
Abstract
The brain's impotence to utilize long-chain fatty acids as fuel, one of the dogmas in neuroscience, is surprising, since the nervous system is the tissue most energy consuming and most vulnerable to a lack of energy. Challenging this view, we here show in vivo that loss of the Drosophila carnitine palmitoyltransferase 2 (CPT2), an enzyme required for mitochondrial β-oxidation of long-chain fatty acids as substrates for energy production, results in the accumulation of triacylglyceride-filled lipid droplets in adult Drosophila brain but not in obesity. CPT2 rescue in glial cells alone is sufficient to restore triacylglyceride homeostasis and we suggest that this is mediated by the release of ketone bodies from the rescued glial cells. These results demonstrate that the adult brain is able to catabolize fatty acids for cellular energy production.
Similar content being viewed by others

Introduction
To guarantee organismal survival, a continuous supply of ATP for cellular energy production is maintained by many regulatory mechanisms at the cellular, endocrine and behavioral level. Under normal feeding conditions, ATP is generated from different substrates, such as carbohydrates, amino and fatty acids, delivered to the various tissues via the circulation. When calories are oversupplied, triacylglycerides (TAG) are synthesized from the different substrates and stored in specific cells, so called adipocytes. Excess body TAG storage is called obesity, a condition that is linked to increased morbidity and mortality. During prolonged starvation, cellular TAG mobilization and catabolism becomes vital. The long-chain fatty acid (LCFA) groups are released from the glycerol backbone and activated with a Coenzyme A (CoA) residue. LCFA-CoA is catabolized through β−oxidation, either in mitochondria or in peroxisomes, but only mitochondrial β-oxidation is coupled to ATP production1. LCFA-CoA does not cross the tight inner mitochondrial membrane: to cross, the CoA residue is replaced with a carnitine residue, a reaction that is catalyzed by carnitine palmitoyltransferase 1 (CPT1). LCFA-carnitine is transported with carnitine-acylcarnitine translocase (CACT) across the inner membrane. Inside mitochondria, carnitine needs to be replaced again by CoA, a reaction catalyzed by carnitine palmitoyltransferase 2 (CPT2), yielding LFCA-CoA that can be catabolized through β−oxidation and generate ATP2,3,4.
Human patients with severe mutations in one of the enzymes of the carnitine shuttle system, CPT1 or CPT2, die early after birth. Patients with less severe mutations are sensitive to starvation throughout life. This is probably a direct consequence of the failure to generate ATP through the TAG-β−oxidation pathway5,6 and illustrates the critical importance of this pathway.
Although CPT2 deficiency patients have brain defects7,8 and CPT1 and CPT2 expression has been demonstrated in adult brain5,9, it has been stated that the brain does not use β−oxidation for energy production10. In fact, under conditions of starvation brain metabolism is thought to be restricted to use ketone bodies derived from β-oxidation of long-chain fatty acids in the liver. It has been argued that the reason behind this might be that β−oxidation11 is associated with the production of free oxygen radicals, causing neuronal loss. While liver or muscle cells, where β−oxidation occurs at high rates during starvation, are replaceable, brain cells are post-mitotic, unique in terms of circuitry and therefore not easily, if at all, replaceable. Another reason for the denial of β−oxidation in the brain may reside in the limited space in the scull or head capsule to allow for extensive lipid storage without compromising brain function.
To challenge the view that the brain does not use long chain fatty acids for energy production, we here directly tested the contribution of the brain in organismal β−oxidation by removing dCPT2 in Drosophila melanogaster. Our results show that β−oxidation does in fact occur in the glial cells of the adult Drosophila brain in vivo and may contribute to organismal energy supply.
Results
Drosophila CPT2 deficiency resembles human CPT2 deficiency
To determine the importance of fatty acid catabolism for energy production in brain, we searched the Drosophila melanogaster genome for the orthologue of the highly conserved bottleneck enzyme of β-oxidation, carnitine palmitoyltransferase 2 (CPT2).
Based on BLAST search on the Drosophila genome, we identified CG2107, from now on called dCPT2 (Fig. 1a), on chromosome 3L and generated loss-of function alleles by imprecise excision of P-element P(GSV7)GS21694 situated in the 5′ UTR of the only dCPT2 transcript, 92 base pairs before the translational start site. Two mutant alleles, dCPT2δ3 and dCPT2δ18, we considered null mutants, since a major part of the ORF (508 and 124 of the 667 amino acids, respectively) was deleted (Fig. 1b) and both mutants failed to complement each other, or Deficiencies Df(3L)BSC428 and Df(3L)BSC671 spanning the CG2107 locus (Fig. 1c). As wild type control line in our experiments we used the revertant line giving rise to a precise excision of P(GSV7)GS21694.

CG2107 mutants and rescue with mammalian CPT2.
