
Abstract
In this paper we consider the problem of optimal search strategies on multi-linked networks, i.e. graphs whose nodes are endowed with several independent sets of links. We focus preliminarily on agents randomly hopping along the links of a graph, with the additional possibility of performing non-local hops to randomly chosen nodes with a given probability. We show that an optimal combination of the two jump rules exists that maximises the efficiency of target search, the optimum reflecting the topology of the network. We then generalize our results to multi-linked networks with an arbitrary number of mutually interfering link sets.
Similar content being viewed by others
Introduction
Complex networks are ubiquitous in nature and play a role of paramount importance in many contexts. Internet and the cyberworld, which permeate our everyday life, are self-organized hierarchical graphs. Urban traffic flows on intricate road networks, which impact both transportation design and epidemic control. In the brain, neurons are cabled through heterogeneous connections, which support the propagation of electric signals. In all these cases, the true challenge is to unveil the mechanisms through which specific dynamical features are modulated by the underlying topology of the network. Here, we study the utterly general problem of optimal search strategies on multi- linked networks, i.e. graphs which have independent sets of edges that connect the same group of nodes.
To this aim, let us start by considering a given agent (e.g. an electric pulse, an excitation, an animal or a human individual, such as a web surfer) located at a node of a network. The agent can hop to a neighbouring node, provided a link exists as specified by the adjacency matrix associated with the graph. The walker wanders on the network through a sequences of steps, that allow for a local exploration of its support. In such situations, the efficiency in reaching a specified location may be quantified by the mean first passage time, a robust and widely used measure of transport efficiency on networks in many contexts5,4, from biology19 and ecology8,10,40 to road network dynamics17 and quantum systems38,16.
However, local moves are not always the best option to reach a target efficiently. For example, facilitated diffusion in the cell nucleus, a mix of one-dimensional gliding along the DNA and three-dimensional jumps to adjacent DNA strands, is believed to account for the efficiency of transcription factors in finding their binding sites7,32,9. Analogously, inspired by the behaviour of foraging animals, it has been hypothesised that the local exploration of a connected territory might be complemented by intermittent relocation phases in order to optimize the searching strategy23. Accordingly, the animal would venture off-track through ballistic runs from time to time, thus sampling larger portions of space. In such examples, the relative duration of the local and relocation stages may control the optimization of the dual-stage strategy36.
Walkers on complex networks could in principle rely on similar integrated strategies, possibly tuned to the heterogeneous nature of the underlying support. In the seminal paper by Kleinberg25, the navigation on a regular two dimensional lattice distorted with the inclusion of long-range connections was studied and an optimum for the delivery time found, when tuning the probability of off-lattice connections between nodes. Exacts results and new insights on the navigability of small-world networks were later reported independently in Refs. 14 and 15. Since the pioneering contribution by Kleinberg, the problem of assisted exploration of a network has been addressed in a number of different contexts22,26,28,29,30,35,37, mainly relying on numerical investigations targeted to the problem at hand.
To help intuition let us consider, for example, web surfing. Starting the exploration from an arbitrary web page, one usually proceeds by following the hyperlinks which are therein available. This is a local search, which the user abandons when she opens a new tab to look for a different, potentially related topic, eventually landing into another virtual compartment which will be again probed locally for some time. On a different level, the brain displays multi-layered architectures of connections, that assist the finely orchestrated spatio-temporal patterns underlying brain function6. One may then speculate that electric signals can be transmitted across different layers, thus realizing de facto non-local jumps in the overall brain connectome between single-layer connected components. Non-local jumps (moves directed towards target sites which are not physically linked to the departure node) may be also loosely referred to as long-range, even though the underlying network is not necessarily endowed with a metric.
Building upon these ideas, we start by investigating the conditions for optimal target searches on a generic network of  nodes. In order to quantify search efficiency on a given network, we shall analytically compute mean first passage times31,34,24, which are widely used to gauge search strategies in many contexts42,2,21,46. To investigate the combined effect of local and non-local moves, we study a simple stochastic process which accommodates for both local diffusion and non-local relocation to sites which are not connected through the adjacency matrix.
nodes. In order to quantify search efficiency on a given network, we shall analytically compute mean first passage times31,34,24, which are widely used to gauge search strategies in many contexts42,2,21,46. To investigate the combined effect of local and non-local moves, we study a simple stochastic process which accommodates for both local diffusion and non-local relocation to sites which are not connected through the adjacency matrix.
