
Abstract
The leaves of many plants are red during particular stages of their lives, but the adaptive significance of leaf colouration is not yet clearly understood. In order to reveal whether anthocyanins play a similar role (i.e. antioxidants) in different seasonal contexts, this study investigated species with red young leaves in the subtropical forest of Dinghushan biosphere reserve (South China) during summer and winter and compared group leaf characteristics between the two seasons. Of 62 total species, 33 exhibited red young leaves in summer only, 6 in winter only, and 23 in both seasons. The anthocyanins extracted from most of these species had an absorption peak at ~530 nm. Frequency distribution analysis showed that the species containing anthocyanins at levels ranging from 0.02 to 0.04 μmol cm−2 occurred most frequently in summer or winter. Based on conditional grouping of the species, no significant variation was observed in the average anthocyanin contents and antioxidant abilities between summer and winter; the flavonoid content in summer was 2-fold that in winter, whereas the anthocyanin:flavonoid ratio in summer was only half that in winter. Moreover, a positive correlation between anthocyanins and flavonoids was found in summer. Therefore, it is less likely for anthocyanins to serve as antioxidants in summer than winter, because such a function in summer leaves is readily replaced by other flavonoids.
Similar content being viewed by others


Introduction
In tropical and subtropical evergreen broad-leaved forests, the conspicuous red to purple pigmentation of young leaves due to the presence of anthocyanins can be observed during all seasons, including winter1. This developmental pattern is extremely common, and according to previous investigations conducted in tropical forests, 7% to 62% of the total vascular flora in a single site exhibited red pigmentation in the young leaves2. Red pigmentation is present in a variety of unrelated plant families and can be found in the vacuoles of different leaf cell layers or features, including the adaxial/abaxial epidermis, palisade mesophyll, spongy mesophyll, bundle sheath cells and trichomes3,4. Red pigmentation is influenced by one or more stress conditions, such as strong light5, UV-B radiation6, low temperatures7, low pH8, drought9, nutrient deficiencies10, pathogen infections and herbivory11. In nature, anthocyanins are present mainly in the form of hybrid heterosides, and their compositions can change in response to different stresses even in the same species12.
The biological functions of anthocyanins in leaves are as varied as their cellular location and environmental stimuli. Current research has shown that the anthocyanins that accumulate in leaves can act as ROS scavengers13, light screens14, metal chelators15, and antimicrobial agents16. Anthocyanins may also signal the presence of chemical defences17 or function in crypsis to herbivores18. Recently, an increasing number of documents showed that anthocyanins participate in photoprotection by screening the excessive light energy. In general, anthocyanins absorb visible light with a peak in the 500–550 nm waveband19. Thus, anthocyanins can serve a photoprotective function by absorbing excessive blue-green light that could otherwise be absorbed by chloroplasts in the subjacent mesophyll20. The light energy absorbed by anthocyanins is eventually dissipated into heat through a series of ultra-fast excited-state processes21. In addition to absorbing blue-green light, anthocyanins can also scavenge free radicals. However, as anthocyanins constitute only a minor portion of the total antioxidant pool in leaves, whether they protect plants from oxidative stress by acting as scavengers of free radicals remains a subject of debate. Anthocyanins belong to the group of compounds known as flavonoids, and most flavonoids are powerful antioxidants because of their strong capacity to donate electrons or hydrogen atoms22. Therefore, analysing the relationships between the accumulation of anthocyanins, flavonoids and antioxidants in specific groups of species and in a specific site may increase our understanding of whether it is necessary for anthocyanins to function as antioxidants.
Dinghushan biosphere reserve (DHSBR) is the first natural reserve in China and is located near the Tropic of Cancer. Two-thirds of the global area near this latitude consists of deserts and semideserts. However, the DHSBR is covered by a subtropical evergreen broad-leaved forest, and the climax community in the area is more than 400 years old23. Because of its unique geographical location and the presence of native vegetation with little human interference, the DHSBR has become one of the most valuable areas for scientific research. Like many other evergreen broad-leaved forest, the subtropical forest has abundant plants with coloured leaves. To better understand whether anthocyanins play an antioxidant role in the plants in DHSBR area, species with red young leaves during summer and winter were investigated, and the absorption spectra of anthocyanins, the relationships among anthocyanin contents, flavonoid contents and antioxidant abilities in different groups of species were explored during the two contrasting seasons.
