
Abstract
Gametophytic apomixis is a way of asexual plant reproduction by seeds. It should be advantageous under stressful high altitude or latitude environment where short growing seasons, low temperatures, low pollinator activity or unstable weather may hamper sexual reproduction. However, this hypothesis remains largely untested. Here, we assess the reproductive mode in 257 species belonging to 45 families from the world’s broadest alpine belt (2800–6150 m) in NW Himalayas using flow cytometric seed screen. We found only 12 apomictic species, including several members of Poaceae (Festuca, Poa and Stipa), Rosaceae (Potentilla) and Ranunculaceae (Halerpestes, Ranunculus), which are families typical for high apomict frequency. However, several apomictic species were newly discovered, including the first known apomictic species from the family Biebersteiniaceae (Biebersteinia odora), and first apomicts from the genera Stipa (Stipa splendens) and Halerpestes (Halerpestes lancifolia). Apomicts showed no preference for higher elevations, even in these extreme Himalayan alpine habitats. Additional trait-based analyses revealed that apomicts differed from sexuals in comprising more rhizomatous graminoids and forbs, higher soil moisture demands, sharing the syndrome of dominant species with broad geographical and elevation ranges typical for the late-successional habitats. Apomicts differ from non-apomicts in greater ability of clonal propagation and preference for wetter, more productive habitats.
Similar content being viewed by others

Introduction
There is a general opinion that gametophytic apomixis, the mode of plant reproduction via seeds formed by asexual way from gametophytic tissue, is more frequent in cold environments of higher latitudes and altitudes1,2,3,4. Apomicts might be better colonizers than sexual species in arctic and alpine areas because of uniparental reproduction and ability to set up populations by single individuals5,6, thereby compensating for reduced fecundity resulting from pollen limitation in small and isolated populations7. However, the hypothesis of higher frequency of apomictic plants in cold environments has been rarely tested8, but when tested, the prominent role of apomixis is questioned8,9,10. Several studies on Ranunculus kuepferi found clear connection between higher elevation and apomixis, but simultaneously between apomixis and polyploidy. It seems that these genomic and environmental features are bound together – polyploids appear rather in colder climate of high elevation, and polyploids have tendency to reproduce by apomixis11,12,13. As understanding of this polyploid complex increased, also opposite relationship was proposed – polyploidy might be bound to apomixis14,15. Here we analyse to our knowledge the largest dataset of alpine plant species to assess whether the occurrence of apomictic plants increase along Himalayan elevational gradient, starting around 3000 m and ending at the absolute limit of vascular plants at 6000 m a.s.l.
There are several mutually non-exclusive explanations why the frequency of apomicts should increase upwards or polewards. (1) Cold areas may lack pollinators necessary for certain plants to reproduce sexually16,17; this, however, does not explain pseudogamy where pollination is still required for endosperm development18. (2) As there is no recombination during meiosis, apomixis keeps all successful gene combinations and hence suitable adaptations, which is advantageous in special but stable habitat (occurrence of so called fixed heterozygosity in apomicts is reviewed by Gornall19). (3) Pleistocene glaciations have increased the proportion of apomictic species by promoting hybridisation and afterwards polyploidization between previously isolated populations that came in contact within glacial refugia20,21; isolation of such hybrids in the newly deglaciated arcto-alpine areas followed22,23,24,25. (4) Species in cold areas have mainly disjunctive populations, therefore, according to the Baker´s law26,27 (the law formulated by Stebbins28), the autonomic or self-compatible species might be better able of colonising these areas.
Other predictions, supporting the affinity of apomicts to cold areas, have been more recently added to the original above-mentioned arguments defined by Stebbins5. For instance, apomixis may save energy otherwise used for expensive process of meiosis24., or, the second way of sparing energy is creating no pollen, which, however, is not universal, as pseudogamous plants need pollen to fertilise their endosperm29. Nevertheless, some of the abovementioned arguments are just theories, which can be easily challenged. Firstly, the Baker´s law is a generalized view on behaviour of an apomictic population based only on few examples of apomictic animal populations. Secondly, the ecological argument presupposing absence of pollinators was recently refuted as there are still effective pollinators occurring in cold arctic and alpine habitats30,31. Moreover, pollination does not depend on pollinators solely, as partial or full autogamy is widespread among angiosperms including apomicts32,33,34. Despite that, all these theoretical predispositions about higher occurrence of apomixis in extreme conditions have been considered as a fact for a long time1 without proper testing.
The problematic of higher frequency of apomictic plants in cold environments was not thoroughly studied until recently (but see the extensive research on R. kuepferi11,12,13,14,15). Gregor35 made a statistical analysis of frequency of apomicts in phytosociological relevés from Central Europe. The apomixis was highly positively correlated with altitude in his compilation. On the contrary, Hörandl et al.8 tried to verify the theory of higher frequency of apomicts in the European Alps by analysing 14 subnival species, all dependent on insect pollinators, to find out whether there is some effect of pollinator limitation8. From these species, only Potentilla crantzii (Crantz) Beck ex Fritsch, an apomictic plant described earlier by Smith36 was apomictic while other species showed seeds originated by sexual way, indicating that apomixis might be rather rare in high elevations. However, there are extreme high mountain areas where the relation between apomixis and elevation might be stronger. Himalayas provide extensive alpine environment potentially less favourable for insect pollinators compared to European mountains and therefore more favourable for formation of apomixis due to severe ecological constraints37.
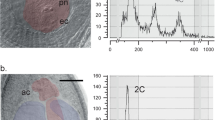
There are several ways how to study the apomixis, most effective being a flow cytometric seed screening (FCSS38), which became popular in the last few decades8,15,39. FCSS measures DNA content in mature angiosperm seeds and infer the reproductive mode based on DNA content of the embryo and the endosperm without a need to know the actual genome sizes or ploidy levels40,41. Sexual angiosperm seed derived from reduced gametes and normal double fertilization has 2C embryo (formed from a 1C egg cell and a 1C sperm cell) and 3C endosperm (a 2C central nucleus of the embryonic sac plus a 1C sperm cell)7,42. Apomictic seeds have different ratios the embryo and the endosperm genome sizes43. The ratios 2/4, 2/5 or 2/6 are the most frequent under automonous endosperm development, pseudogamy with a reduced male gamete and an unreduced male gamete, respectively41. However, there are many other possibilities depending on the actual way of embryo and endosperm formation and ploidy levels of taxa involved (in cases when pollen- and egg- parents have different ploidy levels) but most of these combinations are reliably distinguished from sexual seeds39,44,45.