An interesting application of such analysis is random walk on two networks that share the same nodes but are endowed with two different sets of links. Altogether, the two nested graphs sharing the same nodes might be termed a multi-linked network. Only local moves are allowed in this case, as prescribed by the two adjacency matrices associated with the two link sets. Moving from this observation, in the second part of the paper we will examine a more general setting where agents can diffuse in multi-linked networks endowed with an arbitrary number of independent link sets. The ensuing sub-networks can be imagined to be assembled as a stratified hierarchy of independent levels, each hosting an identical replica of the nodes, as in the spirit of a multiplex20,11,33,1. Indeed, it is straightforward to extend our analysis to the relevant and more general situation where only a subset of the nodes is present in each nested level.
Results
Relocation-assisted diffusion
Let  denote the
denote the  adjacency matrix of the network, with
adjacency matrix of the network, with  if
if  and
and  are physically connected by a link and
are physically connected by a link and  otherwise. The degree of node
otherwise. The degree of node  is given by
is given by  . The probability that a particle sitting at node
. The probability that a particle sitting at node  jumps on any other node
jumps on any other node  is specified by the following matrix
is specified by the following matrix

where  are the entries of a random symmetric sparse
are the entries of a random symmetric sparse  matrix, that controls the relocation via non-local hops (self-loops are, in principle, also allowed for). The density of ones in
matrix, that controls the relocation via non-local hops (self-loops are, in principle, also allowed for). The density of ones in  is measured by the parameter
is measured by the parameter  , so that the average number of nodes that can be reached from any node
, so that the average number of nodes that can be reached from any node  via off-network long-range jumps is
via off-network long-range jumps is  . The parameter
. The parameter  tunes the relative strength of the two competing mechanisms, local diffusion and random relocation. When
tunes the relative strength of the two competing mechanisms, local diffusion and random relocation. When  the walker explores the network according to a purely local rule, while in the opposite limit,
the walker explores the network according to a purely local rule, while in the opposite limit,  , hopping towards disconnected sites are the only allowed moves. For
, hopping towards disconnected sites are the only allowed moves. For  , the matrix
, the matrix  is filled with ones and
is filled with ones and  becomes the known Google matrix used in the PageRank Algorithm12,27.
becomes the known Google matrix used in the PageRank Algorithm12,27.
We define the search time as the time needed by a particle starting at node  to reach an absorbing trap located at node
to reach an absorbing trap located at node  . This satisfies the following relation (see Methods)
. This satisfies the following relation (see Methods)

where  . The subscript
. The subscript  indicates an
indicates an  submatrix obtained by suppressing the
submatrix obtained by suppressing the  -th row and the
-th row and the  -th column, while
-th column, while  denotes the identity matrix of size
denotes the identity matrix of size  . To assess the overall ability of the walker to find a target, we introduce a global parameter
. To assess the overall ability of the walker to find a target, we introduce a global parameter  by averaging Eq. (2) over all possible starting nodes (
by averaging Eq. (2) over all possible starting nodes ( ) and trap locations (
) and trap locations ( ), that is,
), that is,

In short,  quantifies the ability of the walker to search for targets at the global scale of the network. The shorter
quantifies the ability of the walker to search for targets at the global scale of the network. The shorter  , the more efficient the search. The quantity
, the more efficient the search. The quantity  acts as a free parameter – it can be adjusted to select the optimal balance between local and non-local hops, with the aim of minimizing the global exploration time.
acts as a free parameter – it can be adjusted to select the optimal balance between local and non-local hops, with the aim of minimizing the global exploration time.
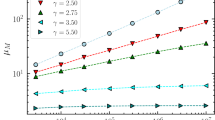
Figure 1 illustrates how  changes as a function of the relative weight of local and non-local moves for two different classes of synthetic undirected networks, the scale-free3,13 and the small-world44 networks. The curves display a clear minimum, implying the existence of an optimal value of
changes as a function of the relative weight of local and non-local moves for two different classes of synthetic undirected networks, the scale-free3,13 and the small-world44 networks. The curves display a clear minimum, implying the existence of an optimal value of  which minimizes the search time. Exactly the same behaviour is displayed by directed networks. The location of the minimum depends on the topology of the network, which defines the backbone for local diffusion, but also on the average number of sites that can be reached through a single long-range hop,
which minimizes the search time. Exactly the same behaviour is displayed by directed networks. The location of the minimum depends on the topology of the network, which defines the backbone for local diffusion, but also on the average number of sites that can be reached through a single long-range hop,  . Remarkably, the fewer sites are accessible through non-local jumps (i.e. the smaller
. Remarkably, the fewer sites are accessible through non-local jumps (i.e. the smaller  ), the more pronounced the optimality condition (see upper insets in Fig. 1).
), the more pronounced the optimality condition (see upper insets in Fig. 1).

The average search time on synthetic networks displays an optimum as a function of the relative weight of local and non-local moves.
