
Abstract
Pesticide residues in agricultural produce pose a threat to human health worldwide. Although the detoxification mechanisms for xenobiotics have been extensively studied in mammalian cells, information about the regulation network in plants remains elusive. Here we show that brassinosteroids (BRs), a class of natural plant hormones, decreased residues of common organophosphorus, organochlorine and carbamate pesticides by 30–70% on tomato, rice, tea, broccoli, cucumber, strawberry and other plants when treated externally. Genome-wide microarray analysis showed that fungicide chlorothalonil (CHT) and BR co-upregulated 301 genes, including a set of detoxifying genes encoding cytochrome P450, oxidoreductase, hydrolase and transferase in tomato plants. The level of BRs was closely related to the respiratory burst oxidase 1 (RBOH1)-encoded NADPH oxides-dependent H2O2 production, glutathione biosynthesis and the redox homeostasis and the activity of glutathione S-transferase (GST). Gene silencing treatments showed that BRs decreased pesticide residues in plants likely by promoting their metabolism through a signaling pathway involving BRs-induced H2O2 production and cellular redox change. Our study provided a novel approach for minimizing pesticide residues in crops by exploiting plants' own detoxification mechanisms.
Similar content being viewed by others


Introduction
Pesticides are important chemicals used in modern agriculture for protection against pests and weeds, as up to 80% of crop yields could be lost without pesticides1. The extensive use of pesticides has resulted in contamination of produce and the environment in most countries. This has become a major food safety issue because many pesticides have not only acute toxicity, but also sublethal effects by disrupting the endocrine balance and affecting sperm quality and reproductive development2,3,4,5,6,7. In mammalian cells, the detoxification process can be divided into several phases, including oxidation by cytochrome P450s or peroxidases (phase I), conjugation with glutathione and glucose through the action of glutathione S-transferases (GST) or glycosyltransferase (phase II) and extrusion of the xenobiotic conjugates and metabolites from the cell by specific membrane-associated transporters such as ABC transporters (phase III)8,9,10,11. In plants, the expression of specific genes involved in the processes related to detoxification is responsive to xenobiotic compounds such as pesticides12, suggesting that the intrinsic detoxification mechanisms of higher plants may be exploited to decrease pesticide residues in food produce. Brassinosteroids (BRs) are a class of phytohormones involved in the regulation of plant growth, development and stress response13, Previously, we found that BRs-induced stress tolerance is associated with changes in cellular redox homeostasis and expression of a wide range of stress-related genes including those encoding P450 and GST involved in metabolism of xenobiotic compounds14,15. Here, we show that BRs could enhance pesticide degradation most likely by enhancing glutathione metabolism and glutathione S-transferase (GST) activity via a Respiratory burst oxidase homologue1 (RBOH1)-dependent pathway. Our study revealed yet another potentially important application of these plant hormones for improving the food safety of agricultural produce.
Results
Involvement of BR signaling in the regulation of chlorothalonil (CHT) degradation in various plants
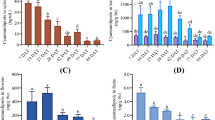
Chlorothalonil (CHT) is among the most widely used pesticides in crop production. We first compared the effects of different concentrations of 24-epibrassinolide (EBR), an active brassinosteroid, from levels within physiological range to saturation level on CHT residues in tomato leaves. Time course of CHT residue revealed that CHT residue rarely decreased after 6 d of CHT application (Supplementary Fig. S1). EBR at the low concentration of 0.02 μM dramatically decreased the residue by 38.9% (Fig. 1a). Increasing the concentration of EBR, however, did not further accelerate CHT degradation (Fig. 1a; Supplementary Fig. S1). This result indicated that EBR concentration within physiological ranges is sufficient to upregulate the machinery of pesticide metabolism.

(a) Chlorothalonil (CHT) residues in tomato leaves as influenced by the application dose of 24-epibrassinolide (EBR). Plants at the 6-leaf stage were sprayed with EBR at different concentrations and then exposed to CHT at 11.2 mM. Leaves were taken at 5 d after application of CHT. (b) Effects of EBR application on the pesticide residues in various food crops. EBR at 0.1 μM was applied 6 h prior to pesticide application and pesticide residues were determined 7 d afterward. Data are means of four biological replicates (±SD).
To determine whether BRs are general activators of pesticide metabolism in plants, we further tested with other crop plants and a variety of pesticides. EBR at 0.1 μM was co-treated with chlorpyrifos, phoxim, chlorothalonil, omethoate, cypermethrin, carbofuran or 3-hydroxycarbofuran at commercially recommended doses on rice, tea, cucumber, broccoli, asparagus, strawberry, celery, garlic, Chinese chives and Chinese cabbage plants. As found in tomato, there were significant decreases of pesticide residues in all of these crops, by an average of 34.0–71.3%, during the first 7 d after pesticide application (Fig. 1b). These results indicate that BRs are generally effective in enhancing the degradation of various pesticides in higher plants of agricultural importance.