Results
Species with red young leaves in the DHSBR in summer and winter
The DHSBR forest was rich in plant species with red young leaves, most of which were woody species (Table 1; Fig. S1 available as Additional Information). A total of 62 different species were observed with red (with anthocyanin) young leaves in the two field investigations, and species examined consisted of 38 trees, 11 shrubs, 12 vines and 1 herb. Among the species, Lindera chunii and Macaranga sampsonii are endemic plants in the DHSBR. The number of species (56) with red young leaves was greater in summer than in winter (29). There were 33 and 6 species with red young leaves only in summer or winter, respectively. Twenty-three species had red young leaves in both seasons, of which 5 species (Acmena acuminatissima, Ficus fistulosa, Machilus chinensis, Blastus cochinchinensis, Eriocarpous Glochidion and Rourea microphylla) were visibly redder in winter than in summer, and 2 species (Craibiodendron kwangtungense and Erycibe schmidtii) were the opposite. We noted that the young leaves of some of the species were not really red; for example, those of Xanthophyllum hainanense and Smilax china were blue and purple-black, respectively. In addition to showing temporal diversity, there was also diversity in the distribution of anthocyanin pigments in the tissues of the plants in the DHSBR. For most of the species, there was pigmentation on both the adaxial and abaxial sides of the young leaves. However, for a few species, there was pigmentation only on the adaxial side (such as Eurya groffii and E. Glochidion), on the abaxial side (such as Melastoma sanguineum), or in trichome (such as Castanopsis fissa and Schefflera octophylla).
Absorption spectra and anthocyanin contents of the different species
Leaf samples of 44 species and 22 species were collected in the DHSBR during summer and winter, respectively, and the absorption spectra of their anthocyanin extracts are shown in Fig. 1. As seen in Fig. 1, 40 out of 44 species in summer (Fig. 1(a2–c4)) and 20 out of 22 species in winter (Fig. 1(d1–e4)) had absorption peaks at 530 ± 5 nm, which was consistent with the anthocyanin standard, cyanidin-3-O-glucoside (Fig. 1(a1)). C. fissa, S. china, C. kwangtungense and Sapium discolor accumulated the most anthocyanins in summer, and R. microphylla, E. Glochidion, Rhaphiolepis indica and Syzygium rehderianum accumulated the most in winter (Figs. 1 and 2). Unlike most species, the absorption peaks of E. schmidtii, Pteris vittata and X. hainanense in summer occurred at approximately 496 nm rather than 530 nm (Figs 1 and 2). Among the three species, only E. schmidtii was found to have young red leaves in both summer and winter, and the sample collected in winter showed an absorption spectrum that was identical to that in summer. Ixora chinensis was the only species that did not show an evident absorption peak in its spectrum either in summer or winter. With wavelengths ranging from 600 to 450 nm, the absorption spectra of Ixora chinensis showed a distinctive increasing trend.

Absorption spectra of the anthocyanin extracts from the red young leaves of 44 and 22 species in the Dinghushan biosphere reserve in summer (panels a2-c4) and winter (panels d1-e4), respectively. Panel a1 is the anthocyanin standard cyanidin-3-O-glucoside. Vertical dotted lines indicate a wavelength at 530 nm. According to visually-similar absorption peaks and the absorbance at 530 nm from high to low, every 4 species formed a group in summer and every 2 or 3 species formed a group in winter.

Some outstanding species in the Dinghushan biosphere reserve. The upper and middle 4 panels were the top 4 species with the highest accumulated anthocyanins in their young leaves in summer and winter, respectively. The species in lower panels exhibited absorption spectra that are different from most other plants with red leaves. (Photo by T.-J. Zhang).
The anthocyanin content of the species with an absorption peak at approximately 530 nm was calculated using cyanidin-3-O-glucoside as the standard. The frequency distributions of 40 species in summer and 19 species in winter based on anthocyanin content are shown in Fig. 3a. The anthocyanin contents that occurred most frequently ranged from 0.02–0.04 μmol cm−2, with 17 species in summer and 7 species in winter. In order to facilitate the comparisons of anthocyanins and other characteristics between summer and winter, the species were classified: those in summer and winter with anthocyanin contents >0.02 μmol cm−2 refer as SI and WI groups, respectively; for those with red young leaves in both the two seasons, they refer as SII and WII groups when in summer and winter, respectively. The statistical results showed that there were no significant variation in the mean anthocyanin contents between the SI and WI groups or between the SII and WII groups (Fig. 3b).

(a) Frequency histogram of the species whose young leaves had anthocyanin contents >0.005 μmol cm−2 in summer (Sum) and winter (Win). (b) Average anthocyanin content in the young leaves of the 4 species groups (SI, WI, SII, and WII) in the two seasons. Statistical significant differences are indicated: ns, non-significant; *P < 0.05; **P < 0.01; ***P < 0.001.
Chlorophyll (Chl) content in the different species groups
The mean Chl content was higher in the young leaves of the SI group than of the WI group (P < 0.05), while the mean Car content was not significantly different between the two groups (Fig. 4). As a result, the Car:Chl ratio in the young leaves of the SI group was significantly lower than that of the WI group (P < 0.05). Similar changes in Chl content, Car content and Car:Chl ratio were found in the mature leaves between the SI and WI groups and in the young and mature leaves between the SII and WII groups.