Upper panel: scale-free network generated with the preferential attachment method3 with  and average connectivity
and average connectivity  . Lower panel: Watts and Strogatz small-world random network with
. Lower panel: Watts and Strogatz small-world random network with  44,
44,  and average connectivity
and average connectivity  . Here the sparse symmetric matrix
. Here the sparse symmetric matrix  has been generated with
has been generated with  (scale-free) and
(scale-free) and  (Watts-Strogatz). The insets show the position of the minimum
(Watts-Strogatz). The insets show the position of the minimum  and the corresponding shortest average time
and the corresponding shortest average time  (normalized to the case of a purely local walker,
(normalized to the case of a purely local walker,  ) as a function of the average fraction of long-range accessible nodes
) as a function of the average fraction of long-range accessible nodes  . The data are averaged over
. The data are averaged over  independent realizations of the random matrices
independent realizations of the random matrices  and error bars correspond to one standard deviation.
and error bars correspond to one standard deviation.
When  ,
,  approaches (but never reaches) the limiting solution
approaches (but never reaches) the limiting solution  . In this case, the walker can virtually land on any node with just one jump (the matrix
. In this case, the walker can virtually land on any node with just one jump (the matrix  is completely filled with ones) and local diffusion contributes modestly to further reduce the average searching time. Although a minimum always exists also for
is completely filled with ones) and local diffusion contributes modestly to further reduce the average searching time. Although a minimum always exists also for  (the Google Matrix case),
(the Google Matrix case),  is very close to
is very close to  , the time the walker needs to reach an isolated trap when
, the time the walker needs to reach an isolated trap when  is exactly set to zero. Conversely, when
is exactly set to zero. Conversely, when  , non-local short-cuts are only available towards a subset of nodes. This is a more plausible situation, bearing in mind the afore-mentioned applications. When surfing the web, from time to time one will abandon a given area of exploration to look for the presumed central node of a new region that she wishes to sample. Similarly, long-range connections in the brain, established through trans-layer channels, are certainly fewer than those accounting for effective bridges among the
, non-local short-cuts are only available towards a subset of nodes. This is a more plausible situation, bearing in mind the afore-mentioned applications. When surfing the web, from time to time one will abandon a given area of exploration to look for the presumed central node of a new region that she wishes to sample. Similarly, long-range connections in the brain, established through trans-layer channels, are certainly fewer than those accounting for effective bridges among the  nodes of a given layer.
nodes of a given layer.
The relocation-assisted search is 10–15% more efficient with respect to the purely local dynamics for intermediate values of the density  of available distant nodes (insets in Fig. 1). The same analysis performed with different values of the average connectivity
of available distant nodes (insets in Fig. 1). The same analysis performed with different values of the average connectivity  (scale-free network) and of the rewiring parameter p (Watts-Strogatz) yields similar results. In particular, upon decreasing
(scale-free network) and of the rewiring parameter p (Watts-Strogatz) yields similar results. In particular, upon decreasing  one recovers the same qualitative behaviour as obtained when increasing
one recovers the same qualitative behaviour as obtained when increasing  (data not shown). In this paper, we have chosen to focus on small networks because of the computational cost required to invert the matrix Zj, for any given j. However, the method is utterly general and may be applied to systems of arbitrary size N. In the Supplementary Information, we show that
(data not shown). In this paper, we have chosen to focus on small networks because of the computational cost required to invert the matrix Zj, for any given j. However, the method is utterly general and may be applied to systems of arbitrary size N. In the Supplementary Information, we show that  progressively decreases as N increases, while keeping
progressively decreases as N increases, while keeping  constant. Conversely, working at fixed size N one can change the position of the minimum as sought, by acting on
constant. Conversely, working at fixed size N one can change the position of the minimum as sought, by acting on  . The counter-intuitive conclusion is that the density of non-local links should be reduced as N gets larger to keep the optimum value of
. The counter-intuitive conclusion is that the density of non-local links should be reduced as N gets larger to keep the optimum value of  fixed. Interestingly, the advantage brought about by the assisted search over purely local diffusion is preserved if N and
fixed. Interestingly, the advantage brought about by the assisted search over purely local diffusion is preserved if N and  are simultaneously tuned so to keep
are simultaneously tuned so to keep  fixed. The data reported in the Supplementary Information refer to the Watts-Strogatz setting but the same conclusions hold in general. As a side remark, we recall that the searching time can be expressed as a function of the nodes connectivity, for specific classes of networks, in the large N limit43. Such large networks could share the same characteristics of searchability for the limiting choice
fixed. The data reported in the Supplementary Information refer to the Watts-Strogatz setting but the same conclusions hold in general. As a side remark, we recall that the searching time can be expressed as a function of the nodes connectivity, for specific classes of networks, in the large N limit43. Such large networks could share the same characteristics of searchability for the limiting choice  . When introducing the effect of long-range teleportation one can in principle remove such degeneracy and obtain distinct optimal values of
. When introducing the effect of long-range teleportation one can in principle remove such degeneracy and obtain distinct optimal values of  .