It has been widely assumed that apomixis is frequently associated with vegetative, clonal propagation. Vegetative reproduction, in which parental genets produce new ramets capable of independent growth and dispersal46, is commonly associated with the perennial life form, longevity, and occurrence in cold habitats in which sexual recruitment is often restricted. According to Barrett47, extensive vegetative growth by clonal rhizomes, stolones and root sprouts can disrupt the functioning of sexual polymorphisms; mutations reducing fertility may lead to sexual dysfunction and even the loss of sex in populations in which clonal propagation predominates. However, there have been few efforts documenting whether apomicts differ from sexuals in the adaptive strategies including vegetative propagation and other ecophysiological traits enabling plants to cope with harsh alpine conditions.
In this study, we focused on testing the hypothesis about increasing frequency of apomicts with increasing elevation in the northwestern Himalayas. By collecting 257 species along unprecedented elevation gradient including cold deserts, steppes, alpine meadows and subnival zone with world’s highest growing plants37 (Fig. 1), we aimed at (1) screening representative number of taxa across major habitats and the angiosperm groups for determining their reproductive mode using flow cytometry, (2) determining the specific way of reproduction in apomictic species, (3) linking the occurrence of apomictic species with habitat conditions, especially species’ elevational optima, (4) assessing whether apomicts differ from sexuals in their abilities to cope with harsh alpine conditions by comparing their plant ecophysiological traits, clonal growth strategies and species ecological preferences.

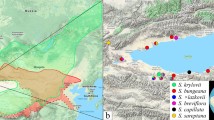
Map of localities in Ladakh. Background map is based on. OpenStreetMap data available under the Open Database License: http://opendatacommons.org/licenses/odbl/1.0/.
Results and Discussion
Out of 257 measured taxa, reproductive mode was inferred for 220 while 37 species had no detectable endosperm signal. 211 species showed sexual way of seeds formation (sometimes sexuality was found together with apomixis in a species). We found only 13 apomictic taxa (including two subspecies of Poa pratensis) belonging to seven genera and four families. Most of the apomictic species studied in the Himalayas were not tested for apomixis yet, so we cannot compare reproduction systems within species with other locations. The most of the few cases, which we were able to compare across species ranges, indicate that apomictic species have the same reproduction system at lower elevations as well as at higher elevations
3 out of 12 apomictic species belong to the genus Potentilla (Rosaceae), namely Potentilla pamirica Th.Wolf, Potentilla sericea L., Potentilla sojakii Dikshit & Panigrahi and 4 belong to the genus Poa (Poaceae): Poa alpina L., Poa attenuata Trin., Poa pratensis L., Poa sterilis M.Bieb. Other apomictic species were Biebersteinia odora Fisch. (Biebersteiniaceae), Festuca olgae (Regel) Krivot., Stipa splendens Trin (=(Achnatherum splendens (Trin.) Nevski) (both Poaceae), Halerpestes lancifolia (Bertol.) Hand.-Mazz., and Ranunculus membranaceus Royle (both Ranunculaceae). Poa, Potentilla, and Ranunculus are genera well-known for high frequency of apomictic species. Although Stipa splendens have no apomictic relatives in the genus and apomicts in the genus Festuca are also rather rare, both genera belong to family Poaceae, which is among the three families with the highest frequency of apomixis (Asteraceae, Rosaceae, Poaceae). Although there is no information on apomixis in Halerpestes, this genus is closely related to Ranunculus48 in which apomixis in the family is known. There is no information on apomixis within Bierbersteiniaceae yet; our data on B. odora are the first for this family in this respect.
Differences in plant traits and elevation optima between apomictic and sexually reproducing species
Apomicts and sexuals did not differ in elevation optima, minima and maxima, Raunkiær’s plant life-forms, leaf and root nutrient concentrations and root non-structural carbohydrates (Table 1). They however significantly differed in elevation ranges (pseudo F = 5.1, P = 0.035), clonal growth forms (pseudo F = 3.3, P = 0.004), ecological indicator values (pseudo F = 2.5, P = 0.031), and geographical occurrence (pseudo F = 12.7, P = 0.004). Apomictic species were clonal rhizomatous graminoids and forbs, with higher frequency of occurrence and soil moisture demands, sharing the syndrome of dominant, competitive species with broad geographical and elevation ranges (Fig. 2) typical for the stable, late-successional habitats. Apomicts did not show preference for higher elevations, even in these extreme Himalayan high-mountain habitats; they were instead bound to taxonomically related groups known for apomixis.

Apomictic species and their elevation optima and ranges in Ladakh, NW Himalayas. Note that it spans the whole range of elevations in the area and the apomictic species are not concentrated in higher elevations.
Our findings corroborate previous studies46 indicating that apomixis is associated with the on-spot persistence, high plant longevities, and occurrence in stable habitats where extensive clonal growth may disrupt the functioning of sexual polymorphisms, resulting in mutations reducing fertility, which may lead to sexual dysfunction and even the loss of sex. Apomicts in NW Himalayas form ecologically distinct group differing from non-apomicts in greater ability of clonal propagation and preference for wetter, more productive habitats. This implies that apomicts are often effective in reproducing and spreading, but successful are only when growing in rather stable habitats, being less successful in colonizing rapidly changing environment such as new substrate after deglaciation49,50.
Apomictic species of genus Potentilla
Three apomicts recruited from the genus Potentilla, including P. pamirica, P. sericea, and P. sojakii. They all showed pseudogamous development of seeds. The ratio of embryo/endosperm was mostly 2/6, which means fertilisation of central cell by an unreduced sperm cell or by two reduced sperm cells (Fig. 3e). The second possible way is more probable because we know from former studies that pollen of Potentilla is mostly reduced51,52. Moreover, this type of endosperm fertilisation was also observed in Rosaceae45,53. Alternatively, other species of higher different ploidy level might contribute unreduced-like pollen at site where several species co-occur54.

FCSS histograms of selected apomictic species and a sexual species Stipa subsessiliflora. Peeks show the amount of nuclei with relative DNA amount represented by intensity of DAPI fluorescence.
Apomixis in the genus has been well studied. Dobeš et al.55 provides a summary of reproduction systems for 22 series of the genus Potentilla, of which apomixis was confirmed in 14 series, but the sexuality co-occurred in 10 out of these 14 series. The apomictic series occur exclusively in phylogenetically young core Potentilla. However, study of apomixis in this genus has its limits. The first is caused by hardly detectable endosperm tissue56,57, which is sometimes completely missing58. In our case, 44 out of 57 samples were without detectable endosperm tissue. The second obstacle arises with occasional deviation from standard eight nuclei in embryo-sac. Dobeš et al.55 observed only one polar nucleus in endosperm of Potentilla indica, with flow cytometer histograms indicating regular sexuality; however, an AFLP analysis of progeny revealed an apomictic origin of seeds.
