
Abstract
We report the first occurrence of microfossils in Ediacaran strata of the Camaquã Basin. The assemblage includes simple (Leiosphaeridia sp. predominantly) and ornamented acritarchs associated with microbial mats. They are related to the Ediacaran Complex Acanthomorph Palynoflora (ECAP) and Late Ediacaran Leiosphere Palynoflora (LELP) due to the similar morphology and time interval assigned to those assemblages, though the observed specimens are a lot simpler and less diversified. However, different from the usual occurrences, this case study reports Neoproterozoic cosmopolitan communities living in marine (basal unit) and lacustrine (middle units) settings. Fossils within non-marine strata in the Precambrian record are rare. Therefore, this first finding of microfossils in the Camaquã Basin constitutes a new piece of the puzzle related to the history of the Panafrican-Brasiliano basins and shed some light on possible settings where the Ediacaran eukaryotes have evolved.
Similar content being viewed by others


Introduction
Precambrian sedimentary basins have been intensively investigated all around the world in terms of eukaryotic protists that arise in the Late Paleoproterozoic Era but intensively diversify during the Ediacaran Period. The Proterozoic life is generally regarded as dominated by marine and intertidal biota preserved in both carbonate1,2 and siliciclastic3,4 rocks. However, a few studies also indicate that Precambrian land surfaces housed biota in paleokarst5, lateritic paleosols6, lacustrine7 and subaerial non-marine settings8. Actually, non-marine, organic-walled microfossils identified as acritarchs were first described long ago, in a Torridonian sequence of northwest Scotland9, and were later refined by other authors10,11.
Given the particular importance of Neoproterozoic biota for the comprehension of the spreading of life across diverse ecological niches, we register the first occurrence of Ediacaran, organic-walled microfossils preserved in fine-grained siliciclastic strata of the Camaquã Basin, southernmost Brazil. Furthermore, dealing with an Ediacaran succession, most of the forms are simple and low-diversity, but related to both marine and lacustrine settings, hence representing cosmopolitan microorganisms.
Results
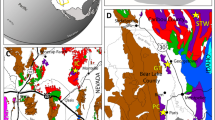
Even though the Camaquã Basin (Fig. 1) ichnological content is studied since the 1990’s12,13,14,15, this is the first record of body fossils on it. Microfossils occur in siliciclastic mudstones of the Maricá, Bom Jardim and Santa Bárbara groups (Fig. 2). This fossil record comprises abundant acritarchs, identified as simple and ornamented forms and remains of bacterial mats.

Tectono-sedimentary evolution of the Camaquã Basin (modified from 12). White stars represent stratigraphic distribution of the sampled outcrop sets (see Supplementary File): 1- Outcrop set 1: Maricá Group – Santa Bárbara sub-basin; 2 – Outcrop set 2: Santa Bárbara Group - Santa Bárbara sub-basin, base; Outcrop set 3: Santa Bárbara Group- Santa Bárbara sub-basin, top; Outcrop set 4: Bom Jardim Group – Guaritas sub-basin; Outcrop set 5a: Bom Jardim Group - Piquirí sub-basin; Outcrop set 5b: Maricá Group – Piquirí sub-basin. Radiometric ages are assigned to specific authors in Paim et al.12.