.
To confirm the existence of an optimal searching strategy on real data sets, we have extracted the adjacency graph of small portions of the web, starting from the home-pages of four main European newspapers (see Methods). The top panel of Fig. 2 shows that the general picture described above for synthetic data sets is valid for real networks too. This has nothing to do with the peculiar structure of the Web, for the same analysis performed on small neural networks of different animals confirms the existence of a clear minimum in the average search time (bottom panel in Fig. 2).

The average search time on real data sets displays an optimum as a function of the relative weight of local and non-local moves.
Top: average time  as a function of
as a function of  for four real Web subgraphs. The
for four real Web subgraphs. The  adjacency matrices have been mapped out by a Web crawler starting from the web sites of four major European newspapers (see Methods). The sparse symmetric matrix
adjacency matrices have been mapped out by a Web crawler starting from the web sites of four major European newspapers (see Methods). The sparse symmetric matrix  has been generated with
has been generated with  . Bottom: search time in two neuronal networks: cortical connectivity network of cats (
. Bottom: search time in two neuronal networks: cortical connectivity network of cats ( nodes, left39) and macaques (
nodes, left39) and macaques ( nodes, right45). In both cases we have used
nodes, right45). In both cases we have used  .
.
In all the cases examined,  appears to be a convex smooth function of
appears to be a convex smooth function of  with a clear minimum. One may ask whether this is a widespread feature of many graphs. More generally, it would be helpful to work out a quantitative criterion to predict whether an optimal search strategy exists at all, possibly also identifying the optimal balance between local and non-local moves required to place oneself in such regime. Unfortunately, the exact dependence of
with a clear minimum. One may ask whether this is a widespread feature of many graphs. More generally, it would be helpful to work out a quantitative criterion to predict whether an optimal search strategy exists at all, possibly also identifying the optimal balance between local and non-local moves required to place oneself in such regime. Unfortunately, the exact dependence of  on
on  is hidden in the inverse of the matrix
is hidden in the inverse of the matrix  which, in general, cannot be computed explicitly. However, a principle of this sort can be formulated by resorting to a perturbative approach. If we assume that the stationary point is located at sufficiently small values of
which, in general, cannot be computed explicitly. However, a principle of this sort can be formulated by resorting to a perturbative approach. If we assume that the stationary point is located at sufficiently small values of  , then we may consider a search time of the form
, then we may consider a search time of the form

In this case, the coefficients  ,
,  and
and  , which depend on the topology of the network, can be computed analytically (see Methods). A necessary and sufficient condition for a meaningful minimum to occur is
, which depend on the topology of the network, can be computed analytically (see Methods). A necessary and sufficient condition for a meaningful minimum to occur is  and
and  with
with  , which ensures that
, which ensures that  . This provides a handy rule to enquire about the existence of an optimality condition for any given network.
. This provides a handy rule to enquire about the existence of an optimality condition for any given network.
In all the cases that we examined, the coefficients  turn out to be positive. Therefore a minimum is always predicted to exist under the quadratic approximation and closed expressions for both
turn out to be positive. Therefore a minimum is always predicted to exist under the quadratic approximation and closed expressions for both  and
and  can be readily obtained. These match well the exact data computed through Eq. (3). The agreement is of course better when the minimum is found close to
can be readily obtained. These match well the exact data computed through Eq. (3). The agreement is of course better when the minimum is found close to  (Fig. 2–4). Explicit analytical expressions can be obtained in some limiting cases. When
(Fig. 2–4). Explicit analytical expressions can be obtained in some limiting cases. When  one recovers the Google matrix and the transition rate from node
one recovers the Google matrix and the transition rate from node  to node
to node  reads
reads  . In this case it is not difficult to show that
. In this case it is not difficult to show that  ,
,  and
and  , where
, where  (see Supplementary Information for the full derivation). In the case of a regular lattice of connectivity
(see Supplementary Information for the full derivation). In the case of a regular lattice of connectivity  , one immediately finds
, one immediately finds  and
and  . The more links per node are added, the larger the value of
. The more links per node are added, the larger the value of  (
( ) and the deeper the minimum for
) and the deeper the minimum for  vs
vs  . Although
. Although  is shorter than the search time
is shorter than the search time  obtained for
obtained for  , the associated correction is just
, the associated correction is just  .