(A) Protein sequence alignment of Drosophila melanogaster (DM), Drosophila simulans (Dsim), Drosophila pseudoobscura (Dpse), Danio rerio (danio), gallus gallus (chick), homo sapiens (human), mus musculus (mouse), rattus norvegicus (rat) and consensus CPT2. (B) Region 63A2 of Drosophila melanogaster chromosome 3L, bearing dCPT2 (CG2107), P-element insertions, deficiencies Df(3l)428 and Df(3L)671 spanning the CG2107 locus and the sequences deleted in two newly generated mutants dCPT2 δ18 and δ3. (C) Eclosion rates of dCPT2 mutant flies with the indicated genotypes in percent of expected (n > = 3 experiments, **** p < 0.0001). Rescue was performed with wild type (rCPT2) or mutant (rCPT2 H372A) rat CPT driven by the ubiquitous driver da-Gal4. (D) Western blot from mouse liver or whole fly extract from wild type or rCPT2 expressing flies, stained with anti-CPT2 to confirm ectopic expression in rescue flies (65 kDa band marked with an arrow), or anti-Histone 3 as loading control.
The majority (62%) of the dCPT2δ3 mutant flies died within the first three days after eclosion, reminiscent of the postnatal lethality in severe human CPT2 deficiency patients. This phenotype was fully rescued with ubiquitous expression of wild type rat CPT2 (rCPT2) (p = 0.0003), but not with inactive rat CPT2 bearing the H372A mutation of the catalytic site that gives rise to human disease12 (Fig. 1c). This confirms that CG2107 is indeed orthologous to mammalian CPT2 and that dCPT2δ3, from now on called dCPT2−/−, specifically affects dCPT2. The ectopic expression of rat CPT2 in flies was confirmed by Western blot (Fig. 1d).
dCPT2 is required in adult life
Developmental analysis showed that most of the dCPT2−/− lethality occurred in the first three days after eclosion (p = 0.003) and very little during larval or pupal development (Fig. 2a). dCPT2−/− flies that survived the immediate post-eclosion period still showed a severely reduced adult lifespan of 22 and 10 days in females and males, respectively, compared to 47 and 42 days in control females and males (Fig. 2b). The reduced lifespan could be fully reversed by ubiquitous rescue of CPT2. This indicates that flies require β-oxidation for survival during adulthood.

Phenotype of dCPT2−/− animals.
(A) Percent survival of wild type (Rev) or dCPT2−/− larvae, pupae or young adults, in percent (n > = 3 experiments; ** p < 0.01; ***p < 0.005). (B) Lifespan of wild type or dCPT2−/− adults with or without ubiquitous rescue with rCPT2, in days +/− standard error of the means (n > = 4 experiment for a total of > = 105 animals per genotype). (C) Percent survival of starving 1 week old adults on 1% agarose, in hours +/− standard error of the means (n = 3 for a total of > = 209 animals per genotype). (D) Weight per adult fly, in mg (n > = 394; * p < 0.05; ***p < 0.005)). (E) Length of adult flies, in mm (n = 10). (F) Ratio of weight per length in mg/mm. (G) Total motor activity time of adult male flies, in counts per 24 h (n = 6).
Another core symptom of human CPT2 deficiency in adults is sensitivity to starvation. Similar to human CPT2 deficiency patients, dCPT2−/− flies were hypersensitive to starvation. Under conditions of complete starvation, the average survival time of dCPT2−/− flies dropped from 63.1 +/− 7.5 hours to 37.5 +/− 2.6 hours in female (p = 0.005) and from 32.0 +/− 3.4 to 21.3 +/− 3.3 hours in male (p = 0.017) dCPT2−/− flies when compared to wild type (Fig. 2c).
No obesity but brain TAG accumulation in adult dCPT2−/− flies
We asked if systemic lack of mitochondrial β-oxidation causes obesity, since it is the main pathway for long-chain fatty acid (LCFA) degradation. In contrast to our expectations, normally fed dCPT2−/− flies were not heavier (Fig. 2d) but even lighter than controls, 0.72 +/− 0.04 mg vs. 0.67 +/− 0.04 mg for male (p = 0.02) and 1.04 +/− 0.05 mg vs. 0.97 +/− 0.02 mg for female (p = 0.002) wild type vs. dCPT2−/− flies, respectively. Body length was also somewhat reduced in dCPT2−/− flies when compared to controls (2.70 +/− 0.13 vs. 2.61 +/− 0.08 mm in female (p = 0.02) and 2.25 +/− 0.11 mm vs. 2.23 +/− 0.16 mm in male (not significant) wild type vs. dCPT2−/− flies, respectively (Fig. 2e), but the ratio of weight and size (BMI) was not increased in dCPT2−/− flies (Fig. 2f). Motor activity over a period of 24 h was also unchanged, reaching from 469 +/− 63 beam passes in control to 535 +/− 70 in heterozygotes to 538 +/− 80 in dCPT2−/− homozygotes (Fig. 2g), ruling out increased muscle work as the cause for the lack of an obesity phenotype in dCPT2−/− animals. Together this suggests that in Drosophila the regulation of energy balance is independent from mitochondrial β-oxidation.
