphologic gap between the Psittacosauridae and other basal ceratopsians, but suggests a couple of evolutionary scenarios. If Mosaiceratops is the most basal neoceratopsian, then certain features (discussed above) are plesiomorphic for neoceratopsians rather than being derived solely for psittacosaurids and others have a distinctly homoplastic distribution. On the other hand, if Mosaiceratops is actually a basal psittacosaurid (the less parsimonious hypothesis), then it would suggest that psittacosaurids evolved from a much more neoceratopsian-like ancestor than currently thought.
The Early Cretaceous Psittacosauridae have been considered intermediate in phylogenetic position between the Jurassic ceratopsians, such as Yinlong and the Chaoyangsauridae and all other Cretaceous ceratopsians (i.e., the Neoceratopsia)9,21,39. However, a few studies suggest that the Psittacosauridae are more basal than the Chaoyangsauridae in ceratopsian phylogeny5,13,14. Our phylogenetic analysis incorporating new information from Mosaiceratops strongly supports the hypothesis that the Psittacosauridae are more derived than Chaoyangsauridae. The (Chaoyangsauridae + (Psittacosauridae + Neoceratopsia)) phylogenetic hypothesis is more consistent with the stratigraphic record than the (Psittacosauridae + (Chaoyangsauridae + Neoceratopsia)) hypothesis, indicated by our comparative stratigraphic fit indexes (MSM* values are 0.20 and 0.19 respectively). However, it should be pointed out that in general the calculated MSM* value is low and many relatively long ghost lineages exist (including the lineage leading to Mosaiceratops). Those significant gaps in the fossil record suggest great potential for future paleontological discoveries that hopefully will resolve some of the evolutionary history of the major features of the ceratopsian skeleton.
Additional Information
How to cite this article: Zheng, W. et al. A psittacosaurid-like basal neoceratopsian from the Upper Cretaceous of central China and its implications for basal ceratopsian evolution. Sci. Rep. 5, 14190; doi: 10.1038/srep14190 (2015).
References
Sereno, P. C. Phylogeny of the bird-hipped dinosaurs (Order Ornithischia). National Geographic Research 2, 234–256 (1986).
Makovicky, P. J. in The complete dinosaur (eds Michael K Brett-Surman, Thomas R Holtz, James O Farlow & Bob Walters ) Ch. 25, 527–550 (Indiana University Press, 2012).
Dodson, P. The horned dinoaurs. 346 (Princeton University Press, 1996).
Sereno, P. C. in The Dinosauria (eds D. B. Weishampel, P. Dodson & H. Osmolska ) 579–592 (University of California Press, 1990).
You, H.-L. & Dodson, P. in The Dinosauria (eds David B. Weishampel, Peter Dodson & Halszka Osmólska ) Ch. 22, 478–493 (The University of California Press, 2004).
Maryańska, T. & Osmólska, H. Protoceratopsidae (Dinosauria) of Asia. Palaeontologica Polonica 33, 133–181 (1975).
Coombs, W. P. J. Juvenile specimens of the ornithischian dinosaur Psittacosaurus mongoliensis. SO - Palaeontology (Oxford). 25(1). 1982, 89–108 (1982).
Osborn, H. F. Two Lower Cretaceous dinosaurs of Mongolia. American Museum Novitates 95, 1–10 (1923).
Xu, X., Forster, C. A., Clark, J. M. & Mo, J. A basal ceratopsian with transitional features from the Late Jurassic of northwestern China. Proceedings of the Royal Society B: Biological Sciences 273, 2135–2140, 10.1098/rspb.2006.3566 (2006).
Zhao, X.-J., Cheng, Z.-W. & Xu, X. The earliest ceratopsian from the Tuchengzi Formation of Liaoning, China. Journal of Vertebrate Paleontology 19, 681–691 (1999).
Zhao, X., Cheng, Z., Xu, X. & Makovicky, P. J. A new ceratopsian from the Upper Jurassic Houcheng Formation of Hebei, China. Acta Geologica Sinica - English Edition 80, 467–473, 10.1111/j.1755-6724.2006.tb00265.x (2006).
Xu, X., Makovicky, P. J., Wang, X.-l., Norell, M. A. & You, H.-l. A ceratopsian dinosaur from China and the early evolution of Ceratopsia. Nature 416, 314–317, 10.1038/416314a (2002).
Sereno, P. C. The origin and evolution of dinosaurs. Annual Review of Earth and Planetary Sciences 25, 435–489, 10.1146/annurev.earth.25.1.435 (1997).
Sereno, P. C. in The Age of Dinosaurs in Russia and Mongolia (eds Michael J. Benton, Mikhail A. Shishkin, David M. Unwin & Evgenii N. Kurochkin ) Ch. 25, 480–516 (Cambridge University Press, 2000).