.

Formula (4) provides a convenient tool to enquire about the existence of an optimality criterion for the search time in a given network.
The average search time in a small random network of  nodes (symbols) computed from Eq. (3) is compared to the approximated quadratic profile described by Eq. (4) (solid line). The inset shows a close-up of the region around the minimum. Other parameters are:
nodes (symbols) computed from Eq. (3) is compared to the approximated quadratic profile described by Eq. (4) (solid line). The inset shows a close-up of the region around the minimum. Other parameters are:  ,
,  .
.

Existence of an optimum for the average search time in a multi-linked network with  .
.
Density plot of the average search time for networks with  nodes and
nodes and  . Data are obtained from formula (3), via numerical inversion of matrix (5), for every choice of
. Data are obtained from formula (3), via numerical inversion of matrix (5), for every choice of  and
and  . In both cases here depicted, the three matrices are generated according to the Watts-Strogatz algorithm, for different choices of the probability
. In both cases here depicted, the three matrices are generated according to the Watts-Strogatz algorithm, for different choices of the probability  and the initial connectivity
and the initial connectivity  , (
, ( ):
):  ,
,  ,
,  ,
,  (lower triangle),
(lower triangle),  ,
,  ,
,  ,
,  (upper triangle). The dashed crosses identify the optimal values for
(upper triangle). The dashed crosses identify the optimal values for  and
and  as predicted by the approximated numerical expression (8).
as predicted by the approximated numerical expression (8).
As remarked at the beginning, the analysis carried out above can be interpreted as diffusion on graphs characterized by a single set of nodes and two independent sets of links, represented by two different adjacency matrices  and
and  . This kind of network could be termed double-linked. Building on this observation, we discuss in the following the generalization of our approach to arbitrary multi-linked networks, where an arbitrary number of independent link sets is imposed upon the same ensemble of nodes.
. This kind of network could be termed double-linked. Building on this observation, we discuss in the following the generalization of our approach to arbitrary multi-linked networks, where an arbitrary number of independent link sets is imposed upon the same ensemble of nodes.
The general case of M networks
Let us consider agents hopping on a given set of nodes, that are joined together through M independent sets of links, represented by as many different adjacency matrices. We term such a structure a multi-linked network, i.e. a graph where links attached to a given node have an identity. For example, in a transportation network two nodes (cities) could be linked through a plane connection (air link-set) and by train (railway link-set). With reference to this setting, we set out to derive the optimal coupling that maximizes the transport efficiency across the system. Again with reference to mobility applications, one can consider the effective design of an integrated urban transportation network18. Underground, cars and bus service define three different networks (three different link systems on the same set of nodes): their relative load in terms of users and a smart planning of the respective infrastructures could be studied via a systemic approach that builds on the formalism developed here.
In a multi-linked graph the probability of transition is specified by the following stochastic matrix

where Al are M matrices of size  and Kl are their associated diagonal matrices of connectivities, namely
and Kl are their associated diagonal matrices of connectivities, namely  . In this case the reduced matrix Zj reads:
. In this case the reduced matrix Zj reads:

where j flags the position of the absorbing trap. The time it takes to reach j starting from  can be computed again by means of expression (2) and the average trapping time
can be computed again by means of expression (2) and the average trapping time  estimated through eq. (3). Following a straightforward generalization of the calculation carried out in the preceding paragraph, one can derive an approximate expression for
estimated through eq. (3). Following a straightforward generalization of the calculation carried out in the preceding paragraph, one can derive an approximate expression for  (see Methods)
(see Methods)

To compute the stationary point of  , one needs to impose the condition
, one needs to impose the condition  , which is equivalent to setting
, which is equivalent to setting

The stationary point of (6) is a global minimum if the associated Hessian matrix is positive defined.
To clarify the interest of the proposed generalization, we fix  and investigate the average trapping time as a function of the two independent parameters
and investigate the average trapping time as a function of the two independent parameters  and
and  . In Fig. 4 we show a density map in the
. In Fig. 4 we show a density map in the  ,
,  plane of the exact search time
plane of the exact search time  computed for two different multi-linked networks following the prescription (3). A minimum is clearly found in both cases, signaling the existence of an optimal choice of the coupling strengths that maximizes the transport efficiency. In the context of the above example, this knowledge could be exploited for the design of efficiently integrated facilities in urban transportation planning, or to favour a smart distribution of users through existing infrastructures.
computed for two different multi-linked networks following the prescription (3). A minimum is clearly found in both cases, signaling the existence of an optimal choice of the coupling strengths that maximizes the transport efficiency. In the context of the above example, this knowledge could be exploited for the design of efficiently integrated facilities in urban transportation planning, or to favour a smart distribution of users through existing infrastructures.