Whole body neutral lipid was not increased in fed dCPT2−/− flies. In fed male dCPT2−/− flies, TAG levels were identical in both dCPT2−/− and wild type flies. In fed female dCPT2−/− flies, TAG levels were even lower than in wild type controls (Fig. 3a). Total TAG was turned over at the same rate in dCPT2−/− and wild type flies under starvation conditions (Fig. 3a). This result further strengthens the notion that pathways other than mitochondrial β-oxidation regulate TAG turnover at the organismal level.

Lipid composition of dCPT2−/− flies and TAG accumulation in dCPT2−/− brains.
(A) Protein, phospholipid (PL) and triacylglyceride (TAG) content of 1 week old adult female flies of the indicated genotype after the indicated hours of starvation (n = 3 experiments). (B) Phospholipid (PL) and triacylglyceride (TAG) content of 1 week old female adult brains of the indicated phenotype (n = 3 experiments, ***p < 0.001). (C) Thin layer chromatography of lipid extracts of dCPT2−/− or wild type brains, fatty acid (FA; oleic acid), diacylglyceride (DAG; 1,2-diolein) and triacylglyceride (TAG, triolein) as standards. (D) Confocal micrograph of adult brains or flight muscle of the indicated genotype, stained with neutral lipid staining (red). (E) Electron micrograph of wild type or dCPT2−/− brain showing accumulation of TAG in dCPT2−/− brain marked with arrows.
Yet, although TAG levels were not increased in the body of dCPT2−/− flies, TAG accumulated massively in dCPT2−/− brains. Consistent with what is known from mammalian brains, in wild type brains TAG was barely detectable (Fig. 3b, c). TAG levels in wild type fly brain were at the detection limit of 13.4 pmol per brain, while reaching 235.1 pmol TAG per dCPT2−/− brain, equivalent to a 17.5 fold increase (p = 0.0004). Brain phospholipid levels were unchanged, ruling out a simple increase in brain size, brain degeneration or loss of brain tissue due to the lack of dCPT2.
In order to validate and further understand the nature of the TAG accumulations in dCPT2−/− brains, we performed brain whole mount staining with a neutral lipid specific dye13. This confirmed widespread accumulation of neutral lipid droplets in dCPT2−/− brains, while basically no signal was obtained from wild type brains (Fig. 3d, middle panel). Similar results were obtained from fly brains mutant for dCPT1 (Fig. 3d, right panel), another bottle-neck enzyme required for mitochondrial β-oxidation, giving rise to a similar human phenotype. Interestingly, lipid droplets also accumulated in flight muscle (Fig. 3d).
Finally, electron microscopy images from dCPT2−/− but not control brains showed frequent droplet-like inclusions of homogenous appearance, a single outer membrane and variable size, characteristic for TAG containing lipid droplets (Fig. 3e).
Altogether, the last series of results demonstrates massive accumulation of TAG-filled lipid droplets in the brains and muscle of dCPT2−/− animals while total body TAG levels or body were not increased in dCPT2−/− animals.
Glial β-oxidation restores lipid homeostasis
We analyzed next if dCPT2 is expressed in the adult fly brain. Using a knock in line where GFP is fused to dCPT2 in the dCPT2 locus, we could demonstrate endogenous expression of dCPT2 in the brain by confocal microscopy (Supplementary data). Knockdown of dCPT2 by RNA interference specifically in neurons or glia caused a decrease of dCPT2 transcript levels to 67% and 28% of control levels in brain, respectively (Fig. 4a). Considering that in Drosophila the large majority of brain cells are neurons this suggests that in adult fly brain dCPT2 is expressed predominantly in glia and the brain TAG deposits in dCPT2−/− flies are located in glial cells.

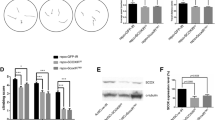
TAG localization in glial lipid droplets and contribution to organismal lipid metabolism.