Xu, X. et al. A new iguanodontian from Sangping Formation of Neixiang, Henan and its stratigraphical implication. Vertebrata PalAsiatica 38, 176–191 (2000).
Zhang, X. et al. A new sauropod dinosaur from the Late Cretaceous Gaogou Formation of Nanyang, Henan Province. Acta Geologica Sinica - English Edition 83, 212–221, 10.1111/j.1755-6724.2009.00032.x (2009).
Wang, D. et al. Discovery of invertebrate zoolite in the Xiaguan Formation of Xiaguan-Gaoqiu Basin, Henan, China and its importance for stratigraphic subdivision comparison. Acta Geologica Sinica 87, 1049–1058 (2013).
Zhou, S.-Q., Zhu, G.-B. & Feng, Z.-J. Xiaguan Formation and its era of Xiaguan-Gaogiu Basin, in Neixiang county, Henan Province. Resources Survey & Environment 24, 69–74 (2003).
Dong, Z. & Azuma, Y. in Sino-Japanese Silk Road Dinosaur Expedition (ed Zhiming Dong ) 68–89 (China Ocean Press, 1997).
You, H.-L. & Dodson, P. Redescription of neoceratopsian dinosaur Archaeoceratops and early evolution of Neoceratopsia. Acta Palaeontologica Polonica 48, 261–272 (2003).
Makovicky, P. J. & Norell, M. A. Yamaceratops Dorngobiensis, a new primitive ceratopsian (Dinosauria: Ornithischia) from the Cretaceous of Mongolia. American Museum Novitates. 1–42, 10.1206/0003-0082(2006)3530[1:YDANPC]2.0.CO;2 (2006).
You, H., Li, D., Lamanna, M. C. & Dodson, P. On a new genus of basal neoceratopsian dinosaur from the Early Cretaceous of Gansu Province, China. Acta Geologica Sinica 70, 593–597 (2005).
Makovicky, P. J. in Mesozoic Vertebrate Life (eds Darren H. Tanke & Kenneth Carpenter ) 243–262 (Indiana University Press, 2001).
Han, F. The Osteology of Yinlong downsi (Ornithischia: Ceratopsia) and the Phylogeny of the ornithischian dinosaurs Doctor of Science thesis, University of Chinese Academy of Sciences (2013).
Dodson, P., You, H.-L. & Tanoue, K. in New perspectives on horned dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium (eds Michael J. Ryan, Brenda J. Chinnery-Allgeier & David A. Eberth ) 221–233 (Indiana University Press, 2010).
You, H.-L., Tanoue, K. & Dodson, P. New data on cranial anatomy of the ceratopsian dinosaur Psittacosaurus major. Acta Palaeontologica Polonica 53, 183–196, 10.4202/app.2008.0202 (2008).
Sereno, P. C. & Chao, S. Psittacosaurus xinjiangensis (Ornithischia: Ceratopsia), a new psittacosaur from the Lower Cretaceous of northwestern China. Journal of Vertebrate Paleontology 8, 353–365, 10.1080/02724634.1988.10011724 (1988).
Sereno, P. C. in New Perspectives on Horned Dinosaurs (eds Michael J. Ryan, Brenda J. Chinnery-Allgeier & David A. Eberth ) Ch. 2, 21–58 (Indiana University Press, 2010).
Tanoue, K., You, H.-L. & Dodson, P. in New perspectives on horned dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium (eds Michael J. Ryan, Brenda J. Chinnery-Allgeier & David A. Eberth ) 234–250 (Indiana University Press, 2010).
You, H., Tanoue, K. & Dodson, P. A New Specimen of Liaoceratops yanzigouensis (Dinosauria: Neoceratopsia) from the Early Cretaceous of Liaoning Province, China. Acta Geologica Sinica - English Edition 81, 898–904, 10.1111/j.1755-6724.2007.tb01011.x (2007).
Morschhauser, E. M. The anatomy and phylogeny of Auroraceratops (Ornithischia: Ceratopsia) from the Yujingzi Basin of Gansu Province, China PhD thesis, University of Pennsylvania (2012).
Sereno, P. C., Zhao, X. & Tan, L. A new psittacosaur from Inner Mongolia and the parrot-like structure and function of the psittacosaur skull. Proceedings of the Royal Society B: Biological Sciences 277, 199–209, 10.1098/rspb.2009.0691 (2010).
Tanoue, K., You, H.-L. & Dodson, P. Comparative anatomy of selected basal ceratopsian dentitions. Canadian Journal of Earth Sciences 46, 425–439, 10.1139/E09-030 (2009).