Chlorophyll and carotenoid content in the young red (Y) and mature leaves (M) of the 4 species groups (SI, WI, SII, and WII) in the two seasons. Statistical significant differences are indicated: ns, non-significant; *P < 0.05; **P < 0.01; ***P < 0.001. The x-axis labels correspond to the group leaf type, e.g., SI_Y represent young leaves in the SI group.
Antioxidant abilities and flavonoid contents in the different species groups
There were no significant differences in the average antioxidant abilities in the young or mature leaves between the SI and WI groups (Fig. 5a,b). In contrast, the average flavonoid content was 1-fold greater in the young or mature leaves of the SI group than in the WI group (Fig. 5c) and 1.8–2.2-fold greater in the young or mature leaves of the SII group than in the WII group (Fig. 5d). The effects of different seasons on the accumulation of flavonoids in the young leaves of the species groups were apparently different from those of the anthocyanins. The summer groups accumulated more flavonoids than the winter groups, and therefore, the differences in the flavonoids:antioxidant abilities (Flav:AA) ratios between either the SI and WI groups or the SII and WII groups were greater than those in the anthocyanins:antioxidant abilities (Antho:AA) ratios (Fig. 6a,b). Moreover, the Antho:Flav ratios in the young leaves of the SI and SII groups were only half those of the WI and WII groups (Fig. 6c). Instead, the SI and SII groups had the Flav:Chl ratios that were slightly higher than the corresponding winter groups (Fig. 6d).

Antioxidant abilities and total flavonoid contents in the young red (Y) and mature leaves (M) of the 4 species groups (SI, WI, SII, and WII) in the two seasons. Statistical significant differences are indicated: ns, non-significant; *P < 0.05; **P < 0.01; ***P < 0.001. The x-axis labels correspond to the group leaf type, e.g., SI_Y represents young leaves in the SI group.

Anthocyanins:antioxidant abilities (Antho:AA), anthocyanins:flavonoids (Antho:Flav), flavonoids:antioxidant abilities (Flav:AA) and flavonoids:chlorophylls (Flav:Chl) in the young red leaves of the 4 species groups (SI, WI, SII, and WII) in the two seasons. Statistical significant differences are indicated: ns, non-significant; *P < 0.05; **P < 0.01; ***P < 0.001.
Correlations between anthocyanins and flavonoids
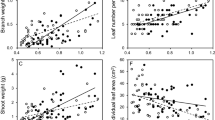
Anthocyanins represent a portion of the flavonoid pool and antioxidant ability in the red leaves. However, there was no linear relationship between the anthocyanins and the antioxidant abilities or between the flavonoids and the antioxidant abilities in the young leaves of any of SI, WI, SII, and WII groups (Figs. 7 and 8). Anthocyanins were positively correlated with flavonoids in the SI group (P < 0.05) and SII group (P < 0.01) but not in the WI and WII groups (Fig. 7c,d). According the distribution of antioxidant abilities versus flavonoids (Fig. 8a), the species in SI group tended to be separated into three subgroups (indicated by the dotted oval). One was characterized by high antioxidant abilities and low flavonoids, including 8 species (e.g. Syzygium jambos, Alchornea trewioides and Syzygium rehderianum). Another included the most species (e.g. Rubus alceaefolius, Rhaphiolepis indica and Microcos paniculata), in which the antioxidant abilities were associated with flavonoids. The third had two species (Craibiodendron kwangtungense and Desmos chinensis), which was characterized by low antioxidant abilities and high flavonoids.

Correlations between the anthocyanin contents and the antioxidant abilities and flavonoid contents in the young leaves in the 4 species groups (SI, WI, SII, and WII) in two seasons. A significant correlation was only found in SI and SII in the two lower panels, and that the best fit lines only correspond with those data.

Correlations between the antioxidant abilities and flavonoid contents in the young leaves in the 4 species groups (SI, WI, SII, and WII) in two seasons.
Discussion
Species with red young leaves
Some species in the DHSBR continuously produce new leaves all year round, other produce them only during specific seasons. This may represent different kinds of life history strategies. But the pigmentation pattern of young leaves is not directly related to any specific life history strategy, because there are always some species with young red leaves in the DHSBR all year round. Our investigations found that 62 different species produced new leaves that are red in the DHSBR during summer and winter. Since our investigations were carried out for a limited period of time and in a limited area, the result was unable to cover all plant species with young red leaves in the DHSBR. For example, a previous investigation has demonstrated that the young leaves of Lindera communis, Litsea verticillata, Rhododendron henryi and Elaeocarpus chinensis in the forest were also red24. So, on a conservative estimate, there may be approximately 80 species with red young leaves in the DHSBR, of which 70 are woody species. Based on the investigation of a 20 ha plot, there are 210 woody species in the DHSBR25. Therefore, woody species with red young leaves account for a third of the total woody species in the subtropical forest, which included nearly all the dominant species.