(A) Relative expression levels of dCPT2 mRNA in adult brain after specific knockdown in neurons (Nsyb-Gal4) or glia (Repo-Gal4) (n = 3 experiments). (B) Confocal micrograph of adult dCPT2−/− brains expressing LSD2-GFP (in green) in neurons (Nsyb-Gal4) (upper row) or glia (Repo-Gal4) (lower row) and stained with neutral lipid staining (NLS) (red). (C) Confocal micrograph of adult brains and muscle from dCPT2−/− or dCPT2−/− with glial dCPT2 rescue (lower) flies stained with neutral lipid staining (red). (D) Eclosion viability of dCPT2−/− flies or dCPT2−/− flies with dCPT2 rescue in glia (Repo-Gal4), muscle (Mef2-Gal4), fat body (Cg-Gal4) or neurons (Nsyb-Gal4) (n = 3 experiments, ***p < 0.001). (E) Lifespan of male or female wild type or dCPT2−/− adults with or without glial rescue with rCPT2, in days +/− standard error of the means (n > = 57 flies). (F) Eclosion viability of dCPT2−/− flies or dCPT2−/− flies with dCPT2 rescue with the indicated glial subtype driver (n = 4 experiments, ***p < 0.001). (G) qPCR results for the indicated transcripts involved in different metabolic pathways, from RNA extracted from heads of the indicated genotypes with or without 24 h starvation (ratio starved/nonstarved) (n = 3 experiments, ***p < 0.01). (H) qPCR results for the indicated transcripts involved in ketone body formation, from RNA extracted from heads of the indicated genotypes with or without 24 h starvation (ratio starved/nonstarved) (n = 3 experiments, *p < 0.05).
To prove this point, we ectopically expressed lipid storage droplet-2 (LSD-2), a molecular marker for lipid droplets14 specifically in neurons or glia by driving UAS-LSD-2 with Nsyb-Gal4 or Repo-Gal4, respectively and analyzed the brains of adult flies by confocal microscopy. We found co-localization of LSD-2 with TAG deposits in glia but not neurons of dCPT2−/− flies (Fig. 4b), demonstrating stabilization of LSD-2 in lipid droplets in glia but not in neurons.
Is glial β-oxidation functionally relevant? To answer this, we performed glial-specific dCPT2 rescue experiments. Glial-specific rescue of dCPT2−/− flies reversed brain TAG accumulation (Fig. 4c) and peri-eclosion lethality (p = 0.0009) (Fig. 4d) and normalized lifespan partially (Fig. 4e).
To specify further the role of dCPT2 in glial cells, we performed rescue experiments in subpopulations of glial cells of dCPT2−/− flies15,16. This revealed that dCPT2 expression in cortex glia (NP2222-Gal4; p = 0.004 and Nrv2-Gal4; p = 0.0009) or astrocyte-like glia (dEAAT1-Gal4; p = 0.0005) but not in ensheathing glia (NP6520) is sufficient to restore eclosion of dCPT2 mutants (Fig. 4f).
To analyze which metabolic changes underlie glial dCPT2−/− function, we compared expression levels of different genes involved in β−oxidation, lipogenesis and gluconeogenesis in heads of starved or non-starved adult wild type, dCPT2−/−, dCPT2−/− with glial dCPT2 rescue and dCPT2−/− with muscular dCPT2 rescue flies. We found that HNF4, the master regulator of the β−oxidation transcriptional program17 was up regulated 1.6 fold in heads of dCPT2−/− glial rescue flies (p = 0.001), but down regulated in heads of wild type (Fig. 4g), dCPT2−/−, or dCPT2−/− flies with muscular rescue, suggesting that the rescued glial cells perform β-oxidation. In contrast, the glial rescue flies did not induce lipogenesis, since SREBP levels remained constant and gluconeogenesis was blocked, as indicated by the increase in PEPK levels (p = 0.006).
How would β-oxidation restricted to glia provide energy to other cells such as neurons or muscle? This could happen in a manner similar to ketone body production in the mammalian liver, where this pathway is up regulated under starvation conditions. Thus we tested the brain for expressional changes of enzymes involved in ketone body generation. Strikingly, all four different enzymes involved in ketone body formation (HMGL, HMGS, CG1140 and CG13377) were up regulated 1.7 to 2.0 fold in dCPT2−/− flies with glial rescue (p = 0.02 to 0.03), but not in wild type, dCPT2−/− or muscular rescue flies (Fig. 4h). This suggests that glial cells can provide energy to the organism by generation and export of ketone bodies derived from β-oxidation.
Discussion
The brain is a metabolically highly active tissue and more sensitive to nutrient and oxygen deprivation than any other organ. In fact, irreversible damage occurs already after minutes of hypoglycemia or ischemia18, while other organs survive for hours. Yet, a central dogma in biology is that β−oxidation of long-chain fatty acids (LCFA), an important source of ATP provision under starvation conditions, is completely absent from brain. Triacylglycerides (TAG) from which LCFA arise are also thought to be absent from this organ.
However, a number of observations challenge the dogma. First, the enzymes involved in mitochondrial β−oxidation are expressed in adult brain9. Second, individuals affected by a mutation in CPT2, an enzyme required for β−oxidation, have a morphological brain phenotype7. Third, glial cells such as astrocytes are known to have a potential to provide metabolites such as lactate to neurons19. Fourth, neurons and glia are known to be able to store TAG under certain in vitro conditions20.