We next examined whether endogenous BR biosynthesis and signaling affect pesticide metabolism in tomato plants by comparing between BR-deficient mutant dim and its corresponding wild-type (WT), or between the BR receptor BRI1-silenced (pTRV-BRI1) and nonsilenced (pTRV) plants for the levels of CHT residues with or without EBR application. As shown in Fig. 2a, the levels of CHT residues in the dim and the pTRV-BRI1 plants were 21.7% and 30.2% higher than those in the WT plants and the non-silenced (pTRV) plants, respectively. Pretreatment with EBR decreased the CHT residues in the WT plants and dim plants by 31.6% and 58.1%, respectively (Fig. 2a). In contrast, silencing of BRI1 (pTRV-BRI1) abolished the EBR-induced reduction in CHT residues in plants (Fig. 2b). These results strongly suggest that BRs are involved in the regulation of CHT metabolism in tomato plants.

(a) Chlorothalonil (CHT) residue in the wild-type (WT) and BR-deficient mutant dim tomato plants as influenced by the application of 24-epibrassinolide (EBR). (b) CHT residues in the control (pTRV) and BRs receptor gene BRI1-silenced (pTRV-BRI1) plants as influenced by the application of EBR. EBR at 0.1 μM was applied 6 h prior to pesticide application and leaves were taken at 5 d after application of CHT at 11.2 mM. Data are means of four biological replicates (±SD). Means denoted by the same letter did not significantly differ at P < 0.05 according to Tukey's test.
Plant responses to pesticide and brassinosteroids might be regulated by common signaling transduction pathway
The signaling pathways associated with the response to xenobiotic compounds in mammalian cells involve both xenobiotic ligand-activated transcription factors and redox sensing proteins10,11. In plants, however, no signaling components involved in plant responses to xenobiotic compounds such as pesticides have been identified. To determine the molecular responses of plant to pesticides and EBR, Affymetrix GeneChip Tomato Genome Array with more than 9,200 gene-specific probes was used to identify differentially expressed genes. A total of 1584, 1545 and 1725 genes were differentially expressed in EBR-, CHT- and EBR+CHT-treated plants, respectively. Among these differentially regulated genes, 544, 670 and 700 genes were up-regulated by EBR, CHT and EBR+CHT, respectively (Fig. 3a). Of striking interest is that 301 genes were commonly upregulated by EBR, CHT and EBR+CHT treatments and many of these genes encode proteins belonging to the catalytic category (Supplementary Table S1). In addition to those encoding cytochrome P450, GST, hydrolase and oxidoreductase, there were many upregulated genes encoding putative transcription factors or proteins associated with signal transduction, including ethylene response factor, WRKY factors, NADPH oxidase and protein kinases. Analysis of qRT-PCR further showed that both EBR and CHT differentially induced the transcripts of RBOH1 (Respiratory burst oxidase homologue1, encoding NADPH oxidase), the cytochrome P450 encoding genes (accessions AI776109 and BF112381), GSH1-2 (encoding γ-glutamyl cysteine synthetase and glutathione synthetase, respectively), GR (encoding glutathione reductase), GST1-7 and ABC1-4 (encoding ABC transporters) (Fig. 3b; Supplementary Fig. S2). Furthermore, the transcripts for most of these genes were more strongly induced by the combined EBR+CHT treatment than by the EBR or CHT alone treatment, suggesting an additive effect of BR and CHT on the induction of detoxifying mechanisms. Consistent with gene transcripts, accumulation of H2O2 and glutathione pool (reduced glutathione GSH and oxidized glutathione GSSG) and GST activity were induced by EBR and CHT (Fig. 3c; Supplementary Fig.S3). Changes of these compounds were again stronger in the EBR+CHT co-application treatment. Contrary to the EBR-induced sharp increase in GSH/GSSG ratio, there was a decrease in the GSH/GSSG ratio in CHT-exposed plants and the BR-deficient mutant dim plants (Supplementary Fig.S3). All these results suggest that BR is capable of inducing the transcripts and metabolism associated with CHT degradation in tomato plants.

(a) Venn diagram showing the number of genes with more than 2-fold changes after treatment with EBR, CHT or EBR+CHT in comparison with the untreated controls. ↑, up-regulated; ↓, down-regulated. (b) Comparison of expression of sets of detoxification genes in EBR, CHT and EBR+CHT treated and control plants. (c) Leaf H2O2 content, glutathione homeostasis and GST activity as influenced by EBR, CHT and EBR+CHT treatments. EBR at 0.1 μM was applied 6 h prior to pesticide application and leaves were taken at 5 d after application of CHT at 11.2 mM. Data are means of four biological replicates (±SD). Means denoted by the same letter did not significantly differ at P < 0.05 according to Tukey's test.