In spite of missing data on reproduction mode in some Himalayan Potentilla spp., for some species within our dataset, reports from other areas are available. Predisposition for apomixis was already known in one species (P. sericea; polyploidization by hybridization)59. Potentilla venusta is known to be apomictic55; our analyses were, however, without readible signal and we therefore cannot determine the reproduction system. Potentilla multifida showed sexual reproduction in our sample, even at 5000 m a. s. l., while Dobeš et al.55 described 18 apomictic embryo-sacs, seven parthenogenetic embryos, and no seeds of sexual origin. The same case is Potentilla atrosanguinea with 15 out of 22 studied seeds being clearly apomictic in Dobeš et al.55, while all our samples were all sexual. Christoff & Papasova60 also described P. atrosanguinea as a sexual, but his material came from botanical gardens, and so it is not directly comparable with our wild populations. Concerning reproduction modes of other sampled species which were unfortunately without clear signal, Rani et al.61 observed meiotic abnormality in Potentilla gelida in Western Himalayas, while another study by Jeelani et al.59 showed polyploidy in Potentilla sericea but no meiotic abnormalities. There are no previous studies concerning breeding systems in P. bifurca, P. evestita, P. pamirica, P. sojakii or P. turczaninowiana. However, P. bifurca was involved in a phylogenetic study by Eriksson et al.62. It placed the species outside the core Potentilla, which suggests that P. bifurca should not be able to reproduce by apomixis, which was confirmed by our findings.
Apomictic species in Ranunculaceae
The species Ranunculus membranaceus and closely related species Halerpestes lancifolia produced seeds in the analogous way as Potentilla mentioned above: the ratio was always 2/6, which indicates pseudogamy with one or two sperm cells involved in formation of endosperm (Fig. 3c,f). This type of endosperm fertilisation was described in Ranunculus auricomus7,45,63, the intensively studied facultative apomictic complex64. We can expect that apomixis in R. membranaceus is of the same type, as this species is sister to the clade containing the R. auricomus complex65. As apomictic Ranunculus spp. are extensively investigated, mechanism of polyploid apomictic seeds has been suggested13. It seems that polyploidization happens regularly mainly via triploid bridge with an unreduced egg cell. This loss of meiosis in female gametes might be the first step for development of apomixis in Ranunculus polyploids. However, Klatt et al.15 supported rather stress-dependent loss of sex resulting in polyploidization rather than polyploidization as a preadaptation for apomixis. The second apomictic species of Ranunculaceae was Halerpestes lancifolia with the same embryo/endosperm ratio as R. membranaceus. It is an unexpected observation, because no other apomictic genera except Ranunculus were discovered within the family Ranunculaceae so far. Nevertheless, the genus Halerpestes is closely related to Ranunculus s.str.66 and occurrence of the apomixis of Ranunculus-type might be expected.
Apomictic species in Poaceae
The family Poaceae is also known for high frequency of apomictic species. In our data set, 16 species of Poaceae were tested, from which one is an apomictic species of the genus Festuca (namely Festuca olgae), four belong to the genus Poa (Poa alpina, Poa attenuata, Poa pratensis, Poa sterilis) and one species belong to the genus Stipa (Stipa splendens).
Festuca olgae is a facultative type of an apomict. We measured 22 seeds from 3 plants and just two seeds were apomictic with endosperm/embryo ratio 6/2 (Fig. 3b). Such facultative apomixis is known to occur in Poaceae67 and it happens mostly in an aposporous plant which creates an embryo-sac directly from nucellus68. A facultative apomict might have an advantage over strict sexual or apomictic. In fact, it has advantages from both of them. The plant can produce genetically variable offspring to improve its features in changing environment, on the other hand it produces also stable offspring which is well adapt to current conditions. Even though apomixis in the genus is not very common, it was observed already69 and; in addition, the genus Festuca is quite close related to the genus Poa70, which is one of the best known apomictic genera67.
The genus Poa had mostly pseudogamous mode of reproduction in our analyses (in majority cases with the ratio 5/2, in the only one case with the ratio 6/2; Fig. 3d). This type of apomixis appeared in all four tested species: P. alpina, P. attenuata, P. pratensis and P. sterilis. One seed of Poa attenuata from our measurement shows autonomy. Although the presence of pseudogamy and autonomy in one species is very rare, it has been described in Poa nervosa67. Nevertheless, the second and more probable explanation can be found: Nogler63 described a situation where one of two central nuclei was degraded, therefore one unreduced or two reduced sperm cells may cause the “autonomic” ratio.
Similarly to Potentilla spp., the genus Poa was studied well for apomixis and for many species data about reproduction mode are available. Poa attenuata (collected on the north side of Ťan-Šan, in 1200 m a. s. l.) was tested by Kelley et al.67 with exclusively pseudogamous apomixis (with the embryo/endosperm ratio 2/5). Nevertheless, the genus is characteristic for facultative apomixis, which was demonstrated by two sexual seeds in two Himalayan Poa species (P. pratensis subsp. staintonii, P. sterilis). This phenomenon was described for both species67.
The case of Stipa splendens is also unique. There is no reported apomixis in Stipa71 or related taxa, however, our samples showed pseudogamous and autonomous endosperm formation, similarly to e.g. what was observed by Kelley et al.67 in Poa nervosa. Within the pseudogamous seeds, majority of S. splendens seeds had ratio 2/5 (Fig. 3g), but also the ratio 2/6 was not unique. One seed had a ratio 3.32 which should be formally closer to the ratio 2/7, nevertheless, this minor deviation is probably caused by pollen with bigger genome size. That at least two genome sizes of embryos occur in a population showed our measurements with standard (Supplementary Table S1), corresponding probably to diploids and triploids, so the ratio close to 2/7 on a diploid mother may correspond to pollination with triploid pollen. Our data revealed also two samples of Stipa splendens with unusual peaks ratios 2.11 and 2.17. In one case the sample might be a triploid and the ratio was than consistent with pollination with diploid’s reduced (haploid) pollen (3/7), in the other case we do not have a clear explanation for the observed pattern (the role of rare embryonic sac modifications and/or unbalanced pollen may be suspected).
Apomixis in Biebersteinia
Biebersteinia odora was one of the unexpected apomictic species revealed in Ladakh as there is no record of apomixis in the family so far. Moreover, more ploidy levels of embryos and several ways of endosperm origin were revealed. The results showed stable genome size ratio of mother plant to standard (2.7–3), however, the ratio of embryo to standard varied (Supplementary Table S2).