These reasons prompted us to investigate the potential of the brain to perform β−oxidation. We chose to study this under complete absence of β−oxidation in the whole organism, assuming that this might reveal β−oxidation induction deficits in the brain. We used the fruit fly as an animal model that is genetically tractable and created CPT2−/− flies. Those flies showed a phenotype reminiscent of human CPT2 deficiency. They die shortly after birth; the survivors have a reduced lifespan and are hypersensitive to starvation.
In support of our predictions, we could now demonstrate that CPT2 is expressed in the brain, mostly in glia. CPT2−/− but not wild type brains accumulated large amounts of TAG within lipid droplets in vivo, a phenomenon that has been observed only in vitro so far20. Detailed analysis demonstrated specific localization in glial cells, supporting their role as nurturing cells. To test the functional importance of glial cells in β−oxidation, we generated CPT2−/− animals in which CPT2 was rescued in glial cells only. This had both cell autonomous and non-autonomous effects. Autonomously, glial TAG accumulation was completely abolished, indicating that LCFA are degraded in glia and that glial cells are indeed capable of performing β−oxidation. Non-autonomously, organismal lethality and TAG accumulation in muscle is reversed. This could be mediated by an effect on the brain directly, causing improved brain function. Alternatively, glial cells could release energy metabolites such as ketone bodies back into the hemolymph for systemic use21. In support of the latter possibility, we found that enzymes involved in ketone body formation are up regulated in starving brains expressing CPT2 only in glia. This result is in turn consistent with early work showing that astrocytes in culture have the potential to produce ketone bodies from fatty acids22, altogether suggestive that the adult brain, under special circumstances like starvation, may act as a coordinator of organismal energy metabolism23.
Interestingly, CPT2−/− flies are not obese. This suggests that β−oxidation is not rate limiting for organism-wide energy storage in vivo and therefore not a potential target for the treatment of obesity, a condition associated with increased risk for age-related diseases and associated with a shortened lifespan24. Alternative mechanisms must exist that regulate the balance between organismal energy uptake and usage.
Our data are consistent with the following model of β−oxidation: (A) In wild type animals β−oxidation occurs in different tissues: TAG do not accumulate and enough energy is provided for survival. (B) When β−oxidation is ubiquitously impaired (such as in hereditary diseases), not enough energy can be provided leading to organismal dysfunction and death. In these situations TAG accumulate in the brain. (C) Allowing the animals with reduced (blocked) β−oxidation to perform β−oxidation specifically in glia results in energy production from LCFA β−oxidation and the export of ketone bodies to other tissues. This reverses TAG accumulation and prevents death. In support, a number of publications have reported that fatty acid oxidation/ketone body formation play a protective role.
The brain is a privileged organ in terms of substrate use for cellular energy production25. However, the brain is not completely but only relatively different from body cells, because in case of absolute need it is also capable of catabolizing LCFA for its own and even for systemic needs. This becomes relevant in ageing or obese organisms, where the substrate restriction might be increasingly loosened and β−oxidation less repressed26, as indicated by increasing expression of β−oxidation genes in ageing brains27,28. This might contribute to ageing phenotypes of the brain and increasing prevalence of age-related neurodegenerative diseases such as Alzheimer or Parkinson disease.
Methods
Stocks
P(GSV7)GS21694, CPT1 EY20655/CyO, Df(3l)BSC428, Df(3L)BSC671 were received from Bloomington stock center. UAS > LSD2-GFP was received from R. Kühnlein (MPI Göttingen); Repo-Gal4, Mef2-Gal4, Cg-Gal4, Nsyb-Gal4, NP 2222, Nrv2-Gal4, NP6520 and Eaat-Gal4 were received from M. Koch (VIB, Leuven).
Flies were raised on standard cornmeal molasses fly food29 and kept at 25C in a 12 h light/dark cycle.
dCPT2δ18 and dCPT2δ3 flies were generated by imprecise excision of P(GSV7)GS21694, located 92 bp upstream from the transcriptional start site of CG2107. dCPT2δ18 is a 456 bp deletion of CG2107 exon 1. dCPT2δ3 is a 3224 bp deletion of the 5′ UTR and the first 505 amino acids of the CG2107 ORF. dCPT2Rev is a complete revertant and was used as control line. For targeted knockdown of dCPT2, UAS-line CG2107R2 from the NIG stock center was driven by Nsyb-Gal4 or Repo-Gal4.