Approximate analytical expressions for the optimal values of  and
and  can be readily obtained from system (7), which gives
can be readily obtained from system (7), which gives

The white crosses in Fig. 4 refer to the above solution. As it happens in the case of  analyzed in the previous section, the approximate solution is a better estimate of the true minimum when this is found close to
analyzed in the previous section, the approximate solution is a better estimate of the true minimum when this is found close to  .
.
Discussion
Summarising, in this paper we have addressed the problem of search on networks. To this end, we have preliminarily studied the trapping problem for a modified random walk that combines local hops along the links of the graph with non-local relocation jumps, i.e. not along the links contained in the adjacency matrix. We have shown, both for artificial and real datasets, that an optimal balance between local and non-local moves exists which minimizes the average time required to reach a trap. Furthermore, we have derived closed analytical expressions, that enable one to predict the optimal combination as a function of the network topology.
In the second part of the paper we have focused on a random walker diffusing on a arbitrary number of interconnected networks, which share the same set of nodes but are characterized by different adjacency matrices. We have termed a complex structure of this kind a multi-linked network. Also in this generalized setting an optimum for the average transport time is found, which can be controlled by tuning the different coupling strengths among the individual sub-networks. In conclusion, the optimality criterion seems to be a universal dynamical mechanism, which might have exerted a critical pressure in the evolutionary selection of many naturally occurring network architectures of the multi-linked kind and that might equally well be exploited in the optimization of man-made technological solutions.
Methods
Computation of first passage times
The mean first passage time  , namely the time it takes for a walker starting at site
, namely the time it takes for a walker starting at site  to get to any one of
to get to any one of  randomly placed traps, can be computed by extending to the case of a network the standard argument used in the continuum limit for a random walk on a line. Let us consider the interval
randomly placed traps, can be computed by extending to the case of a network the standard argument used in the continuum limit for a random walk on a line. Let us consider the interval  on the real axis and a random walk with two absorbing boundaries located at
on the real axis and a random walk with two absorbing boundaries located at  and
and  . The time interval between two jumps is
. The time interval between two jumps is  and the lattice spacing is
and the lattice spacing is  . The exit time
. The exit time  obeys to
obeys to  meaning that the walker can be regarded as starting one step in the future with equal probability from either
meaning that the walker can be regarded as starting one step in the future with equal probability from either  or
or  . The generalization of this equation for a random walk on a network is simply given by
. The generalization of this equation for a random walk on a network is simply given by  , a formulation which proves particularly convenient to investigate the trapping problem. Indeed, re-labelling the nodes of the network so as to have non-trap nodes going from
, a formulation which proves particularly convenient to investigate the trapping problem. Indeed, re-labelling the nodes of the network so as to have non-trap nodes going from  to
to  and all traps located at nodes
and all traps located at nodes  to
to  , one obtains a matrix
, one obtains a matrix  with the last
with the last  rows equal to zero. Rearranging correspondingly the array
rows equal to zero. Rearranging correspondingly the array  and recalling that
and recalling that  for
for  , one finds that the exit times are solution of the linear system
, one finds that the exit times are solution of the linear system  , where we have denoted by
, where we have denoted by  the upper-left
the upper-left  block square sub-matrix of
block square sub-matrix of  . Eq. (2) is the formal solution of this last equation.
. Eq. (2) is the formal solution of this last equation.
Adjacency matrices of sub-networks from the web
To gather real data from the Web we have used the Web crawler surfer.m (http://www.mathworks.com/). Starting from a selected URL, the crawler identifies all the hyperlinks in the page and adds them to the list of URLs to visit. Once all these URLs are visited, the procedure is repeated recursively for each URL in the list until the assigned number of websites is reached. The outcome of the algorithm is stored in an adjacency matrix where nodes represent the visited pages: the entries of the matrix are  if two pages are connected trough a hyperlink,
if two pages are connected trough a hyperlink,  otherwise. The matrix is then symmetrized.
otherwise. The matrix is then symmetrized.
Perturbative expansion of a sum of two matrices
Let  and
and  be two arbitrary non-singular square matrices of the same dimension and let us introduce the operator
be two arbitrary non-singular square matrices of the same dimension and let us introduce the operator  , that returns the sum of all the elements of a given square matrix. Starting from the relation
, that returns the sum of all the elements of a given square matrix. Starting from the relation  and expressing
and expressing  by a Neumann series41, it follows
by a Neumann series41, it follows  . To apply this approximation to Eq. (3), we introduce two diagonal matrices associated with
. To apply this approximation to Eq. (3), we introduce two diagonal matrices associated with  and
and  , namely
, namely  and
and  . In this way,
. In this way,  takes the form
takes the form  . Consequently, by denoting again by
. Consequently, by denoting again by  the position of the trap, the terms of the reduced matrix
the position of the trap, the terms of the reduced matrix  can be easily rearranged by collecting together those proportional to
can be easily rearranged by collecting together those proportional to  . In formulae:
. In formulae:  . Setting
. Setting  ,
,  and
and  and applying the operator
and applying the operator  to
to  , we recover Eq. (4) with
, we recover Eq. (4) with  ,
,  and
and  .