BRs enhance CHT degradation by apoplastic H2O2–dependent increases in glutathione biosynthesis and GST activity
BRs induce H2O2 production at the apoplast15. To examine whether BR-induced H2O2 and subsequent glutathione biosynthesis and regeneration play a role in the CHT degradation, we used diphenyleneiodonium (DPI), an inhibitor of NADPH oxidase, dimethylthiourea (DMTU), a scavenger of H2O2 and 6-aminonicotinamide (6-AN), an inhibitor of the pentose phosphate pathway to block the changes in GSH/GSSG ratio in response to EBR or CHT treatment. EBR-induced glutathione (GSH+GSSG) accumulation, GST activity and the transcripts of the genes involved were all decreased by these inhibitors or scavenger but increased by H2O2 treatment and the effect was especially significant for GST activity (Supplementary Figs. S4 & 5). Consistent with this observation, DPI, DMTU and 6-AN all compromised EBR-induced degradation of CHT in the leaves (Supplementary Fig. 6). All these results suggested that BR-induced H2O2 accumulation and associated GSH biosynthesis and regeneration played an important role in BR-induced CHT degradation.
To provide genetic evidence for the role of apoplastic H2O2, glutathione biosynthesis and regeneration in CHT metabolism, we silenced genes involved in the apoplastic H2O2 production (RBOH1), GSH biosynthesis (GSH1 and GSH2) and regeneration (GR1) and genes encoding GSTs. Plants silenced with BRI1 (pTRV- BRI1) and RBOH1 (pTRV- RBOH1) all showed similar GSH and GSSG accumulation and GST activity as compared to the pTRV plants (Fig. 4). Importantly, EBR- and CHT-induced GSH and GSSG accumulation and GST activity were largely compromised in the pTRV- RBOH1 and pTRV- BRI1 plants (Fig. 4). Meanwhile, pTRV- RBOH1 plants showed increased CHT accumulation as compared to pTRV plants and CHT accumulation in pTRV- RBOH1 was not influenced by EBR application (Fig. 5a). In comparison, plants silenced for GSH1 (pTRV-GSH1), GSH2 (pTRV-GSH2) and GR1 (pTRV-GR1) all showed increased CHT accumulation, which was associated with decreased GSH and GSSG accumulation and GST activity as compared to those of the pTRV plants (Fig. 5a, Supplementary Fig. S7). Importantly, EBR co-treatment decreased CHT residue in pTRV, pTRV-GSH1 and TRV-GR plants but not in TRV-GSH2 plants. Furthermore, EBR-induced increases in GSH and GSSG accumulation, the GSH/GSSG ratio and GST activity were abolished by the silencing of GSH1, GSH2 and GR (Supplementary Fig. S7). All these results indicate that BR-enhanced CHT degradation is dependent not only on H2O2 production but also on biosynthesis and regeneration of glutathione.

Involvement of BRI1 and RBOH1 in brassinosteroid (BR) and chlorothalonil (CHT)-regulated glutathione metabolism and GST activity.
EBR at 0.1 μM was applied 6 h prior to pesticide application and leaves were taken at 5 d after application of CHT at 11.2 mM. Data are means of four biological replicates (±SD). Means denoted by the same letter did not significantly differ at P < 0.05 according to Tukey's test.

(a) Chlorothalonil (CHT) residues in RBOH1-, GSH1-, GSH2- and GR- silenced plants with or without EBR treatment. (b) Chlorothalonil (CHT) residues in GST1-7 silenced plants with or without EBR treatment. EBR at 0.1 μM was applied 6 h prior to pesticide application and leaves were taken at 5 d after application of CHT at 11.2 mM. Data are means of four biological replicates (±SD). Means denoted by the same letter did not significantly differ at P < 0.05 according to Tukey's test.
It is known that GST conjugates glutathione to xenobiotics, thus converting the xenobiotics to nonreactive water-soluble conjugates that are easily excreted9. The pTRV-GST1, pTRV-GST3, pTRV-GST4, pTRV-GST6 and pTRV-GST7 plants all showed an increased CHT accumulation as compared to the pTRV plants (Fig. 5b). In contrast, CHT residues in pTRV-GST2 and pTRV-GST5 plants were not significantly different from those in pTRV control plants. Importantly, EBR co-treatment decreased CHT accumulation in the pTRV, pTRV-GST1 and pTRV-GST 4 plants, but had little effect on CHT residues in the pTRV-GST2, pTRV-GST3, pTRV-GST6 and pTRV-GST7 plants. A comparison of the transcripts revealed that transcript levels of GST3, GST6 and GST7 were affected more significantly by EBR than by CHT (Fig. 3b). Taken together, EBR-accelerated CHT degradation is also dependent on some members (e.g. GST3, GST6 and GST7) of the GST gene family in tomato plants.
Discussion
Currently, efforts to reduce health risk from pesticide residues in agricultural produce have been largely through regulations or product cancellations and there are few studies that focused on reducing pesticide residue in intact plants, mostly due to lack of understanding of the mechanisms of pesticide metabolism and its regulation pathway in plants. As shown in this comprehensive study, altered BR levels or BR signaling significantly changed the efficiency of pesticide metabolism in plants, which was associated with altered expression of a subset of detoxifying genes, GSH biosynthesis and the GST activity (Figs. 2,3). Thus, endogenous BRs are critical for the induction of detoxifying response against pesticides.