Rescue constructs
Rat CPT2 cDNA was cloned into the EcoRI site of pUAST and checked for orientation. The H371A mutation was generated by site directed mutagenesis and the cDNA was recloned into pUAST.
cDNA was cloned into pH-Pelican with the mammalian Hsp70 promoter and UAS binding sites. All constructs were verified by sequencing the complete insert and introduced into the germline by standard P-element mediated transformation.
dCPT2-GFP knock-in
The knock-in of GFP has been performed by the fosmid tagging method of Ejsmont et al. 200930. In brief, fosmid pFlyFos 015048, a clone from a genomic fosmid library containing an AttB sequence recognized by φC31 integrase, the eye promoter–driven dominant selectable marker dsRed for isolation of transformants and a 40 kb genomic sequence flanking dCPT2 was used to C-terminally tag dCPT2 with super folder GFP by recombineering. The modified fosmid was verified by sequencing the tag-fosmid junction and injected into the germline of Drosophila melanogaster AttP2 flies. Revertant flies were selected with the help of dsRed driven in the adult eye.
Lipid droplet staining
Adult brains were dissected in ice-cold PBS, fixed in 4% paraformaldehyde for 15 min, washed 3× in PBS. Neutral Lipid Stain (HCS LipidTOX red neutral lipid stain, Molecular Probes, Eugene, Oregon) was added 1:1000 in PBS for 30 min light protected at room temperature, brains were 3× washed, mounted in PBS and analyzed directly in a Biorad Radiance confocal microscope at 594 nm.
qPCR
RNA was extracted from adult fly brains using Trizol. cDNA was synthesized with RevertAid First Strand cDNA Synthesis Kit using random primers. Quantitative PCR was performed with Light Cycler and SYBR green according to the manufacturer's protocol (Roche, Basel, Switzerland). Primers were designed to span an intron. Relative expression levels were quantified using the 2−ΔΔCt method.
act5c AGTCCGGCCCCTCCATT, CTGATCCTCTTGCCCAGACAA,
HNF4 CTGTCCAGATCCCCTTGTGT; GGCAGGATGAGCAGAATCTC,
SREBP AAGCTGAACAAGTCCGCAGT; CGCTTCTTGGATGCTCTACC,
PEPK GTGCCATCAACCCAGAGAAT; GCCCAACCAGTCAGTGATTT,
ACC CG11198 TAACAACGGAGTCACCCACA, CAGGTCACAACCGATGTACG,
HMG-CoA lyase TCAACCAATTGTCCGAAACA, AGCACTCTCGAATCCCTTCA,
HMG-CoA synthase TGGTGTCTCTGTTGATTAGCG, CTGATCGAGTACATGGAAGCAG, CG1140 CCTCATACTTTGGCTCCGATG, CCACCCATTCCCTTAACCAG,
CG13377 AGTGTGGTCGCTTTGGATAC, CCGCTCGATCACATGAAGAG,
Lethality
Homozygous larvae were collected and added to a new food vial at non-crowded density. The percentage of eclosed flies was assessed.
Elextron microscopy
Adult brains were fixed in 2.5% glutaraldehyde and 2% paraformaldehyde in 0.1 M Na-cacodylate buffer pH7.4 for 2 hours at room temperature and washed in 0.1 M Na-cacodylate buffer. Then, they were post-fixed in 1% osmiumtetroxide/H2O for 2 hours, dehydrated in a graded ethanol series and stained in uranyl acetate/70% ethanol. Finally, the samples were infiltrated with 1:1 mixture of epoxy resin (Agar100):propylene oxide for 60 min at 4°C and left overnight in 2:1 mixture agar:propylene oxide, embedded and polymerized for 2 days in 100% agar. Finally, 50–60 nm sections were cut with a Leica UCT ultramicrotome and post-stained with uranyl acetate for 10 min and lead citrate for 3.5 min. Brains were examined and photographed using a transmission electron microscope JEOL JEM-1400, equipped with a 2 k × 2 k Veleta (Olympus SIS, Tokyo, Japan) camera.
Western blot
Flies were killed by freezing, homogenized with 10 strokes of an Eppendorf pestle in a buffer containing 1% Triton X 100 and spun for 10 min at maximum speed in a table top centrifuge at 4C. The supernatant was diluted to 2 ug/ul protein content, boiled in SDS sample buffer, loaded on a Tris gel, transferred to a nitrocellulose membrane, blocked in non-fat dry milk and visualized with 1:1000 anti-CPT2 (H-300 from Santa Cruz), a rabbit polyclonal antibody raised against amino acids 51–350 of human CPT2, or 1:1000 anti-Histone 3 (#9715 from Cell Signaling) as loading control.
Life span
Flies were collected after birth and kept at 25°C in a humidified incubator with 12 hour light/dark cycle on regular corn molasses fly food. The vials were changed 3 times per week. Dead flies were scored every day.
Starvation
One week old flies (n > = 80) were kept on 1% agarose at 25°C in a humidified incubator with 12 hour light/dark cycle. Death was removed and scored every 8 hours.