.
Generalization to the case of  matrices
matrices
 matrices
matricesFollowing the same perturbative scheme detailed above, one can expand the inverse of  defined in eq. (5) as:
defined in eq. (5) as:  .
.
Applying the operator  and summing over all trap locations, one obtains expression (6) for
and summing over all trap locations, one obtains expression (6) for  , where:
, where:  ,
,  ,
,  and
and  .
.
References
Asllani, M., Busiello, D., Carletti, T., Fanelli, D. & Planchon, G. Turing patterns in multiplex networks. Phys. Rev. E 90, 042814 (2014).
Agliari, E. & Burioni, R. Random walks on deterministic scale-free networks: Exact results. Phys. Rev. E 80, 031125 (2009).
Barabási, A. L. & Albert, R. Emergence of scaling in random networks. Science 286, 509-512 (1999).
Baronchelli, A. & Loreto, V. Ring structures and mean first passage time in networks. Phys. Rev. E 73, 026103 (2006).
Barrat, A., Barthélemy, M. & Vespignani, A. Dynamical processes in complex networks. (Cambridge University Press, New-York., 2008).
Bassett, D. S. & Lynall, M. E. [Network methods to characterize brain structure and function] Cognitive Neurosciences: the Biology of the Mind [Gazzaniga, G. R. M. M. & Ivry, R. B. (ed.)] (W. W. Norton., 2014).
Bauer, M. & Metzler, R. Generalized facilitated diffusion model for dna-binding proteins with search and recognition states. Biophys. J. 102, 2321–2330 (2014).
Bénichou, O., Coppey, M., Moreau, M., Suet, P. H. & Voituriez, R. Optimal search strategies for hidden targets. Phys. Rev. Lett. 94, 198101 (2005).
Bénichou, O., Chevalier, C., Meyer, B. & Voituriez, R. Facilitated Diffusion of Proteins on Chromatin. Phys. Rev. Lett. 106, 038102 (2011).
Bénichou, O., Loverdo, C., Moreau, M. & Voituriez, R. Intermittent search strategies. Rev. Mod. Phys. 83, 81–129 (2011).
Bianconi, G. Statistical mechanics of multiplex networks: Entropy and overlap Phys. Rev. E 87, 062806 (2013).
Brin, S. & Page, L. [The anatomy of a large-scale hypertextual web search engine] Seventh International World-Wide Web Conference (WWW 1998). (1998).
Caldarelli, G. Scale-Free Networks: Complex Webs in Nature and Technology (Oxford University Press., 2007).
Caretta Cartozo, C. and De Los Rios, P. Extended Navigability of Small World Networks: Exact Results and New Insights. Phys. Rev. Lett. 102, 238703 (2009).
Carmi, S., Carter, S., Sun, J. and ben-Avraham, D. Asymptotic Behavior of the Kleinberg Model. Phys. Rev. Lett. 102, 238702 (2009).
Caruso, F., Huelga, S. F. & Plenio, M. B. Noise-enhanced classical and quantum capacities in communication networks. Phys. Rev. Lett. 105, 90501 (2010).
Crisostomi, E., Kirkland, S. & Shorten, R. A google-like model of road network dynamics and its application to regulation and control. Int. J. Control 84, 633–651 (2011).
De Domenico, M., Solé-Ribalta, A., Gómez, S. & Arenas, A. Navigability of interconnected networks under random failures. Proc. Natl. Acad. Sci. U.S.A. 111, 8351–8356 (2014).
Gallos, L. K., Song, C., Havlin, S. & Makse, H. A. Scaling theory of transport in complex biological networks. Proc. Natl. Acad. Sci. U.S.A. 104, 7746–7751 (2007).
Gómez-Gardeñes, J., Reinares, I., Arenas, A. & Floria, L. M. Evolution of cooperation in multiplex networks Sci. Rep. 2, 620 (2012).
Haynes, C. P & Roberts, A. P. Global first-passage times of fractal lattices. Phys. Rev. E 78, 041111 (2008).
Lee, S. H., Holme P. . Exploring Maps with Greedy Navigators Phys. Rev. Lett. 108, 128701 (2012).