In two populations of three sampled populations, genome size of maternal plants (measured from leaves) was uniform, and these plants produced mainly embryos of the same genome size; we consider them diploid (normal). Majority of seeds showed the embryo/endosperm ratio 2/5 (Fig. 3a), which indicates pseudogamous apomictic embryonic sac pollinated by one sperm of reduced pollen. Indeed, it is known that in apomictic groups mainly female meiosis is hampered while male meiosis often remains unchanged71. About 10% of seeds showed the ratios 2/6 (diploid pollen or two sperms of haploid pollen) or 2/7 (tri-nucleate central cell and haploid pollen). Single seeds had the profiles 2/8 (tri-nucleate central cell and one diploid or two haploid sperms), 2/10 (probably tri-nucleate central cell and two diploid sperms), we also found a few seeds with ratios indicating presence of aneuploid (between 1x and 2x pollen). One seed had 4x embryo and 6x endosperm and most likely is so called BIII hybrid (firstly described by firstly Rutishauer72, also “U-hybrids” by Asker73) resulting from fertilization of apomictic (unreduced) embryonic sac with diploid pollen. Such embryos are usually formed as 2n + n hybrids and, more rarely, as 2n + 2n hybrids74, and are important mechanism allowing ploidy level change. One seed had probably 3/8 profile; we do not have any simple explanation of its origin (types or reproduction systems are showed in the Supplementary Table S2). The last population showed remarkable variation in embryo genome size/ploidy levels. Unfortunately, it was the first population of B. odorata we collected and as we did not expect any variation, leaf samples were not available. Most of the embryos had calculated ploidy levels between 2x and 3x, some 2x and some 4x. About half of seeds had similar embryo/endosperm ratio as above (2/5 to 2/8) while the rest are probably aneuploid. Altogether, the recorded variation in all populations show that there are probably more ploidy levels in B. odora populations and probably a mixture of sexual and apomictic processes, involving also unbalanced gametes of odd ploidy levels (probably triploids). However, to understand this variation, sampling of more populations, more individuals from each population and both maternal plant and seed ploidy levels is needed. Biesternia odorata is thus an interesting organism for further study.
Absence of apomixis in certain families
Finally, very important is to mention the families which we expected to be apomictic but no apomictic representative was found. Firstly, Asteraceae contains considerable amount of all known apomictic species but we did not find any apomicts among 36 different species from several genera we tested. Among the tested genera there were for example Crepis75, Leontopodium76,77, Erigeron78,79,80,81,82 which all possess apomixis but our samples were clearly sexual. We analysed many species of Artemisia83,84 which is known for apomictic tendencies but with the same negative result. On the other hand we did not have an opportunity to sample Taraxacum; it is likely that in this strongly apomictic genus85 some apomictic lineages might be present even in the Ladakh flora. Among Boraginaceae with known apomixis, the genus Cynoglossum86 in our dataset was sexual. Similarly it holds true for the other families with sporadic presence of apomixis (e.g. Brassicaceae87,88 or Lamiaceae89).
Conclusion
Data obtained from seed samples collected in Ladakh revealed 12 species with gametophytic apomixis out of total 257 species collected. Among 12 apomictic species, four species were already described as apomictic (Poa alpina, Poa attenuata, Poa pratensis, Poa sterilis). Most of the apomictic species were not investigated through apomixis yet, therefore we cannot say whether there is some diversity between reproduction systems in different geographical conditions within the species. Nevertheless, in genera Potentilla, Poa and Ranunculus are well-known apomicts and apomixis within these genera was expected. Newly discovered apomixis in Halerpestes lancifolia, Stipa splendens and Biebersteinia odora was surprising and this finding is a valuable contribution to knowledge about apomictic groups. Our results clearly show that Ladakh apomicts mostly occur in taxonomical groups or lineages with already known apomictic species, which leads to negation of the theory of higher incidence of apomicts in high mountains and can reflect only tendency to apomixis within individual taxonomical lineages. Furthermore, in Ladakh occur two sexual species of the genus Potentilla, which were already described as possible apomictic. The trait-based comparison revealed that apomicts differed from sexuals in comprising more rhizomatous graminoids and forbs, higher soil moisture demands, sharing the syndrome of dominant species with broad geographical and elevation ranges typical for the late-successional habitats. Apomicts in NW Himalayas have greater ability of clonal propagation and preference for wetter, more productive and stable habitats where they effectively reproduce and spread; they seems to be less successful as pioneers in climate-induced uphill migration or new substrate colonization after deglaciation.
To sum up, predisposition for apomixis seems to be bound to specific phylogenetic lineages. The genetic principles of development of apomixis are still not fully described, but it is known that it includes many genetic and epigenetic mechanisms90. As the mechanism is so complex, apomixis is not a dominant reproductive strategy (less than 1% of Angiosperms91) and its establishment is difficult and is thus known only in a few lineages which succeeded in evolving such unusual method of asexual dispersal.
Material and Methods
Study area
The study was conducted in the Himalayan Mts. in Ladakh, Jammu and Kashmir State, India. Ladakh encompasses ca. 80.000 km2 of the Trans-Himalaya, delimited by the Eastern Karakoram Range in the north, the Great Himalaya Range in the southwest and Tibetan Plateau in the east. The studied localities ranged from 3000 m in Indus-Shayok-Nubra Valleys to 6150 m in Tso Moriri Lake area (Fig. 1). Desert and semi-desert occupy the (relatively) lowest elevations of the Indus Valley and its major tributaries (ca. 2900–3800 m a.s.l.). The subalpine vegetation belt stretches from approximately 3800 up to 5000 m a.s.l. in east Ladakh, and is widely dominated by cold steppe vegetation37. The alpine belt extends between ca. 4500 and 5200 m (occasionally to 5500 m) a.s.l. and hosts alpine grasslands, including the characteristic moist alpine turf of Kobresia pygmaea (C.B.Clarke) C.B.Clarke. A very sparse subnival vegetation zone is developed up to 6150 m a.s.l.
The elevational gradient in our study region is tightly related to a temperature and aridity gradient49,50. It is therefore warmer at low-elevation deserts, with intense drought due to lower precipitation and colder and more humid at high-elevation alpine and subnival zones. The length of the growing season decreases linearly with increasing elevation, from 270 to 30 frost-free days, while mean growing season temperature decreases from 14 to 3 °C between 3000 and 6150 m. Daily temperatures in the study area vary from 0 to 30 °C in summer and from –40 to –10 °C in winter. Annual precipitation is about 50 mm in the desert and semi-desert belt (falling mostly during the Indian summer monsoon), 100 mm in the alpine steppes, and 150–250 mm in the alpine and subnival zones92. Precipitation above 5000 m a.s.l. is mostly falling as snow and is more frequent in summer than in winter37. Soils have a coarse-grained structure, with a high percentage of large gravel, low water and organic matter content, high pH (7–8) and relatively high concentrations of total N and P93.