Although many studies revealed that phytohormones are involved in the response against both biotic and abiotic stresses, little is known about their role in the regulation of xenobiotics metabolism in plants. Our previous study has shown that exogenous applied BR decreased pesticide residue in cucumber16. In this study, we have shown that the effects of EBR on pesticide metabolism are not unique to cucumber but also effective in a subset of crops including tomato, rice, tea and broccoli etc. with an array of pesticides (Fig. 1). Importantly, reduction of BRs level in dim mutant plants or silencing of BRI1 (pTRV- BRI1), which encodes BRs receptor significantly impaired the capacity of pesticide metabolism, indicating that endogenous BRs were critical for the induction of detoxification in response to pesticides.
From microarray data, it is clear that the BR response involves induction of various detoxifying genes, especially those encoding cytochrome P450, GSH, GST and transporters, with corresponding induction of enzyme activities and this is especially significant in the presence of CHT. The similarity of transcriptional profile of CHT- and EBR- treated plants suggested that both responses shared common signaling components. In fact, an array of genes involved in hormonal, stress and redox response are regulated by both CHT and EBR. These genes included MYB-related factors, WRKY, NAC domain factor, cytochrome P450, GST and transporters and ethylene response factor. There are many reports showing the crosstalk of BR with ethylene, jasmonic and ABA17,18,19,20. Therefore, the pesticide metabolism was thought to be influenced by hormonal status and developmental and environmental factors.
Diverse xenobiotics such as barbiturates, pesticides, herbicide safeners and chlorophenols trigger a xenobiotic defense response in plants21,22. How pesticide is perceived and the signaling is transmitted within plant cells remains elusive. In mammals, the cysteine of receptor protein for xenobiotics was modified resulting in release of Nrf2 transcription factor into nucleus where it binds to regulatory antioxidant responsive elements, leading to the induction of genes encoding detoxifying enzymes23,24. Various xenobiotics are known to induce ROS production and ROS can function as cellular second messengers that are likely to modulate many different genes and proteins thus leading to a variety of responses25. Both microarray and RT-PCR data have identified the RBOH1 to be upregulated by CHT and BR. Consistent with the upregulation of RBOH1, treatment of CHT and EBR induced significant increase in H2O2. RBOH encoded the essential components of a plant NADPH oxidase and is required for the ROS production at the apoplast and signal amplification in defense and stress responses26,27,28. We have recently found that modulation of plant stress response by BR was dependent on H2O215. H2O2 plays an important role in upregulation of defense gene expression and antioxidant capacity. In accordance with this, induction of H2O2 content after treatment with CHT and EBR was associated with increased expression of GST and pool of GSH. More importantly, blocking the apoplastic H2O2 accumulation by the RBOH1 silencing, NADPH oxidase inhibitor DPI or ROS scavenger DMTU all significantly inhibited the induction of CYP724B2, GST2, GSH2 and ABC, the increase of GSH pool and pesticide metabolism efficiency. This suggested the involvement of apoplastic H2O2 signal in mediating the induction of detoxifying genes in response to pesticides. This hypothesis is also supported by the upregulation of several detoxifying genes, including GST, P450 and UGT in cat2 deficient Arabidopsis, which had increased accumulation of H2O229.
H2O2 also enhances the activity of enzymes involved in sulfate assimilation and Cys or GSH biosynthesis30. In our study, EBR enhanced GSH biosynthesis in response to CHT treatment, probably through increasing the H2O2 signal which regulates both GSSG regeneration by GR and GSH biosynthesis30,31. Meanwhile, BR-induced H2O2 production in the apoplast could increase the CO2 assimilation which could enhance GSH biosynthesis by providing the carbon skeleton for sulphur assimilation14,32. The increase in the GSH level led to a higher GSH/GSSG, which may act as signal regulating the activity of transcription factors, enzymes and other proteins. It is apparent that the transcript of detoxification related gene such as P450, GSH2, GST2 and ABC as well as the GST activity were all dependent on the ROS accumulation and the cellular glutathione redox statue (Supplementary Fig. S4 & 5), suggesting that the pesticide detoxification process are largely dependent on the redox homeostasis within the cells. Recently, TGA transcription factors have been implicated in a general detoxification network, presumably via activation of redox signaling22,25. The reduction of redox states induced by BR as shown by the increased GSH/GSSG ratio was correlated well with the expression of detoxifying gene expression and pesticide metabolism in this study. Therefore, pesticides like CHT may utilize a similar redox-regulated protein through thiol modification to induce detoxifying genes present in mammals.