Absorbance profiles of anthocyanins in the young red leaves
Absorption of the anthocyanin extracts varied in different species may due to different compositions of the pigments. For 90% of the species investigated in this study, the absorption peaks occurred at 530 ± 5 nm, which are roughly consistent with consistent with cyanidin-3-O-glucoside (Fig. 1), a common anthocyanin in pant vegetative organs. Though cyanidin-3-O-glucoside is not the only anthocyanin that absorbs at 530 nm26, it is a useful standard for estimation of anthocyanin contents in most of the species. To understand the exact compositions of anthocyanins in the various species, much more work is needed. Nevertheless, cyanidin derivatives (not necessarily cyanidin-3-O-glucoside) have been extensively identified in the vegetative tissues of many plants27,28,29. So, it is likely that the red pigments accumulated in the species with maximum absorption at about 530 nm consisted mainly of cyanidin derivatives. A few species showed absorption peaks at approximately 496 nm (including Erycibe schmidtii, Pteris vittata and X. hainanense) or did not even have an evident absorption peak (i.e., Ixora chinensis), suggesting that compositions of anthocyanins in these species are very different from most of other species. As the pigments with an absorption peak at ~496 nm is consistent with that of luteolinidin-5-O-glucoside30, they might contain lots of luteolinidin derivatives rather than cyanidin derivatives.
Effects of seasonal variations on group leaf characteristics
Temperatures, irradiances and precipitations are different between summer and winter in the DHSBR area. In summer, growth and photosynthetic function in leaves are readily influenced by high temperatures and high irradiances. As the season changes, chilling temperatures and drought in winter become the major factors that inhibit the growth of leaves and their functions. The variations of the Chl and flavonoid contents indicate that the species in the DHSBR took different strategies to adapt summer winter stress. Reduced Chl content in the leaves of winter groups resulted in a higher a Car:Chl ratio (Fig. 4). Since winter is drier and colder than summer, desiccation and low-temperature stress in plants should be relatively higher, which would be accompanied by stomatal closure and photoinhibition of photosynthesis. As some specific carotenoids, such as xanthophylls, are involved in the mechanisms to cope with the absorption of excess light energy in photosynthetic apparatus, high Car:Chl ratio can imply that the capacity of photoprotection was enhanced in the leaves31,32. This may allow the species to adapt to winter stresses.
In contrary to the Car:Chl ratio, the flavonoids were dramatically higher in the summer leaves than the winter ones (Fig. 5). Flavonoids can be involved in photoprotection by acting as antioxidants or light attenuators. Thus, the upregulation of flavonoids in plants has been observed under various environmental stresses, such as high light33,34,35 and chilling temperatures36,37,38. Though flavonoids didn’t have a remarked effect on total antioxidant abilities either in the summer or winter leaves, the higher flavonoids could make a greater contribution to the antioxidant abilities in summer leaves. Since there was no difference in the antioxidant abilities between summer and winter leaves, flavonoids in the summer leaves may substitute some other antioxidants. Perhaps, flavonoids are better kind of antioxidants for the plants to adapt summer stresses. In addition, winter leaves accumulated less flavonoids is likely due to the coordination of different photoprotective mechanisms but not the inhibition of environmental conditions, because they still had the ability to produce large amounts of anthocyanins.
Unlike the changes of Chls and flavonoids, the anthocyanins tended to maintain at the same level between the summer and winter groups of species (Fig. 3). The anthocyanin concentrations observed in the two seasons may be adequate for achieving their functions in either season. In summer, the anthocyanins were positively correlated with flavonoids (Fig. 7), suggesting that the accumulation of anthocyanins can be partly attributed to the active metabolism of flavonoids. In other words, some anthocyanins in the young leaves may be “by-products” of flavonoid metabolism in summer. In winter, however, the production of anthocyanins were independent of other flavonoids, consistent with previous studies showing that the anthocyanins increased in some plants under environmental stresses without affecting the flavonoids5,39. The larger amount of falovanoids observed in summer may be caused by light-induced activation of enzymes for falovanoid pathway. As summer conditions are more beneficial than winter conditions for photosynthesis of plants, maybe there are also more available energy for the synthesis of flavonoids in summer. Nevertheless, as summer leaves had high flavonoids, the anthocyanins acting as scavengers of free radicals can be easily replaced by other flavonoids. In this respect, the photoprotective function afforded by anthocyanins in young leaves might be less important in summer than in winter. As summer temperatures are more favourable for insect herbivores than winter ones40, it may be more important for anthocyanins to play an anti-herbivory role in summer than in winter.