Seed collections
Seeds were collected during August–September 2013–2015 at 137 localities. To reveal possible variability in reproduction systems, up to five individuals per species and population were collected; in selected species 10–15 populations was sampled. Seeds from each individual were stored in a separate bag. We used also some seeds from 25 species collected in 2009 (for a different study), however, in that case only population samples containing a mixture of seeds from several individuals were available. The seeds were stored in paper bags, allowing air to flow into the bag drying the seeds. Vouchers are deposited in herbarium of Institute of Botany, Academy of Science of the Czech Republic, Třeboň. Seeds of 36 species were collected directly from the herbarium vouchers that were collected between years 1999–2006. Locality details for all samples are specified in Supplementary Table S3.
Flow cytometry
The relative fluorescence intensities of embryo and endosperm nuclei were analysed using the Partec PA II flow cytometer (Partec GmbH., Münster, Germany, now Sysmex) equipped with a mercury arch lamp. DAPI was used as a DNA stain. At least 5,000 particles were recorded. Two different protocols of sample preparation were tested and the one that gave sufficient results for a particular species (low coefficients of variance of the peaks and low background noise) was selected. (1) One-step protocol employing a seed buffer (Matzk et al.38 with minor modifications of Krahulcová & Suda94). Seeds were chopped with a razor blade in 1 ml of the seed buffer [0.1 M Tris, 5 mM MgCl2.6H2O, 85 mM NaCl, 0.1% (v/v) Triton X-100]. The suspension with filtrated through a 42 μm mesh and DAPI was added immediately after filtration (final concentration 4 μl/ml). The samples were analysed after a few minutes of staining. (2) Simplified two step protocol (Doležel et al.95, with minor modifications). Seeds were chopped in 400 μl of ice-cold Otto I buffer [0.1 M citric acid monohydrate, 0.5% (v/v) Tween 20]. The suspension with filtrated through a 42 μm mesh. After several minutes of incubation at room temperature, 800 μl of the staining solution was added, consisting of Otto II buffer [0.4 M Na2HPO4.12H2O], β-mercaptoethanol (0.2%) and DAPI (final concentration 4 µl/ml). Samples were run on the flow cytometer after several minutes of staining. In case of an insufficient results, different incubation time (both longer and shorter) were tested.
One to three seeds originating from one individual were pooled in a sample (however, in case of insufficient quality one-seeded analysis was always performed). If available, at least 3 seeds per individual and at least 3 individuals per population were analysed. In case of very small seeds, five or even ten seeds per sample were pooled to get strong enough signal. When genome or ploidy variation or variation in reproductive mode was suspected from pooled samples, additional analyses of single seeds were performed. In cases of genome size variation (in Stipa splendens Trin. and Biebersteinia odora Stephan ex Fisch.), samples were measured with an internal standard, which was a leaf tissue of Pisum sativum ‘Ctirad’ (2C = 9.0996) or of Bellis perennis (2C = 3.38 pg). For Biebersteinia odora, during the second fieldwork in 2014 we collected also leaf samples of the individuals from which seeds were collected. The leaf samples were dried in silica-gel and analysed using the two-step protocol (see above). Number of measured seeds and the protocol used are specified in Supplementary Table S4 for each species.
The fluorescence histograms were analysed in FlowJo 10 software (FlowJo LLC, Ashland, Oregon). The fluorescence intensity, coefficient of variation and number of nuclei were recorded for the G0/G1 peaks of the embryo and the endosperm. In case when the first peak of the endosperm was low due to strong endopolylpoidy (e.g. in Astragalus spp., Oxytropis spp., Cicer arietinum L., Colutea nepalnesis Sims, Atriplex schugnanica Iljin, Corispermum tibeticum Iljin, Christolea crassifolia Cambess, Galium pauciflorum Willd. ex DC., Leptorhabdos parviflora (Benth.) Benth., Pedicularis spp.), the parameters of the second peak of the endosperm were recorded and the mean fluorescence intensity divided by the two. The ratio of the mean fluorescence intensities of the embryo and the endosperm was calculated and a reproductive system was inferred from it following Matzk41.
For variable Biebersteinia odora seeds measured with the internal standard, we calculated the relative genome sizes of the embryo and the endosperm to estimate their origin (genome sizes of the gametes and various modes of fertilization, following e.g. Dobeš et al.39). We considered sexual reproduction, apomixis with autonomous endosperm and apomixis with pseudogamy, i.e. fertilization of endosperm only with one or two reduced or unreduced sperms. Among the different scenarios, the one under which all calculated gamete genome sizes are close to the embryo genome size (unreduced gametes) or half of it (reduced) was considered the most likely. In a few cases calculated relative genome sizes were multiples of the embryo genome size, indicating that pollen probably originated from an individual of higher ploidy level.
Relative size of embryo and endosperm in seeds
Furthermore, we prepared a summary of a relative amount of the endosperm in seeds, based on counts of nuclei of the embryo and the endosperm peaks (Supplementary Table S5). Percentage of endosperm in a seed was expressed as the ratio of count of nuclei of the endosperm peak to the sum of both peaks. An average of percentage of endosperm in a seed was counted for each family. This summary might serve as a tool for following studies dealing with FCSS, including identifying of families without detectable endosperm.
Plant morphological and ecophysiological trait measurement
To assess which traits are associated with apomicts and those reproducing sexually, several plant traits, clonal growth strategies and species ecological information for all 257 species analysed for the mode of reproduction were recorded based on Cornelissen et al.97 and Doležal et al.37,98. More than 10 individuals were collected for each species at different elevations covering most of the species altitudinal ranges in Ladakh during several expeditions in years 2008–201637. We measured several plant traits relevant to competitive ability (plant height), allocation (total, leaf, stem and root biomass), growth (SLA - specific leaf area, LDMC and StDMC - leaf and stem dry matter content, leaf and root NP concentrations), carbon storage (starch, fructans as non-structural carbohydrates), drought and frost tolerance (free sugars and sugar alcohols), water use efficiency (δ13C), generative (seed mass) and vegetative (clonality and lateral spread) propagation. These traits are important indicators of plant resource-use strategy, reflecting a fundamental trade-off between the efficient conservation of nutrients (low SLA, less negative δ13C, low foliar N and high LDMC) and the rapid production of biomass (e.g. high SLA, high foliar NP, low LDMC, long rhizomes). SHIMADZU UV - 1650PC spectrophotometer was used to determine phosphorus after digestion in HClO4. The δ13C, as well as total carbon and nitrogen, were measured using an elemental analyzer coupled to an IRMS at the Stable Isotope Facility, UC Davis, USA. Megazyme total starch assay procedure (www.megazyme.com) was used to determine starch and fructans; ethanol-soluble sugars were determined through anion exchange chromatography with pulsed amperometric detection. Soluble sugars included sugar alcohols such as glycerol, xylitol and arabitol, and simple sugars such as glucose, fructose, sucrose and galactose. The total non-structural carbohydrates (NSC) were calculated as the sum of all analysed carbohydrates.