Among the detoxification processes of pesticides, GST was the putative major player which catalyzes the conjugation of GSH to pesticides33. Recently, we have also shown that activity and expression of GST was preferentially induced by CHT in a concentration and time-dependent manner34. This indicated that pesticide response was conserved in mammals, microbes and plants. The activation sequence-1 (as-1)-like elements in the promoter of some GST genes has been found to be responsive to ROS35. In agreement with this, different GST genes were differentially induced by EBR and silencing of RBOH1 or BRI1 compromised EBR- and CHT-induced increase in GST activity. All these results provided evidence that BRs-induced changes in cellular redox statue played a critical role in the modulation of GST activity. It seems likely that different GST genes play different roles in the metabolism of different pesticides or subject to the regulation of BRs or redox homeostasis differentially as evidenced by the different induction of GST gene transcript and the CHT residue results in the GST-VIGS plants (Fig. 5).
Lethal and sublethal poisoning due to pesticide residues on crop produce is a serious threat to human health - a problem that is especially widespread in the developing countries due to poor regulations and use of older generations of pesticides. This challenge has remained largely unresolved to date. Here, we have demonstrated that BRs are capable of mitigating residues of a variety of pesticides in different crops in addition to their role in plant growth, development and stress tolerance13. In-depth experiments on tomato further revealed that BRs enhanced pesticide degradation most likely by enhancing glutathione metabolism and GST activity via a RBOH1-dependent pathway (Fig. 4). This function of BRs appears to be applicable for a variety of pesticides in a wide range of crops. This discovery may provide us with the possibility of minimizing pesticide residues on crop produce and the associated risks. To our knowledge, this is the first report with genetic evidence showing that pesticide degradation by an enzymatic system in higher plants is regulated by the phytohormone signaling. In addition to the foliar application of BRs, other means such as genetic engineering of BR biosynthesis and signaling could be exploited to minimize the risk of pesticide contamination in food crops. Recently, BRs were recognized and explored as a biotechnological target for enhancing crop yields and stress tolerance36 and our findings probably offer another exciting potential for improving the safety of human foods.
Methods
Plant materials and virus-induced gene silencing (VIGS) of detoxification related genes
Tomato seeds (Solanum lycopersicum cv Condine Red) and its corresponding BR-deficient mutant (dim) were obtained from TGRC (Tomato Genetic Resource Center) at University of California, Davis. The plants were raised in plastic pots (15 cm diameter and 15 cm height, one seedling per pot) filled with a mixture of peat and vermiculite (7:3, v/v) and were watered daily with Hoagland nutrient solution. The growth conditions were as follows: a 12-h photoperiod, temperature of 25/20°C (day/night) and photosynthetic photon flux density (PPFD) of 600 μmol m−2 s−1.
The tobacco rattle virus (TRV) VIGS vectors used for silencing of tomato genes were constructed as described previously37. The cDNA fragments for BRI1, RBOH1, GSH1, GSH2, GR and GST1-7 were PCR-amplified using the gene-specific primers listed in Supplemental Table 2. The amplified fragment was digested with SacI and XhoI and ligated into the same sites of pTRV2. The resulting plasmid was mobilized into Agrobacterium tumefaciens GV310. VIGS was performed by infiltration on 15-d-old seedlings with a mix of pTRV1- and pTRV2-carrying Agrobacterium tumefaciens. Plants were then kept at 23/21°C under PPFD of 200 μmol m−2 s−1 for 30 d before they were used for the experiments.
Chemical treatments
When the 6th true leaf fully expanded, the plants were pretreated with different concentrations of 24-epibrassinolide (EBR, Sigma, USA). After 6 h, the plants were treated by application of chlorothalonil (CHT) at 11.2 mM with 30 mL per plant. Distilled water was sprayed onto tomato leaves as a control treatment. To determine the role of H2O2 and glutathione homeostasis in the regulation of pesticide metabolism, we pretreated leaves with 100 μM diphenyl iodonium (DPI)38, 5 mM dimethylthiourea (DMTU) and 5 mM 6-aminonicotinamide (6-AN)39. Then 0.1 μM EBR and CHT were applied 8 h and CHT 14 h later, respectively. Plant samples were harvested 5 d after the CHT treatment for analysis of CHT residues. The plants were also sampled at 5 d after pesticide treatment for molecular and biochemical analysis.
To investigate the effects of EBR on the degradation of various organophosphorus, organochlorine and carbamate pesticides on various crops, plants of rice, tea, broccoli, strawberry, celery, Chinese chives, asparagus, cucumber, garlic and Chinese cabbage grown in greenhouse were first sprayed with distilled water or 0.1 μM EBR and 6 h later, with chlorpyrifos, phoxim, chlorothalonil, omethoate, cypermethrin, carbofuran and 3-hydroxycarbofuran at commercially recommended doses. After 7 d, tissue samples were taken for the analysis of pesticide residues.