Though the difference of mean antioxidant abilities between the summer and winter groups of species was consistent with that of mean anthocyanin concentrations, a low correlation was observed between the anthocyanins and the antioxidant abilities. This might be partly caused by the fact that while anthocyanins are mainly located certain cells within the leaves, other antioxidative compounds are located in the whole profile of the leaf tissue. Future synchronous analyses of anthocyanins, flavonoids and antioxidant potentials in specific leaf tissues in situ may provide more accurate information for us to learn about the antioxidation effect of anthocyanins in different seasonal conditions. Moreover, future investigations of the leaf physiological characteristics in the different species groups, which are divided by anthocyanin distribution features (e.g. in epidermis or trichome) or antioxidant traits (e.g. high antioxidant abilities and low flavonoids or low antioxidant abilities and high flavonoids), will also provide insights into the adaptive significances of the anthocyanins accumulated in the young leaves of a variety of species in subtropical forests.
Materials and Methods
Investigation location and sampling
The DHSBR is located at latitude 23°09′21″–23°11′30″N and longitude 112°30′39″–112°33′41″E in Zhaoqing, Guangdong Province, China, approximately 80 km west of Guangzhou. It has an area of 1154 hm2 and a hilly terrain, with altitudes ranging between 10 and 1000 m. The geology of this area consists of sandstone and shale bedrock formed by Devonian-aged sediments overlain with lateritic red soil. Although the DHSBR is located near the Tropic of Cancer, the area has a subtropical monsoon humid climate due to the effect of the Pacific and Indian Oceans. The mean annual temperature is 21 °C, and the temperatures in summer and winter are on average 28.0 °C and 12.0 °C, respectively. Occasionally, they can reach to 38 °C in summer and −2 °C in winter41. The annual rainfall ranges from 1560 to 2278 mm, with a mean of 1900 mm, of which approximately 80% occurs from April to September. The DHSBR has 1956 species of higher plants representing 877 genera from 260 families, of which 210 species of woody plants belong to 119 genera and 56 families25. The investigation of plants with red young leaves in the DHSBR was performed in January (winter) and August (summer) 2016 and mainly focused on 3 areas (A1–A3, Fig. 9). Investigation areas A1, A2 and A3 represented a ravine evergreen broad-leaved forest (altitude 50–90 m), a monsoon evergreen broad-leaved secondary forest (altitude 270–320 m), and a monsoon evergreen broad-leaved forest (altitude 160–210 m), respectively. In particular, A3 was located in the climax community in the DHSBR that is more than 400 years old. Plants with red young leaves were sought along the mountain paths within these areas.

Investigation area (A1, A2 and A3) in the Dinghushan biosphere reserve. Map was drawn using ArcGIS 10.2 software (http://www.esri.com/).
To collect samples for the determination of anthocyanin and other physiological traits in the red leaves, a pair of common branch scissors or a pair of 10 m high branch scissors was used to cut down a branch of a species with red leaves. The branch was immediately placed in a plastic bucket with the cut end in water. After returning indoors, 15–20 leaf discs (10 mm in diameter for most species; 8 mm for Rourea microphylla due to its small leaves) were cut off from the most striking red young leaves and fully expanded mature leaves of each species, packed in tinfoil (5 leaf discs per package), and stored in liquid nitrogen until further processing. Leaf samples were collected from 2–3 individuals of each species with red leaves.
Determination of anthocyanin content
Anthocyanins were extracted from the red leaves in methanol containing 1% HCl. Four leaf discs were immersed in 4 mL of the HCl methanol solution in a 15 mL test tube with screw caps and placed at 4 °C in the dark for approximately 24 h until the leaf colour was completely bleached. Both anthocyanins and damaged chlorophylls (Chls) were dissolved in the extract, which made the extract appear cloudy. To separate anthocyanin from the extract, 4 mL of chloroform and 2 mL of distilled water were added and mixed. After standing for 3 minutes, the anthocyanins and Chls were dissolved in the upper water-methanol phase and the lower chloroform phase, respectively. The volume of the water-methanol phase was measured and recorded. Then, a 3 mL aliquot of the upper anthocyanin extract was transferred to a cuvette, and the absorption spectra were scanned at 1 nm wavelength intervals from 400 to 700 nm using a UV-Vis 2450 spectrophotometer (Shimadzu, Japan). The absorption peaks of the anthocyanin extract for most species occurred at approximately 530 nm, which is consistent with cyanidin-3-O-glucoside. Therefore, the anthocyanin content in these species was calculated using the absorbance at 530 nm, and a calibration curve was constructed with 5–100 μmol L−1 of cyanidin-3-O-glucoside (Sigma) and expressed on an area basis as μmol cm−2. A few species, such as Erycibe schmidtii, Pteris vittata and Xanthophyllum hainanense, had an absorption peak at 496 nm rather than 530 nm. The anthocyanin contents of these species were not calculated due to a lack of standards.