Vegetative clonal propagation
In addition to quantitative traits, each species was classified into one of 20 clonal growth forms based on which organ (rhizomes versus primary tap-roots) provides connections between offspring shoots, if this organ is short or long, if a plant is able to form adventitious roots, and if there are special storage organs. There are three exceptions: annuals and biennials, woody plants and cushion plants are assessed according to their whole morphology. Furthermore, species were divided into four space occupancy strategies, based on the rate of lateral spread (spreading–more than 10 cm per year; non-spreading–less than 10 cm per year) and persistence of connections between ramets (splitters–plants producing adventitious roots with main root decaying; integrators–plants not producing adventitious roots and/or with perennial main root46).
Species elevational optima and ecological indicator values
To obtain a robust estimate of the elevational optima and ranges of the species, we calculated response curves fitted with HOF models50. Species response curves were derived from 4,150 vegetation plots (each 100 m × 100 m) sampled over the entire Ladakh between 1999 and 2014. The dataset contains more than 122,000 records of occurrence of vascular plant species along exceptional elevational gradient from 2800 m to 6150 m. Species’ optima on five environmental gradients (ecological indicator values) were derived from vegetation composition of 369 plots (each 100 m2) sampled in a stratified design to cover major vegetation types over the study area.
Statistical analyses
To determine which set of traits best explain differences between apomicts and sexuals, we used multivariate RDA analysis using the CANOCO 5 software99. We first analysed all traits together, followed by analyses specific for each group of traits (whole plant traits, organ traits, clonal growth forms and species ecological information, see Table 1). Statistical tests were based on 9999 Monte Carlo permutations.
References
Bierzychudek, P. Patterns in plantparthenogenesis. Experientia. 41, 1255–1264 (1985).
Gregor, T. Apomicts in the vegetation of Central Europe. Tuexenia. 33, 233–257 (2013).
Kearney, M. Hybridization, glaciation and geographical parthenogenesis. Trends Ecol. Evol. 20, 495–502 (2005).
Tilquin, A., & Kokko, H. What does the geography of parthenogenesis teach us about sex? Philos Trans R Soc Lond B Biol Sci. 371 (2016).
Stebbins, G. L. Variation and Evolution in Plants. (New York: Columbia University Press, 1950).
Baker, H. G. Support for Baker’s law – as a rule. Evolution. 21, 853–856 (1967).
Hörandl, E., Cosendai, A. C. & Temsch, E. M. Understanding the geographic distributions of apomictic plants: a case for a pluralistic approach. Plant. Ecol. Divers. 1, 309–320 (2008).
Hörandl, E. et al. Apomixis is not prevalent in subnival to nival plants of the European Alps. Ann. Bot-London. 108, 381–390 (2011).
Hartmann, M. et al. The Red Queen hypothesis and geographical parthenogenesis in the alpine hawkweed Hieracium alpinum (Asteraceae). Biol. J. Linn. Soc. 122, 681–696 (2017).
Paudel, B. R., Shrestha, M., Dyer, A. G., & Li, Q. J. Ginger and the beetle: Evidence of primitive pollination system in a Himalayan endemic alpine ginger (Roscoea alpina, Zingiberaceae). PloS one. 12 (2017).
Kirchheimer, B. et al. A matter of scale: apparent niche differentiation of diploid and tetraploid plants may depend on extent and grain of analysis. J. Biogeogr. 43, 716–726 (2016).
Schinkel, C. C. et al. Correlations of polyploidy and apomixis with elevation and associated environmental gradients in an alpine plant. AoB Plants, 8 (2016).
Schinkel, C. C. et al. Pathways to polyploidy: indications of a female triploid bridge in the alpine species Ranunculus kuepferi (Ranunculaceae). Plant syst. Evol. 303, 1093–1108 (2017).
Kirchheimer, B. et al. Reconstructing geographical parthenogenesis: effects of niche differentiation and reproductive mode on Holocene range expansion of an alpine plant. Ecol. Lett. 21, 392–401 (2018).
Klatt, S., Schinkel, C. C., Kirchheimer, B., Dullinger, S. & Hörandl, E. Effects of cold treatments on fitness and mode of reproduction in the diploid and polyploid alpine plant Ranunculus kuepferi (Ranunculaceae). Ann Bot. 121, 1287–1298 (2018).
Warren, S. D., Harper, K. T. & Booth, G. M. Elevational distribution of insect pollinators. Am. Midl. Nat. 325–330 (1988).
McCall, C. & Primack, R. B. Influence of flower characteristics, weather, time of day, and season on insect visitation rates in three plant communities. Am. J. Bot. 79, 434–442 (1992).
Stenseth, N. C. & Kirkendall, L. R. On the evolution of pseudogamy. Evolution. 39, 294–307 (1985).
Gornall, R. J. Population genetic structure in agamospermous plants in Syst. Assoc. Spec. Vol. (eds Hollingsworth, P. M., Bateman, R. M., Gornall, R. J.) 118–138 (London: Taylor & Francis, 1999).
Carman, J. G. Asynchronous expression of duplicate genes in angiosperms may cause apomixis, bispory, tetraspory, and polyembryony. Biol. J. Linn. Soc. 61, 51–94 (1997).
Lynch, M. Destabilizing hybridisation, general purpose genotypes and geographic parthenogenesis. Q. Rev. Biol. 59, 257–290 (1984).
Hörandl, E. Geographical parthenogenesis: Opportunities for asexuality in Lost Sex (eds Schön, I., Martens, K., Van Dijk, P.) 161–186 (Netherlands, Dordrecht: Springer, 2009).
Thompson, S. L. & Whitton, J. Patterns of recurrent evolution and geographic parthenogenesis within apomictic polyploid easter daises (Townsendia hookeri). Mol. Ecol. 15, 3389–3400 (2006).
Van Dijk, P. J. Ecological and evolutionary opportunities of apomixis: insights from Taraxacum and Chondrilla. Philos. T. Roy. Soc. B. 358, 1113–1121 (2003).
Cosendai, A. C., Rodewald, J. & Hörandl, E. Origin and distribution of autopolyploids via apomixis in the alpine species Ranunculus kuepferi (Ranunculaceae). Taxon 60, 355–364 (2011).
Baker, H. G. Dimorphism and monomorphism in the Plumbaginaceae: 1. A survey of the family. Ann. Bot-London. 12, 207–219 (1948).
Baker, H. G. Self compatibility and establishment after long distance. Evolution. 9, 347–349 (1955).
Stebbins, G. L. Self fertilisation and population variability in the higher plants. Am. Nat. 91, 337–354 (1957).