Pesticide residue analysis
To determine pesticide residues in the tested plants, 25 g (fresh weight) of tissue were homogenized for 2 min in 50 mL acetonitrile. The liquid phase was transferred into a separatory funnel containing 5–7 g of anhydrous sodium sulfate and mixed for 5 min. The acetonitrile extract was collected and concentrated to near dryness and then dissolved in 5 mL acetone. Chlorpyrifos, phoxim, omethoate, chlorothalonil and cypermethrin were quantitatively measured on an Agilent 6890 N gas chromatography system (Agilent Technologies, Palo Alto, CA, USA), equipped with FPD or ECD and a DB-17 silica capillary column. For the determination of carbofuran and 3-hydroxycarbofuran, the concentrated mixture was transferred to a Supelclean LC-NH2 SPE (Supelco, Poole, UK), eluted with 4 mL of methanol-methylene dichloride (1:99, v/v), concentrated to near dryness and then dissolved in methanol for analysis on an Agilent 1200 high pressure liquid chromatography (Santa Clara, CA, USA) with FLD equipped with a ZORBAX SB-C18 column (25 cm × 4.6 mm × 5 mm) after derivatization with O-phthaladehyde.
Biochemical analysis
H2O2 was extracted from leaf tissues with 0.2 M HClO4. The homogenate was centrifuged at 2,700 g for 30 min at 4°C and the supernatant was collected, adjusted to pH 6.0 with 4 M KOH and centrifuged at 11,000 g for 10 min at 4°C. The supernatant was placed onto an AG1x8 prepacked column (Bio-Rad, Hercules, CA) and H2O2 was eluted with 4 mL double-distilled H2O. The sample (800 μL) was mixed with 400 μL reaction buffer containing 4 mM 2,2′-azino-di(3-ethylbenzthiazoline-6-sulfonic acid) and 100 mM potassium acetate at pH 4.4, 400 μL deionized water and 0.25 U of horseradish peroxidase. The content of H2O2 was measured at OD412 as described previously40.
Reduced and oxidized glutathione (GSH and GSSG) were determined as described previously. Briefly, plant leaf tissue (0.2 g) was homogenized in 2 mL of 2% metaphosphoric acid containing 2 mM EDTA and the homogenate was centrifuged at 4°C for 10 min at 14,000 g. After neutralization with 0.5 M phosphate buffer (7.5), oxidized glutathione (GSSG) and reduced glutathione (GSH) in the supernatant were quantified using an enzymatic recycling assay as described in Queval & Noctor41.
Enzyme activity assay
Glutathione reductase (GR) and glutathione S-transferase (GST) were extracted from 0.3 g of leaves with 2 mL extraction buffer (50 mM, potassium phosphate, pH 7.5) containing 10 mM KCl, 1 mM EDTA, 5 mM dithiothreitol (DTT), 0.5 mM AEBSF and 1:4 (w/w) polyvinypolypyrrolidone (insoluble PVPP). The activity of GR was measured according to Foyer & Halliwell42 based on the rate of decrease in the absorbance of NADPH at 340 nm. The activity of GST was assayed spectrophotometrically according to Habig & Jakoby43. Spectrophotometric analysis was conducted on a SHIMADZU UV-2410PC spectrophotometer (Shimadzu, Kyoto, Japan).
RNA extraction and qRT-PCR for gene expression analysis
Total RNA was isolated from tomato leaves using TRIZOL reagent (Sangon, China) and the cDNA template for qRT-PCR was synthesized using a RevertAid™ first strand cDNA synthesis kit (Fermentas) by following the manufacturer's instructions. qRT-PCR was performed with an iCycler iQ Multicolor Real-time PCR detection system (Bio-Rad, Hercules, CA). Gene-specific primer pairs were designed with Primer Premier 5.0 (Premier Biosoft International, Palo Alto, CA) and are listed in Supplemental Table 3. Tomato actin gene was used as an internal control. Relative gene expression was calculated according to Livak & Schmittgen44.
Microarray analysis
Affymetrix GeneChip Tomato Genome Array, which is designed specifically to monitor gene expression in tomato, was used for analyzing the gene expression at the whole genome level (http://www.affymetrix.com/products services/arrays/specific/tomato.affx). RNA was extracted from leaves treated with distilled water, EBR, CHT or EBR+CHT using a Qiagen RNeasy kit according to the manufacturer's instructions. Each treatment had three biological replicates. cDNA was prepared using the two cycle target labeling procedure and was used for further synthesis of labeled target copy RNA by in vitro transcription as described in the Affymetrix GeneChip manual. All procedures for probe preparation, hybridization, washing, staining and scanning of the GeneChip Tomato arrays, as well as data collection, were performed at the Beijing Biochip National Engineering Research Center, Beijing, China. Signal summarization, normalization and background subtraction were performed using Robust Multichip Analysis45 in the affymetrix package with default parameters. The statistical test for differentially expressed genes was performed using the LIMMA (linear models for microarray) package46. Genes showing a 2-fold change in the expression level between treatment and control at P < 0.05 were defined as significantly differentially expressed genes. Gene Ontology analysis was carried out by using the BLAST tool in order to match the tomato transcripts to the closest homologs of Arabidopsis with an e value of less than e−5.
Statistical analysis
Data were subjected to analysis of variance and the means were compared using Tukey's test at the 5% level.
References
Oerke, E. C. Crop losses to pests. J. Agri. Sci. 144, 31 (2006).