Determination of the chlorophyll (Chl) content
Chls were extracted in 80% acetone. For every sample, 3 leaf discs were immersed in 4 mL of 80% acetone in a 10 mL test tube, and the screw cap was then tightened. The samples were placed at 4 °C in the dark and shaken at 8 h intervals until the leaf discs were bleached. The Chls were successfully extracted from most species using this immersion method and yielded clear green solutions. However, this method was not applicable for several species, such as Brassaiopsis glomerulata and Pteris vittata. The leaf discs of these two species yielded a turbid brown solution when immersed in 80% acetone, indicating that the Chls were damaged due to unknown reasons. The total Chls and carotenoids (Cars) in the clear green extract were spectrophotometrically measured according to Wellburn42 and expressed on an area basis.
Determination of the flavonoid content and antioxidant ability
The flavonoids and antioxidants were extracted from two leaf discs of each sample in 1.5 mL of methanol in a sealed centrifuge tube at 4 °C in the dark for 48 h. The flavonoid content in each extract was determined by aluminium chloride spectrophotometry43. Briefly, 0.5 mL of 10-fold diluted extract was mixed with 0.2 mL of 5% NaNO2, 0.3 mL of 10% AlCl3 (freshly prepared) and 1 mL of 1 M NaOH (freshly prepared). After standing for 5 minutes, the absorbance of the mixture was measured at 510 nm using another mixture made with deionized water instead of the flavonoid extract as a blank. The flavonoid content was calculated using a calibration curve constructed with 25–1000 μmol L−1 of catechin and expressed on an area basis.
The antioxidant ability in the extract was measured by a 1,1-diphenyl-2-picrylhydrazyl (DPPH) assay according to Nguyen et al.44 A 10 μL aliquot of the extract was added to 3 mL of 100 μM freshly prepared DPPH solution (in 95% methanol). After standing for 5 minutes, the absorbance of the mixture was measured at 517 nm using 95% methanol as a blank. The clearance rate of DPPH was calculated by a calibration curve made with 20–100 μM of DPPH. The antioxidant ability was expressed as μmol of DPPH scavenging capacity per unit area.
Statistical analysis
All statistical procedures were performed using IBM SPSS Statistics v. 19.0 (IBM, NY, USA). A Student’s t test was used to examine the effects of season (winter, summer) on the anthocyanin, Chl and flavonoid contents and the antioxidant abilities based on conditional grouping of the species. Since the concentration-frequency distribution showed that the species with an anthocyanin content ranging from 0.02–0.04 μmol cm−2 occurred most frequently in both summer and winter, the first method for grouping the species was based on the condition that the anthocyanin content was >0.02 μmol cm−2. As a result, 29 species (Castanopsis fissa, Craibiodendron kwangtungense, Smilax china, Sapium discolor, Desmos chinensis, Rubus alceaefolius, Rhaphiolepis indica, Microcos paniculata, Tetracera asiatica, Toxicodendron vernicifluum, Macaranga sampsonii, Sabia limoniacea, Engelhardtia, roxburghiana, Syzygium jambos, Rourea microphylla, Cryptocarya concinna, Cratoxylum cochinchinense, Alchornea trewioides, Bauhinia championi, Machilus pingii, Uncaria rhynchophylla, Syzygium rehderianum, Brassaiopsis glomerulata, Fissistigma glaucescens, Castanopsis chinensis, Marsdenia tinctoria, Aporosa dioica, Eriocarpous Glochidion, and Homalium cochinchinense) were in the summer group (SI), and 14 species (R. microphylla, R. indica, S. rehderianum, E. Glochidion, Ficus fistulosa, E. roxburghiana, C. kwangtungense, Calophyllum membranaceum, Eurya groffii, Blastus cochinchinensis, Ficus esquiroliana, S. jambos, A. dioica, and Acmena acuminatissima) were in the winter group (WI). The second grouping method focused only on species that had red young leaves in both summer and winter, namely the intersection of the two seasons. Ten species (B. cochinchinensis, S. rehderianum, C. kwangtungense, R. microphylla, E. roxburghiana, Garcinia oblongifolia, E. Glochidion, R. indica, A. dioica, and S. jambos) met the criterion, and the corresponding summer and winter groups were designated SII and WII, respectively. The correlations between anthocyanins and the antioxidants and flavonoids in various species groups were analysed using Sigmaplot v. 12.5 (Systat Software Inc., USA).
References
Hughes, N. M. Winter leaf reddening in ‘evergreen’ species. New Phytol. 190, 573–581 (2011).
Dominy, N. J. et al. Why are young leaves red? Oikos 98, 163–176 (2002).
Lee, D. W. Anthocyanins in leaves: distribution, phylogeny and development. Adv. Bot. Res. 37, 37–53 (2002).
Zhang, T.-J. et al. A magic red coat on the surface of young leaves: anthocyanins distributed in trichome layer protect Castanopsis fissa leaves from photoinhibition. Tree Physiol. 36, 1296–1306 (2016).