Hörandl, E. The complex causality of geographical parthenogenesis. New Phytol. 171, 525–538 (2006).
Lefebvre, V., Fontaine, C., Villemant, C. & Daugeron, C. Are empidine dance flies major flower visitors in alpine environments? A case study in the Alps, France. Biol. Letters. 10, 4 (2014).
Williams, P. H., Bystriakova, N., Huang, J. X., Miao, Z. Y. & An, J. Bumblebees, climate and glaciers across the Tibetan plateau (Apidae: Bombus Latreille). Syst. Biodivers. 13, 164–181 (2015).
Rutishauser, A. Die Entwicklungserregung des Endosperms bei pseudogamen Ranunculusarten. Mitt. der Naturf. Ges. Schaffhausen. 25, 1–45 (1954).
Nybom, H. Active self-pollination in blackberries (Rubus subgen. Rubus, Rosaceae). Nord. J. Bot. 5, 521–525 (1986).
Kollmann, J., Steinger, T. & Roy, B. R. Evidence of sexuality in European Rubus (Rosaceae) species based on AFLP and allozyme analysis. Am. J. Bot. 87, 1592–1598 (2000).
Gregor, T. Apomicts in the vegetation of Central. Europe. Tuexenia. 33, 233–257 (2013).
Smith, G. L. Studies in Potentilla L. II. Cytological aspects of apomixis in P. crantzii (Cr.) Beck ex Fritsch. New Phytol. 62, 283–300 (1963).
Doležal, J. et al. Vegetation dynamics at the upper elevational limit of vascular plants in Himalaya. Sci. Rep-UK. 6, 24881 (2016).
Matzk, F., Meister, A. & Schubert, I. An efficient screen for reproductive pathways using mature seeds of monocots and dicots. Plant J. 21, 97–108 (2000).
Dobeš, C., Luckl, A., Hulber, K. & Paule, J. Prospects and limits of the flow cytometric seed screen - insights from Potentilla sensu lato (Potentilleae, Rosaceae). New Phytol. 198, 605–616 (2013).
Krahulcová, A. & Rotreklová, O. Use of flow cytometry in research on apomictic plants. Preslia. 82, 23–39 (2010).
Matzk, F. Reproduction mode screening in Flow Cytometry with Plant Cells: Analysis of Genes, Chromosomes and Genomes (eds Doležel, J., Greilhuber, J., & Suda, J.) 131–152 (John Wiley & Sons, 2007).
Chiarugai, A. Il gametofito femmineo dell Angiosperme nei suoi vari tipi di costruzione e di sviluppo. Nuovo G. Bot. Ital. 34, 5 (1927).
Koltunow, A. M. & Grossniklaus, U. Apomixis: A developmental perspective. Annu. Rev. Plant Biol. 54, 547–574 (2003).
Matzk, F., Meister, A., Brutkovska, R. & Schubert, I. Reconstruction of reproductive diversity in Hypericum perforatum L. opens novel strategies to manage apomixis. Plant J. 26, 275–282 (2001).
Talent, N. & Dickinson, T. A. Endosperm formation in aposporous Crataegus (Rosaceae, Spiraeoideae, tribe Pyreae): parallels to Ranunculaceae and Poaceae. New Phytol. 173, 231–249 (2007).
Klimešová, J., Doležal, J., Dvorský, M., de Bello, F. & Klimeš, L. Clonal growth forms in eastern Ladakh, Western Himalayas: classification and habitat preferences. Folia Geobot. 46, 191–217 (2011).
Barrett, S. C. Influences of clonality on plant sexual reproduction. P. Natl. Acad. Sci. USA 112, 8859–8866 (2015).
Emadzade, K., Lehnebach, C., Lockhart, P. & Hörandl, E. A molecular phylogeny, morphology and classification of genera of Ranunculeae (Ranunculaceae). Taxon 59, 809–828 (2010).
Dvorský, M., Doležal, J., De Bello, F., Klimešová, J. & Klimeš, L. Vegetation types of East Ladakh: species and growth form composition along main environmental gradients. Appl. Veg. Sci. 14, 132–147 (2011).
Dvorský, M., Macek, M., Kopecký, M., Wild, J. & Doležal, J. Niche asymmetry of vascular plants increases with elevation. J. Biogeogr. 44, 1418–1425 (2017).
Asker, S. Chromosome studies in Potentilla. Hereditas. 102, 289–292 (1985).
Asker, S. Polymorphism of Potentilla tamernaemontani and related taxa on Gotland. Hereditas. 102, 39–45 (1985).
Talent, N. & Dickinson, T. A. Chapter 16. Apomixis and hybridization in Rosaceae subtribe Pyrinae Dumort.: a new tool promises new insights. Regnum Veg. 147, 301 (2007).
Asker, S. & Jerling, L. Apomixis in Plants (CRC Press, 1992).
Dobeš, C. et al. Parallel origins of apomixis in two diverged evolutionary lineages in tribe Potentilleae (Rosaceae). Bot. J. Linn. Soc. 177, 214–229 (2015).
Corner, E. J. H. The Seeds of Dicotyledons, Vol 1 (Cambridge University Press, 1976).
Martin, A. C. The comparative internal morphology of seeds. Am. Midl. Nat. 36, 513–660 (1946).
Kalkman, C. Rosaceae in The Families and Genera of Vascular Plants (ed. Kubitzki, K.) 343–386 (Berlin, Heidelberg, Germany & New York, NY, USA: Springer, 2004).
Jeelani, S. M., Kumari, S. & Gupta, R. C. Meiotic studies in some selected angiosperms from the Kashmir Himalayas. J. Syst. Evol. 50, 244–257 (2012).
Christoff, M. & Papasova, G. Die genetischen Grundlagen der apomiktischen Fortpflanzung in der Gattung Potentilla. Mol. Gen. Genet. 81, 1–27 (1943).
Rani, S., Kumar, S., Jeelani, S. M., Gupta, R. C. & Kumari, S. Additions to the cytologically investigated species of Potentilla L. (Rosaceae) from India. Plant Syst. Evol. 298, 485–497 (2012).
Eriksson, T., Donughue, M. J. & Hibbs, M. S. Phylogenetic analysis of Potentilla using DNA sequences of nuclear ribosomal internal transcribed spacers (ITS), and implications for the classification of Rosoideae (Rosaceae). Plant Syst. Evol. 211, 155–179 (1998).
Nogler, G. A. Gametophytic apomixis in Embryology of Angiosperms (ed. Johri, B. M.) 475–518 (Berlin, Germany: Springer, 1984).
Nogler, G. A. Genetics of apospory in apomictic Ranunculus auricomus: 5. Conclusion. Bot. Helv. 94, 411–423 (1984).