Owens, K., Feldman, J. & Kepner, J. Wide range of diseases linked to pesticides. Pestic. You 30, 13–21 (2010).
Whitehorn, P. R., O'Connor, S., Wackers, F. L. & Goulson, D. Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 336, 351–352 (2012).
Peiris-John, R. J. & Wickremasinghe, R. Impact of low-level exposure to organophosphates on human reproduction and survival. Trans. R. Soc. Trop. Med. Hyg. 102, 239–245 (2008).
Giasson, B. T. & Lee, V. M. Y. A new link between pesticides and Parkinson's disease. Nature Neurosci. 3, 1227–1228 (2000).
Alavanja, M. C. R., Ross, M. K. & Bonner, M. R. Increased cancer burden among pesticide applicators and others due to pesticide exposure. CA-Cancer J. Clin. 63, 120–142 (2013).
Eaton, D. L. et al. Review of the toxicology of chlorpyrifos with an emphasis on human exposure and neurodevelopment. Crit. Rev. Toxicol. 38, 1–125 (2008).
Xu, C. J., Li, C. Y. T. & Kong, A. N. T. Induction of phase I, II and III drug metabolism/transport by xenobiotics. Arch. Pharm. Res. 28, 249–268 (2005).
Muntane, J. Regulation of drug metabolism and transporters. Curr. Drug Metab. 10, 932–945 (2009).
Rushmore, T. H. & Kong, A. N. T. Pharmacogenomics, regulation and signaling pathways of phase I and II drug metabolizing enzymes. Curr. Drug Metab. 3, 481–490 (2002).
Wakabayashi, N. et al. Protection against electrophile and oxidant stress by induction of the phase 2 response: Fate of cysteines of the Keap1 sensor modified by inducers. Proc. Natl. Acad. Sci. U. S. A. 101, 2040–2045 (2004).
Ramel, F., Sulmon, C., Serra, A. A., Gouesbet, G. & Couee, I. Xenobiotic sensing and signalling in higher plants. J. Exp. Bot. 63, 3999–4014 (2012).
Gudesblat, G. E. & Russinova, E. Plants grow on brassinosteroids. Curr. Opin. Plant Biol. 14, 530–537 (2011).
Jiang, Y. P. et al. Cellular glutathione redox homeostasis plays an important role in the brassinosteroid-induced increase in CO2 assimilation in Cucumis sativus. New Phytol. 194, 932–943 (2012).
Xia, X. J. et al. Reactive oxygen species are involved in brassinosteroid-induced stress tolerance in cucumber. Plant Physiol. 150, 801–814 (2009).
Xia, X. J. et al. Brassinosteroids promote metabolism of pesticides in cucumber. J. Agri. Food Chem. 57, 8406–8413 (2009).
Ephritikhine, G., Fellner, M., Vannini, C., Lapous, D. & Barbier-Brygoo, H. The sax1 dwarf mutant of Arabidopsis thaliana shows altered sensitivity of growth responses to abscisic acid, auxin, gibberellins and ethylene and is partially rescued by exogenous brassinosteroid. Plant J. 18, 303–314 (1999).
Xiong, L. M., Schumaker, K. S. & Zhu, J. K. Cell signaling during cold, drought and salt stress. Plant Cell 14, S165–S183 (2002).
Rodrigues, A. et al. The short-rooted phenotype of the brevis radix mutant partly reflects root abscisic acid hypersensitivity. Plant Physiol. 149, 1917–1928 (2009).
Divi, U. K., Rahman, T. & Krishna, P. Brassinosteroid-mediated stress tolerance in Arabidopsis shows interactions with abscisic acid, ethylene and salicylic acid pathways. BMC Plant Biol. 10, 151 (2010).
Baerson, S. R. et al. Detoxification and transcriptome response in Arabidopsis seedlings exposed to the allelochemical benzoxazolin-2(3H)-one. J. Biol. Chem. 280, 21867–21881 (2005).
Riechers, D. E., Kreuz, K. & Zhang, Q. Detoxification without intoxication: herbicide safeners activate plant defense gene expression. Plant Physiol. 153, 3–13 (2010).
Balogun, E. et al. Curcurnin activates the haem oxygenase-1 gene via regulation of Nrf2 and the antioxidant-responsive element. Biochem. J. 371, 887–895 (2003).
Kobayashi, A. et al. Oxidative stress sensor Keap1 functions as an adaptor for Cul3-based E3 ligase to regulate for proteasomal degradation of Nrf2. Mol. Cell. Biol. 24, 7130–7139 (2004).
Ramel, F., Sulmon, C., Serra, A., Gouesbet, G. & Couee, I. Xenobiotic sensing and signaling in higher plants. J. Exp. Bot. 63, 3999–4014 (2012).
Torres, M. A., Dangl, J. L. & Jones, J. D. G. Arabidopsis gp91phox homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response. Proc. Natl. Acad. Sci. U. S. A. 99, 517–522 (2002).
Kwak, J. M. et al. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 22, 2623–2633 (2003).