Hoffmann, A. M., Noga, G. & Hunsche, M. Alternating high and low intensity of blue light affects PSII photochemistry and raises the contents of carotenoids and anthocyanins in pepper leaves. Plant Growth Regul. 79, 275–285 (2016).
Mendez, M., Jones, D. G. & Manetas, Y. Enhanced UV-B radiation under field conditions increases anthocyanin and reduces the risk of photoinhibition but does not affect growth in the carnivorous plant Pinguicula vulgaris. New Phytol. 144, 275–282 (1999).
Hughes, N. M., Neufeld, H. S. & Burkey, K. O. Functional role of anthocyanins in high-light winter leaves of the evergreen herb Galax urceolata. New Phytol. 168, 575–587 (2005).
Zhang, Y. et al. Low medium pH value enhances anthocyanin accumulation in Malus crabapple leaves. PloS One 9, e97904 (2014).
González-Villagra, J., Cohen, J. D. & Reyes-Díaz, M. M. Abscisic acid is involved in phenolic compounds biosynthesis, mainly anthocyanins, in leaves of Aristotelia chilensis plants (Mol.) subjected to drought stress. Physiol. Plant 165, 855–866 (2019).
Lea, U. S., Slimestad, R., Smedvig, P. & Lillo, C. Nitrogen deficiency enhances expression of specific MYB and bHLH transcription factors and accumulation of end products in the flavonoid pathway. Planta 225, 1245–1253 (2007).
Hatier, J. -H. B. & Gould, K. S. Anthocyanin function in vegetative organs. In Gould, K., Davies, K. M. & Winefield, C. Eds Anthocyanins: biosynthesis, functions, and applications. New York: Springer, 1–19 (2008).
Kovinich, N. et al. Not all anthocyanins are born equal: distinct patterns induced by stress in Arabidopsis. Planta 240, 931–940 (2014).
Kytridis, V.-P. & Manetas, Y. Mesophyll versus epidermal anthocyanins as potential in vivo antioxidants: evidence linking the putative antioxidant role to the proximity of oxy-radical source. J. Exp. Bot. 57, 2203–2210 (2006).
Nichelmann, L. & Bilger, W. Quantification of light screening by anthocyanins in leaves of Berberis thunbergii. Planta 246, 1069–1082 (2017).
Landi, M., Tattini, M. & Gould, K. S. Multiple functional roles of anthocyanins in plant-environment interactions. Environ. Exp. Bot. 119, 4–17 (2015).
Coley, P. D. & Aide, T. M. Red coloration of tropical young leaves: a possible antifungal defence? J. Trop. Ecol. 5, 293–300 (1989).
Cooney, L. J. Red leaf margins indicate increased polygodial content and function as visual signals to reduce herbivory in Pseudowintera colorata. New Phytol. 194, 488–497 (2012).
Schaefer, H. M. & Wilkinson, D. M. Red leaves, insects and coevolution: a red herring? Trends Ecol. Evol. 19, 616–618 (2004).
Li, D., Wang, P., Luo, Y., Zhao, M. & Chen, F. Health benefits of anthocyanins and molecular mechanisms: Update from recent decade. Crit. Rev. Food Sci. Nutr. 57, 1729–1741 (2017).
Zhang, K.-M. et al. Photoprotective roles of anthocyanins in Begonia semperflorens. Plant Sci. 179, 202–208 (2010).
Costa, D. et al. Photochemistry of the hemiketal form of anthocyanins and its potential role in plant protection from UV-B radiation. Tetrahedron 71, 3157–3162 (2015).
Hernández, I., Alegre, L., Van Breusegem, F. & Munné-Bosch, S. How relevant are flavonoids as antioxidants in plants? Trends Plant Sci. 14, 125–132 (2009).
Shen, C. et al. 14C measurement of forest soils in Dinghushan Biosphere Reserve. Chin. Sci. Bull. 44, 251–256 (1999).
Zhu, H. Photoprotection strategies of young leaves from dominant tree species in mid and late successional stages of a lower subtropical forest community. PhD Thesis, South China Normal University, China (2013).
Ye, W. et al. Community structure of a 20 hm2 lower subtropical evergreen broadleaved forest plot in Dinghushan, China. Acta Phytoecol. Sin. 32, 274–286 (2008).
Harborne, J. B. Spectral methods of characterizing anthocyanins. Biochem. J. 70, 22–28 (1958).
Close, D. C., Davies, N. W. & Beadle, C. L. Temporal variation of tannins (galloylglucoses), flavonols and anthocyanins in leaves of Eucalyptus nitens seedlings: implications for light attenuation and antioxidant activities. Funct. Plant Bio. 28, 269–278 (2001).
Islam, M. S., Yoshimoto, M., Terahara, N. & Yamakawa, O. Anthocyanin compositions in sweetpotato (Ipomoea batatas L.) leaves. Biosci. Biotechnol. Biochem. 66, 2483–2486 (2002).