Hörandl, E. et al. Phylogenetic relationships and evolutionary traits in Ranunculus s.l. (Ranunculaceae) inferred from ITS sequence analysis. Mol. Phylogenet. Evol. 36, 305–327 (2005).
Emadzade, K., Lehnebach, C., Lockhart, P. & Hörandl, E. A molecular phylogeny, morphology and classification of genera of Ranunculeae (Ranunculaceae). Taxon. 59, 809–828 (2010).
Kelley, A. M., Johnson, P. G., Waldron, B. L. & Peel, M. D. A Survey of apomixis and ploidy levels among Poa L. (Poaceae) using flow cytometry. Crop Sci. 49, 1395–1402 (2009).
Richards, A. J. Plant Breeding Systems (London: Allen & Unwin, 2nd edition, 1997).
Hojsgaard, D., Klatt, S., Baier, R., Carman, J. G. & Hörandl, E. Taxonomy and biogeography of apomixis in angiosperms and associated biodiversity characteristics. Cr. Rev. Plant Sci. 33, 414–427 (2014).
Soreng, R. J., Davis, J. I., Voionmaa, M. A. A phylogenetic analysis of Poaceae tribe Poeae sensu lato based on morphological characters and sequence data from three plastid-encoded genes: evidence for reticulation, and a new classification for the tribe. Kew Bull. 425–454 (2007).
Chapman, G. P. (ed.). Reproductive Versatility in the Grasses (Cambridge University Press, 1990).
Rutishauser, A. Pseudogamie und Polymorphie in der Gattung Potentilla. Arch. Julius-Klaus-Srifr. Vererbungsforsch. 23, 267–424 (1948).
Asker, S. Pseudogamy, hybridization and evolution in Potentilla. Hereditas. 87, 179–184 (1977).
Harlan, J. R., de Wet, J. M. J. & On, O. Winge and a prayer: the origin of polyploidy. Bot. Rev. 41, 361–390 (1975).
Whitton, J., Sears, C. J., Baack, E. J. & Otto, S. P. The dynamic nature of apomixis in the angiosperms. Int J Plant Sci 169, 169–182 (2008).
Maugini, E. Morfologia fiorale embriologia ed embriog in Leontopodium alpinum Cass. var. α typicum fiori e Paoletti. Plant Biosystem. 69, 1–18 (1962).
Sokolowska-Kulczycka, A. Apomiksja u Leontopodium alpinum Cass: Apomixis in Leontopodium alpinum Cass. (1959).
Noyes, R. D. Apomixis via recombination of genome regions for apomeiosis (diplospory) and parthenogenesis in Erigeron (daisy fleabane, Asteraceae). Sex Plant Reprod. 19, 7–18 (2006).
Noyes, R. D. Inheritance of apomeiosis (diplospory) in fleabanes (Erigeron, Asteraceae). Heredity. 94, 193 (2005).
Noyes, R. D., Baker, R. & Mai, B. Mendelian segregation for two-factor apomixis in Erigeron annuus (Asteraceae). Heredity. 98, 92 (2007).
Noyes, R. D. & Rieseberg, L. H. Two independent loci control agamospermy (apomixis) in the triploid flowering plant Erigeron annuus. Genetics. 155, 379–390 (2000).
Stratton, D. A. Life history variation within populations of an asexual plant, Erigeron annuus (Asteraceae). Am. J. Bot. 78, 723–728 (1991).
Chiarugi, A. Aposporiae apogamia in Artemisia nitida Bertol. Nuovo Giorn. Bot. Ital. 33, 501–626 (1926).
Noyes, R. D. Apomixis in the Asteraceae: diamonds in the rough. Funct. Plant. Sci. Biotechnol. 1, 207–222 (2007).
Richards, A. J. The origin of Taraxacum agamospecies. Bot. J. Linn. Soc. 66, 189–211 (1973).
Firetti, F. Apomixis in Neotropical Vegetation. Vegetation. 129–148 (2018).
Roy, B. A. & Bierzychudek, P. The potential for rust infection to cause natural selection in apomictic Arabis holboellii (Brassicaceae). Oecologia. 95, 533–541 (1993).
Schranz, M. E., Dobeš, C., Koch, M. A. & Mitchell‐Olds, T. Sexual reproduction, hybridization, apomixis, and polyploidization in the genus Boechera (Brassicaceae). Am. J. Bot. 92, 1797–1810 (2005).
Orellana, M. R., Rovira, A. M., Blanché, C. & Bosch, M. Pollination and reproductive success in the gynodioecious endemic Thymus loscosii (Lamiaceae). Can. J. Bot. 83, 183–193 (2005).
Hand, M. L. & Koltunow, A. M. The genetic control of apomixis: asexual seed formation. Genetics. 197, 441–450 (2014).
Burt, A. Perspective: sex, recombination and the efficacy of selection – was Weismann right? Evolution. 54, 337–351 (2000).
Karger, D. N. et al. Climatologies at high resolution for the earth’s land surface areas. Sci. data. 4, 170122 (2017).
Janatková, K. et al. Community structure of soil phototrophs along environmental gradients in arid Himalaya. Environ. microbiol. 15, 2505–2516 (2013).
Krahulcová, A. & Suda, J. A modified method of flow cytometric seed screen simplifies the quantification of progeny classes with different ploidy levels. Biol. Plantarum. 50, 457–460 (2006).
Doležel, J., Greilhuber, J. & Suda, J. Estimation of nuclear DNA content in plants using flow cytometry. Nat. Protoc. 2, 2233–2244 (2007).
Doležel, J. et al. Plant genome size estimation by flow cytometry: inter-laboratory comparison. Ann. Bot-London. 82, 17–26 (1998).
Cornelissen, J. H. C. et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 51, 335–380 (2003).
Doležal, J. et al. Functionally distinct assembly of vascular plants colonizing alpine cushions suggests their vulnerability to climate change. Ann. Bot. 123, 569–578 (2019).
ter Braak, C. J. & Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.0. (Microcomputer power, 2012).
Acknowledgements
We thank K. Kunertová, for laboratory help. This research was supported by The Czech Science Foundation (grant no. GACR 13-13368S and 17-19376S).
Author information
Authors and Affiliations
Contributions
V.B. and J.D. were responsible for the overall direction of the research. The field data were collected by V.B., J.D. and P.K., V.B. and P.K. performed the cytometric analyses. J.D. collected plant trait data. The paper was written by V.B. with contributions from all authors.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Brožová, V., Koutecký, P. & Doležal, J. Plant apomixis is rare in Himalayan high-alpine flora. Sci Rep 9, 14386 (2019). https://doi.org/10.1038/s41598-019-50907-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-50907-5
This article is cited by
-
Engineering apomixis in crops
Theoretical and Applied Genetics (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