JohnO’Brien, W., Evans, B. & Browman, H. Flexible search tactics and efficient foraging in saltatory searching animals. Oecologia 80, 100–110 (1989).
Kittas, A., Carmi, S., Havlin, S. & Argyrakis, P. Trapping in complex networks. Europhys. Lett. 84, 40008 (2008).
Kleinberg, J. M. Navigation in a small world. Nature 406, 854 (2000).
Komidis, K., Havlin, S. & Bunde, A. Structural properties of spatially embedded networks. Europhys. Lett. 82, 48005 (2008).
Langville, A. M. & Meyer, C. D. Google’s PageRank and Beyond: The Science of Search Engine Rankings (Princeton University Press, Princeton., 2006).
Lee, S. H. & Holme, P. Exploring Maps with Greedy Navigators. Phys. Rev. Lett. 108, 128701 (2012).
Li, G. et al. Towards Design Principles for Optimal Transport Networks. Phys. Rev. Lett. 104, 018701 (2010).
Li, G. et al. Optimal transport exponent in spatially embedded networks. Phys. Rev. E 87, 042810 (2013).
Lin, Y. & Zhang, Z. Mean first-passage time for maximal-entropy random walks in complex networks. Sci. Rep. 4, 06 (2014).
Mirny, L. et al. How a protein searches for its site on DNA: the mechanism of facilitated diffusion. J. Phys. A: Math. Theor. 42, 434013 (2009).
Nicosia, V., Bianconi, G., Latora, V. & Barthelemy M. . Growing Multiplex Networks. Phys. Rev. Lett. 111, 058701 (2013).
Noh, J. D. & Rieger, H. Random walks on complex networks. Phys. Rev. Lett. 92, 118701 (2004).
Oliveira, C. L. N., Morais, P. A., Moreira, A. A & Andrade, J. S. Enhanced Flow in Small-World Networks. Phys. Rev. Lett. 112, 148701 (2014).
Oshanin, G., Lindenberg, K., Wio, H. S. & Burlatsky, S. Efficient search by optimized intermittent random walks. J. Phys. A: Math. Theor. 42, 434008 (2009).
Ramezanpour, A. Intermittent exploration on a scale-free network. Europhys. Lett., 77, 60004 (2007).
Sánchez-Burillo, E., Duch, J., Gómez-Gardeñes, J. & Zueco, D. Quantum navigation and ranking in complex networks. Sci. Rep. 2, 08 (2012).
Scannell, J. W., Burns, G. A. P. C., Hilgetag, C. C., O’Neil, M. A & Young, M. P. The connectional organization of the cortico-thalamic system of the cat. Cerebral Cortex 9, 277–299 (1999).
Shlesinger, M. F. Mathematical physics: Search research. Nature 443, 281–282 (2006).
Stewart, G. W. [Matrix Algorithms: Volume 1: Basic Decompositions] Other Titles in Applied Mathematics (SIAM., 1998).
Tejedor, V. B. é. n. i. c. h. o. u., O. & Voituriez, R. Global mean first-passage times of random walks on complex networks. Phys. Rev. E 80, 065104 (2009).
von Luxburg U., Radl A. & Hein M., Hitting and Commute times in Large Random Neighbourhood Graphs, JMLR15, 1751–1798 (2014).
Watts, D. J. & Strogatz, S. H. Collective dynamics of small-world networks. Nature 393, 440–442 (1998).
Young, M. P. The organization of neural systems in the primate cerebral cortex. Proc. Biol. Sci. 252, 13–18 (1993).
Zhang, Z., Lin, Y. & Ma, Y. Effect of trap position on the efficiency of trapping in treelike scale-free networks. J. Phys. A: Math. Theor., 44, 075102 (2011).
Acknowledgements
The authors would like to thank Levon Chakhmakhchyan and Filippo Caruso for inspiring discussions and Alessio Cardillo for his critical reading of the manuscript. This work has been partially supported by Ente Cassa di Risparmio di Firenze and program PRIN 2012 founded by the Italian Ministero dell’Istruzione, dell’Università e della Ricerca (MIUR).
Author information
Authors and Affiliations
Contributions
F. D. P., D.F. and F. P designed the study and carried out the analysis. F. D. P. carried out the numerical calculations. All authors contributed to the writing of the manuscript.
Supplementary Information accompanies this paper at http://www.nature.com/scientificreports
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Di Patti, F., Fanelli, D. & Piazza, F. Optimal search strategies on complex multi-linked networks. Sci Rep 5, 9869 (2015). https://doi.org/10.1038/srep09869
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep09869
This article is cited by
-
Navigation by anomalous random walks on complex networks
Scientific Reports (2016)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