Sagi, M. et al. Plant respiratory burst oxidase homologs impinge on wound responsiveness and development in Lycopersicon esculentum. Plant Cell 16, 616–628 (2004).
Vanderauwera, S. et al. Genome-wide analysis of hydrogen peroxide-regulated gene expression in Arabidopsis reveals a high light-induced transcriptional cluster involved in anthocyanin biosynthesis. Plant Physiol. 139, 806–821 (2005).
Queval, G. et al. H2O2-activated up-regulation of glutathione in Arabidopsis involves induction of genes encoding enzymes involved in cysteine synthesis in the chloroplast. Mol. Plant 2, 344–356 (2009).
Mhamdi, A. et al. Arabidopsis GLUTATHIONE REDUCTASE1 plays a crucial role in leaf responses to intracellular hydrogen peroxide and in ensuring appropriate gene expression through both salicylic acid and jasmonic acid signaling pathways. Plant Physiol. 153, 1144–1160 (2010).
Kopriva, S. & Rennenberg, H. Control of sulphate assimilation and glutathione synthesis: interaction with N and C metabolism. J. Exp. Bot. 55, 1831–1842 (2004).
Kim, Y. M. et al. Glutathione-dependent biotransformation of the fungicide chlorothalonil. J. Agric. Food Chem. 52, 4192–4196 (2004).
Wang, J. T. et al. The different responses of glutathione-dependent detoxification pathway to fungicide chlorothalonil and carbendazim in tomato leaves. Chemosphere 79, 958–965 (2010).
Garreton, V., Carpinelli, J., Jordana, X. & Holuigue, L. The as-1 promoter element is an oxidative stress-responsive element and salicylic acid activates it via oxidative species. Plant Physiol. 130, 1516–1526 (2002).
Vriet, C., Russinova, E. & Reuzeau, C. Boosting crop yields with plant steroids. Plant Cell 24, 842–857 (2012).
Ekengren, S. K., Liu, Y. L., Schiff, M., Dinesh-Kumar, S. P. & Martin, G. B. Two MAPK cascades, NPR1 and TGA transcription factors play a role in Pto-mediated disease resistance in tomato. Plant J. 36, 905–917 (2003).
Miller, G. et al. The Plant NADPH oxidase RBOHD mediates rapid systemic signaling in response to diverse stimuli. Sci. Signal. 2, ra45 (2009).
Mou, Z., Fan, W. H. & Dong, X. N. Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell 113, 935–944 (2003).
Willekens, H. et al. Catalase is a sink for H2O2 and is indispensable for stress defence in C-3 plants. EMBO J. 16, 4806–4816 (1997).
Queval, G. & Noctor, G. A plate reader method for the measurement of NAD, NADP, glutathione and ascorbate in tissue extracts: Application to redox profiling during Arabidopsis rosette development. Anal. Biochem. 363, 58–69 (2007).
Foyer, C. H. & Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 133, 21–25 (1976).
Habig, W. H. & Jakoby, W. B. [Assays for differentiation of glutathione S-transferases]. Methods in Enzymology [Jakoby, W. B. (ed.)] [398-405] (Academic Press, New York, 1981).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25, 402–408 (2001).
Irizarry, R. A. et al., Summaries of affymetrix GeneChip probe level data. Nucleic Acids Res. 31, e15 (2003).
Smyth, G. K. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol. 3, 3 (2004).
Acknowledgements
Supported by grants from the National Basic Research Program of China (2009CB119000), the National Natural Science Foundation of China (31071790) and the National Key Technology R&D Program of China (2011BAD12B04).
Author information
Authors and Affiliations
Contributions
Z.C., J.G. and J.Y. designed the research; Y.Z., X.X., G.Y., J.W., J.W., M.W., Y.Y., K.S. and Y.Y. performed the experiments; Z.C., J.G. and J.Y. wrote the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplemental tables and figures
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zhou, Y., Xia, X., Yu, G. et al. Brassinosteroids play a critical role in the regulation of pesticide metabolism in crop plants. Sci Rep 5, 9018 (2015). https://doi.org/10.1038/srep09018
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep09018
This article is cited by
-
Exogenous Epigallocatechin-3-Gallate Alleviates Pesticide Phytotoxicity and Reduces Pesticide Residues by Stimulating Antioxidant Defense and Detoxification Pathways in Melon
Journal of Plant Growth Regulation (2024)
-
24-Epibrassinolide alleviates diazinon oxidative damage by escalating activities of antioxidant defense systems in maize plants
Scientific Reports (2023)
-
Foliar-Applied Melatonin Mitigates Carryover Injury Caused by Premix Herbicide Lumax®537.5SE in Potato
Potato Research (2023)
-
Enhancing crop health and sustainability: exploring the potential of secondary metabolites and non-thermal plasma treatment as alternatives to pesticides
Plant Biotechnology Reports (2023)
-
Significance of brassinosteroids and their derivatives in the development and protection of plants under abiotic stress
Biologia (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