Fujiwara, Y., Kono, M., Ito, A. & Ito, M. Anthocyanins in perilla plants and dried leaves. Phytochemistry 147, 158–166 (2018).
Iwashina, T., Kitajima, J. & Matsumoto, S. Luteolinidin 5-O-glucoside from Azolla as a stable taxonomic marker. Bull. Natl. Mus. Nat. Sci. 36, 61–64 (2010).
Varone, L. & Gratani, L. Physiological response of eight Mediterranean maquis species to low air temperatures during winter. Photosynthetica 45, 385–391 (2007).
Elsheery, N. I. & Cao, K.-F. Gas exchange, chlorophyll fluorescence, and osmotic adjustment in two mango cultivars under drought stress. Acta Physiol. Plant. 30, 769–777 (2008).
Tattini, M. et al. Differential accumulation of flavonoids and hydroxycinnamates in leaves of Ligustrum vulgare under excess light and drought stress. New Phytol. 163, 547–561 (2004).
Agati, G., Stefano, G., Biricolti, S. & Tattini, M. Mesophyll distribution of ‘antioxidant’ flavonoid glycosides in Ligustrum vulgare leaves under contrasting sunlight irradiance. Ann. Bot. 104, 853–861 (2009).
Majer, P., Neugart, S., Krumbein, A., Schreiner, M. & Hideg, É. Singlet oxygen scavenging by leaf flavonoids contributes to sunlight acclimation in Tilia platyphyllos. Environ. Exp. Bot. 100, 1–9 (2014).
Brossa, R., Casals, I., Pintó-Marijuan, M. & Fleck, I. Leaf flavonoid content in Quercus ilex L. resprouts and its seasonal variation. Trees 23, 401–408 (2009).
Toda, K., Takahashi, R., Iwashina, T. & Hajika, M. Difference in chilling-induced flavonoid profiles, antioxidant activity and chilling tolerance between soybean near-isogenic lines for the pubescence color gene. J. Plant Res. 124, 173–182 (2011).
Wang, G., Cao, F., Chang, L., Guo, X. & Wang, J. Temperature has more effects than soil moisture on biosynthesis of flavonoids in Ginkgo (Ginkgo biloba L.) leaves. New Forest 45, 797–812 (2014).
Zhu, H. et al. Anthocyanins function as a light attenuator to compensate for insufficient photoprotection mediated by nonphotochemical quenching in young leaves of Acmena acuminatissima in winter. Photosynthetica 56, 445–454 (2018).
Johnson, C. A. et al. Effects of temperature and resource variation on insect population dynamics: the bordered plant bug as a case study. Funct. Ecol. 30, 1122–1131 (2016).
Yan, J.-H., Zhou, G.-Y., Zhang, D.-Q. & Wang, X. Spatial and temporal variations of some hydrological factors in a climax forest ecosystem in the Dinghushan region. Acta Eeol. Sin. 23, 2359–2366 (2003).
Wellburn, A. R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 144, 307–313 (1994).
Heimler, D., Vignolini, P., Dini, M. G. & Romani, A. Rapid tests to assess the antioxidant activity of Phaseolus vulgaris L. dry beans. J. Agr. Food Chem. 53, 3053–3056 (2005).
Nguyen, P. M., Kwee, E. M. & Niemeyer, E. D. Potassium rate alters the antioxidant capacity and phenolic concentration of basil (Ocimum basilicum L.) leaves. Food Chem. 123, 1235–1241 (2010).
Acknowledgements
This work was funded by the National Natural Science Foundation of China (31570398, 31870374). The study was also supported by Guangdong Province Natural Science Foundation (2017A030313167) and Open Foundation of Guangdong Provincial Key Laboratory of Biotechnology for Plant Development, South China Normal University (2018KF0203). We are grateful to managers of the Dinghushan biosphere reserve for their permission to carry out the research and for their assistance during the fieldwork.
Author information
Authors and Affiliations
Contributions
T.-J.Z. and C.-L.P. designed research; T.-J.Z., X.-T.L. and X.-D.H. carried-out the experiments. X.-S.T. and C.-L.P. analyzed the data and discussed the results. T.-J.Z. wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhang, TJ., Tian, XS., Liu, XT. et al. Seasonal variations in group leaf characteristics in species with red young leaves. Sci Rep 9, 16529 (2019). https://doi.org/10.1038/s41598-019-52753-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-52753-x
This article is cited by
-
Why do red/purple young leaves suffer less insect herbivory: tests of the warning signal hypothesis and the undermining of insect camouflage hypothesis
Arthropod-Plant Interactions (2022)
-
The relationship between anthocyanin accumulation and photoprotection in young leaves of two dominant tree species in subtropical forests in different seasons
Photosynthesis Research (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