Weight
The weight of 50 pooled one week old adult flies was quantified in a tared 1 ml Eppendorf tube on a balance (n > = 8).
Mobility
The motor activity of adult flies was quantified using a DAM activity monitoring system (Trikinetics, Waltham, MA).
Body length
The length of 10 adult, one week old flies per genotype was quantified under a Leica stereomicroscope.
Lipid analysis
Lipids were extracted from flies or brains with a modified Bligh & Dyer procedure31 and the extracts analyzed for phospholipids32, triacylglycerides33, fatty acids (NEFA kit; Wako, with some modifications) or separated by thin layer chromatography (silica 60 G plates; 0.25 mm; hexane/diethylether/acetic acid (70/30/1, v/v)), followed by transient visualization with iodine vapor, scraping of spots of interest and analysis. The non-extracted material was dissolved in 1% SDS and analyzed for protein content using bicinchoninic acid34.
Whole mount brain confocal imaging
Briefly, adult brains were hand dissected in ice-cold PBS and fixed for 10 min in 4% PFA, mounted in Vectashield on glass slides and subjected to confocal imaging in a Bio-Rad Radiance confocal microscope (Bio-Rad).
References
Van Veldhoven, P. P. Biochemistry and genetics of inherited disorders of peroxisomal fatty acid metabolism. J Lipid Res 51, 2863–95 (2010).
Bieber, L. L. Carnitine. Annu Rev Biochem 57, 261–83 (1988).
Bonnefont, J. P. et al. Carnitine palmitoyltransferase deficiencies. Mol Genet Metab 68, 424–40 (1999).
Wolfgang, M. J. et al. The brain-specific carnitine palmitoyltransferase-1c regulates energy homeostasis. Proc Natl Acad Sci U S A 103, 7282–7 (2006).
Hug, G., Bove, K. E. & Soukup, S. Lethal neonatal multiorgan deficiency of carnitine palmitoyltransferase II. N Engl J Med 325, 1862–4 (1991).
Taroni, F. et al. Molecular characterization of inherited carnitine palmitoyltransferase II deficiency. Proc Natl Acad Sci U S A 89, 8429–33 (1992).
North, K. N., Hoppel, C. L., De Girolami, U., Kozakewich, H. P. & Korson, M. S. Lethal neonatal deficiency of carnitine palmitoyltransferase II associated with dysgenesis of the brain and kidneys. J Pediatr 127, 414–20 (1995).
Pierce, M. R., Pridjian, G., Morrison, S. & Pickoff, A. S. Fatal carnitine palmitoyltransferase II deficiency in a newborn: new phenotypic features. Clin Pediatr (Phila) 38, 13–20 (1999).
Lavrentyev, E. N., Matta, S. G. & Cook, G. A. Expression of three carnitine palmitoyltransferase-I isoforms in 10 regions of the rat brain during feeding, fasting and diabetes. Biochem Biophys Res Commun 315, 174–8 (2004).
Clarke, D. & Sokoloff, L. Circulation and Energy Metabolism of the Brain. in Basic Neurochemistry (eds. Agranoff, B., Albers, R. W., Fisher, S. K. & Uhler, M. D.) (Lipincott-Raven, Philadelphia, 1999).
Nunnari, J. & Suomalainen, A. Mitochondria: in sickness and in health. Cell 148, 1145–59 (2012).
Brown, N. F., Anderson, R. C., Caplan, S. L., Foster, D. W. & McGarry, J. D. Catalytically important domains of rat carnitine palmitoyltransferase II as determined by site-directed mutagenesis and chemical modification. Evidence for a critical histidine residue. J Biol Chem 269, 19157–62 (1994).
Granneman, J. G., Moore, H. P., Mottillo, E. P. & Zhu, Z. Functional interactions between Mldp (LSDP5) and Abhd5 in the control of intracellular lipid accumulation. J Biol Chem 284, 3049–57 (2009).
Gronke, S. et al. Control of fat storage by a Drosophila PAT domain protein. Curr Biol 13, 603–6 (2003).
Edwards, T. N. & Meinertzhagen, I. A. The functional organisation of glia in the adult brain of Drosophila and other insects. Prog Neurobiol 90, 471–97 (2010).
Melom, J. E. & Littleton, J. T. Mutation of a NCKX eliminates glial microdomain calcium oscillations and enhances seizure susceptibility. J Neurosci 33, 1169–78 (2013).
Palanker, L., Tennessen, J. M., Lam, G. & Thummel, C. S. Drosophila HNF4 regulates lipid mobilization and beta-oxidation. Cell Metab 9, 228–39 (2009).
Auer, R. N., Olsson, Y. & Siesjo, B. K. Hypoglycemic brain injury in the rat. Correlation of density of brain damage with the EEG isoelectric time: a quantitative study. Diabetes 33, 1090–8 (1984).