Brown, B. Leptoceratops, a new genus of Ceratopsia from the Edmonton Cretaceous of Alberta. Bulletin American Museum of Natural History 33, 567–580 (1914).
Dodson, P., Forster, C. A. & Sampson, S. D. in The Dinosauria (eds David B. Weishampel, Peter Dodson & Halszka Osmólska ) Ch. 22, 494–513 (The University of California Press, 2004).
Brown, B. & Schlaikjer, E. M. The structure and relationships of Protoceratops. Annals of the New York Academy of Sciences 40, 133–266 (1940).
You, H.-L., Morschhauser, E. M., Dodson, P. & Li, D.-Q. Auroraceratops sp. (Dinosauria: Neoceratopsia) from the Early Cretaceous of the Mazongshan area in Northwestern China. Vertebrata PalAsiatica 50, 170–180 (2012).
Chinnery, B. J. & Horner, J. R. A new neoceratopsian dinosaur linking North American and Asian taxa. Journal of Vertebrate Paleontology 27, 625–641, 10.1671/0272-4634(2007)27[625:ANNDLN]2.0.CO;2 (2007).
Makovicky, P. J. in New perspectives on horned dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium (eds Michael J. Ryan, Brenda J. Chinnery-Allgeier & David A. Eberth ) 68–82 (Indiana University Press, 2010).
Xu, X., Wang, K., Zhao, X., Sullivan, C. & Chen, S. A new leptoceratopsid (Ornithischia: Ceratopsia) from the Upper Cretaceous of Shandong, China and its implications for neoceratopsian evolution. PLoS ONE 5, e13835, 10.1371/journal.pone.0013835 (2010).
Farke, A. A., Maxwell, W. D., Cifelli, R. L. & Wedel, M. J. A ceratopsian dinosaur from the Lower Cretaceous of Western North America and the Biogeography of Neoceratopsia. PLoS ONE 9, e112055, 10.1371/journal.pone.0112055 (2014).
Goloboff, P. A., Farris, J. S. & Nixon, K. C. TNT, a free program for phylogenetic analysis. Cladistics 24, 774–786, 10.1111/j.1096-0031.2008.00217.x (2008).
Siddall, M. E. Forum. stratigraphic fit to phylogenies: a proposed solution. Cladistics 14, 201–208, 10.1006/clad.1998.0059 (1998).
Pol, D. & Norell, M. A. Comments on the Manhattan Stratigraphic Measure. Cladistics 17, 285–289, 10.1006/clad.2001.0166 (2001).
Xu, X., Tang, Z.-L. & Wang, X.-L. A therizinosauroid dinosaur with integumentary structures from China. Nature 399, 350–354, 10.1038/20670 (1999).
Kohlsdorf, T. & Wagner, G. P. Evidence for the reversibility of digit loss: a phylogenetic study of limb evolution in Bachia (Gymnophthalmidae: Squamata). Evolution 60, 1896–1912, 10.1111/j.0014-3820.2006.tb00533.x (2006).
Wiens, J. J. Re-evolution of lost mandibular teeth in frogs after more than 200 million years and re-evaluating Dollo’s law. Evolution 65, 1283–1296, 10.1111/j.1558-5646.2011.01221.x (2011).
Cohen, K. M., Finney, S. C., Gibbard, P. L. & Fan, J.-X. The ICS International Chronostratigraphic Chart. Episodes 36, 199–204. (2013).
Acknowledgements
The support of the Zhejiang Museum of Natural History aided this project. We thank Yiming Sheng and Chaohe Yu for preparing the specimen, Ming Ding and Hao Wu provided access to the specimen, Corwin Sullivan and Eric Morschhauser provided helpful discussions, Fenglu Han aided with the use of TNT software and Ben Creisler helped with etymological consultations. Ashley Ferguson, Tom Stidham and Frankie Jackson gave useful comments on an earlier version of the manuscript. This project was supported by the National Natural Science Foundation of China (grant number: 41120124002). Free online versions of TNT and Mesquite were provided by the Willi Hennig Society and the Free Software Foundation Inc.
Author information
Authors and Affiliations
Contributions
X.X., W.Z. and X.J. designed the project. W.Z. and X.X. analyzed the data. W.Z. prepared the figures. W.Z., X.X. and X.J. wrote the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zheng, W., Jin, X. & Xu, X. A psittacosaurid-like basal neoceratopsian from the Upper Cretaceous of central China and its implications for basal ceratopsian evolution. Sci Rep 5, 14190 (2015). https://doi.org/10.1038/srep14190
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep14190
This article is cited by
-
A neoceratopsian dinosaur from the early Cretaceous of Mongolia and the early evolution of ceratopsia
Communications Biology (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