Magistretti, P. J. Neuron-glia metabolic coupling and plasticity. J Exp Biol 209, 2304–11 (2006).
Du, L. et al. Starving neurons show sex difference in autophagy. J Biol Chem 284, 2383–96 (2009).
Guzman, M. & Blazquez, C. Is there an astrocyte-neuron ketone body shuttle? Trends Endocrinol Metab 12, 169–73 (2001).
Edmond, J., Robbins, R. A., Bergstrom, J. D., Cole, R. A. & de Vellis, J. Capacity for substrate utilization in oxidative metabolism by neurons, astrocytes and oligodendrocytes from developing brain in primary culture. J Neurosci Res 18, 551–61 (1987).
Rajan, A. & Perrimon, N. Drosophila as a model for interorgan communication: lessons from studies on energy homeostasis. Dev Cell 21, 29–31 (2011).
Haslam, D. W. & James, W. P. Obesity. Lancet 366, 1197–209 (2005).
Cheng, L. Y. et al. Anaplastic lymphoma kinase spares organ growth during nutrient restriction in Drosophila. Cell 146, 435–47 (2011).
Karmi, A. et al. Increased brain fatty acid uptake in metabolic syndrome. Diabetes 59, 2171–7 (2010).
Berchtold, N. C. et al. Gene expression changes in the course of normal brain aging are sexually dimorphic. Proc Natl Acad Sci U S A 105, 15605–10 (2008).
Loerch, P. M. et al. Evolution of the aging brain transcriptome and synaptic regulation. PLoS One 3, e3329 (2008).
Schulz, J. G. et al. Drosophila syndecan regulates tracheal cell migration by stabilizing Robo levels. EMBO Rep 12, 1039–46 (2011).
Ejsmont, R. K., Sarov, M., Winkler, S., Lipinski, K. A. & Tomancak, P. A toolkit for high-throughput, cross-species gene engineering in Drosophila. Nat Methods 6, 435–7 (2009).
Van Veldhoven, P. P. & Bell, R. M. Effect of harvesting methods, growth conditions and growth phase on diacylglycerol levels in cultured human adherent cells. Biochim Biophys Acta 959, 185–96 (1988).
Van Veldhoven, P. P. & Mannaerts, G. P. Inorganic and organic phosphate measurements in the nanomolar range. Anal Biochem 161, 45–8 (1987).
Van Veldhoven, P. P., Swinnen, J. V., Esquenet, M. & Verhoeven, G. Lipase-based quantitation of triacylglycerols in cellular lipid extracts: requirement for presence of detergent and prior separation by thin-layer chromatography. Lipids 32, 1297–300 (1997).
Smith, P. K. et al. Measurement of protein using bicinchoninic acid. Anal Biochem 150, 76–85 (1985).
Acknowledgements
We thank Pedro F. Marrero (U Barcelona) for cDNA rCPT2, P. Tomancaz for pFlyFos 015048, M. Koch (VIB, Leuven), Bloomington and NIG stock centre and R. Kühnlein for stocks, T. Holmes for help with the DAM system, Pieter Baatsen (EM-facility EMoNe, VIB Bio Imaging Core/KU Leuven) for help with electron microscopy, R. Ejsmont for help with dCPT2 GFP knock-in, Guy Froyen (KU Leuven) for help with qPCR, Evelyne de Schryver (KU Leuven) for help with lipid analysis and fatty acid oxidation experiments. This work was partially supported by the Flanders Fund for Scientific Research (FWO G 0.666.10N), NEUROBRAINNET IAP 7/16, Flemish Government Methusalem Grant, Spanish Ministry of Science (SAF2010-14906) and Innovation Ingenio-Consolider (CSD2010-00045) and Spanish Ministry of Economy and Competitiveness (SAF2013-45392).
Author information
Authors and Affiliations
Contributions
J.G.S., A.L., L.V.H., A.G., S.V., J.B. and P.P.V.V. performed the experiments, J.G.S. and C.G.D. supervised the project.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary figure 1 as pdf
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Schulz, J., Laranjeira, A., Van Huffel, L. et al. Glial β-Oxidation regulates Drosophila Energy Metabolism. Sci Rep 5, 7805 (2015). https://doi.org/10.1038/srep07805
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep07805
This article is cited by
-
A neuron–glia lipid metabolic cycle couples daily sleep to mitochondrial homeostasis
Nature Neuroscience (2024)
-
Glial lipid droplets resolve ROS during sleep
Nature Neuroscience (2024)
-
Exploring the molecular makeup of support cells in insect camera eyes
BMC Genomics (2023)
-
Glycolytically impaired Drosophila glial cells fuel neural metabolism via β-oxidation
Nature Communications (2023)
-
Fatty acid oxidation organizes mitochondrial supercomplexes to sustain astrocytic ROS and cognition
Nature Metabolism (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