Photomicrograph of sedimentary features and microfossil content. (A) Fine-grained facies of Maricá Group seen under natural light showing a sphaeromorph acritarch (red arrow), filamentous bacterial mats (blue arrows), and carbonate cement around the grains; (B) Fine-grained facies form Maricá Group seen under polarized light showing a sphaeromorph acritarch (red arrow), grains trapped (yellow arrows) by filamentous bacterial mats (blue arrows), and carbonate cement around the grains (green arrows). (C) Simple spherical microfossil (red arrow) and silt grains involved by filamentous bacterial mats (blue arrow) in the fine-grained facies of the Bom Jardim Group; (D) Siltstone of the Santa Bárbara Group displaying aligned grains of mica (yellow arrow) and opaque rounded microfossil (red arrow) inside a fossilized microbial mat.
Siltstones and very fine-grained sandstones of the Maricá Group are immature and represent storm deposits. Grains are angular to subangular and eventually entrapped by filamentous microbial mats (Fig. 2A,B). Different from the other units, they do not present eodiagenetic hematite coating, but only poikilotopic calcite cement. Although also immature, siltstones and very fine-grained sandstones of both Bom Jardim and Santa Bárbara groups record hyperpycnal turbidity currents. Grains are also angular to subangular, but contain eodiagenetic hematite coatings precipitated before carbonate cementation has taken place (Fig. 2C,D).
Systematic palaeontology
Repository
Described palynological slides are kept in the Museu de História Geológica do Rio Grande do Sul - MHGEO – Unisinos, São Leopoldo, RS, Brazil – and assigned numbers according to the collection (ULVG – UNISINOS/Laboratório de História da Vida e da Terra - Lavigea). Location and geographic coordinates for each sample are presented in the Supplementary File.
Incertae sedis
Group Acritarch Evitt, 1963
Genus Leiosphaeridia, Eisenack, 1958, emend. Turner, 1984,
Type species Leiosphaeridia baltica, Eisenack, 1958
Leiosphaeridia spp.
Figures 3A–C and 4A–C/F
Occurrence and material
Slides ULVG 12495, 12496, 12505, 12506, 12507, 12508, 12511, 12513, 12514, 12517, 12518, 12519, 12520. One hundred forty nine complete specimens and dozens of fragments of sphaeromorphs were extracted from mudstones of the Maricá, Bom Jardim and Santa Bárbara outcrops (see Supplementary File - Table ST1).
Description
Organic-walled, acid-resistant, simple spherical to sub-spherical, compressed, commonly folded vesicles with smooth to shagrinate or granular wall surface. Diameter ranging from 20 to 150 µm.
Remarks
Some species show excystment opening, median split or partial rupture16. Leiosphaeridia is a long-ranging genus, ranging from Paleoproterozoic until present. This genus is the most abundant microfossil in the geologic record, with a great variability of vesicle dimensions17,18. The group comprises four basic species based on the wall thickness and size class: L. minutissima – thin-walled, <70 µm; L. tenuissima – thin-walled, 70–200 µm; L. crassa – thicker-walled, <70 µm; L. jacutica – thicker-walled, 70–800 µm19.
Discussion
The Camaquã Basin palynomorphs includes more than one leiosphaerid species, but it is impossible to assert which species because the poor preservational quality precludes the identification of wall internal details and thickness. Based on the size of the recovered specimens it is possible to associate then with two potential species: L. minutissima and L. crassa. However, the lack of diagnostic features about wall-thickness within this group of acritarchs makes difficult to ascribe them to species level.
Genus Tanarium Kolosova, 1991,
emend. Moczydlowska, Vidal and Rudavskaya, 1993
Type species Tanarium conoideum Kolosova, 1991,
emend. Moczydlowska, Vidal and Rudavskaya, 1993
Description
Organic-walled, acid-resistant microfossils consisting of spherical to sub-spherical vesicle bearing numerous hollow, cylindrical processes distributed around the vesicle outline. The processes are more or less of equal size (in a single specimen), differing in number between specimens and not very evenly distributed. Diameter ranging from 80 to 200 µm.
Tanarium irregulare Moczydlowska, Vidal and Rudavskaya, 1993
Figure 3F
Occurrence and material
Slides ULVG 12487, 12493, 12514, 12515. Six complete and dozens of fragmented vesicles appear in palynological material extracted from mudstones of the Maricá, Bom Jardim and Santa Bárbara outcrops (see Supplementary File - Table ST1).
Description
Organic-walled, acid-resistant microfossils consisting of originally spherical to sub-spherical vesicles irregular in outline ornamented with long, tubular and conical, simple and branching protrusions. Diameter ranging from 50 to 150 µm and spines ranging from 10 to 20 µm.
Remarks
The specimens are associated with diagnosis characteristics of Tanarium irregular sp.20,21. The name derives from Latin irregularis – irregular, referring to the processes shape.
Genus Lophosphaeridium Timofeev, 1959,
emend. Downie, 1963, emend. Lister, 1970
Type species Lophosphaeridium rarum Timofeev, 1959,
emend. Downie, 1963
Lophosphaeridium sp.
Figure 3 D-E and 4D
Occurrence and material
Slides ULVG 12494, 12506, 12507, 12509, 12510, 12511, 12513, 12515. Twenty-six entire and dozens of fragmented specimens extracted from mudstones of the Maricá, Bom Jardim and Santa Bárbara outcrops (see Supplementary File - Table ST1).
Description
Organic-walled, acid-resistant vesicle, circular to sub-circular in outline. Compactional folds present on some specimens. The surface of the vesicle displays numerous tightly arranged, small conical spines that are visible as evenly small hairs distributed around the vesicle outline. Vesicle diameter ranging from 50–150 µm. Conical projections are approximately 10–15 µm long.
Remarks
The studied specimens are comparable to Lophosphaeridium sp.22,23,24 regarding their morphological features and size, but it not possible to ascribe them to species level.
Genus Germinosphaera Mikhailova, 1986, emend. Butterfield, 1994
Type species Germinosphaera bispinosa Mikhailova, 1986
Germinosphaera sp.
Figures 3G–J and 4E
Occurrence and material
Slides ULVG 12487, 12498, 12503, 12505, 12494, 12506, 12507, 12509, 12510, 12511, 12513, 12515, 12517, 12518, 12519, 12520. Forty-two microfossils extracted from mudstones of Maricá, Bom Jardim and Santa Bárbara outcrops (see Supplementary File - Table ST1).
Description
Organic-walled, acid-resistant spheroidal vesicles with one or more open-ended, tubular and occasionally branched processes that communicate freely with the vesicle. Bodies 70–100 µm long and 30–70 µm wide, occasionally with a vase-shaped form with short or elongated processes, 5–15 µm wide and 10–50 µm.
Remarks
All complete specimens of Germinosphaera sp.25,26 found in the analysed samples bear a single filamentous process.
Discussion
Fine-grained facies of the Camaquã Basin show organic-walled microfossils identified as simple and ornamented acritarchs as well as bacterial mats (filamentous and coccoidal). These organisms occur along the three basal units of the Camaquã Basin: Maricá, Bom Jardim and Santa Bárbara groups. Each unit record a distinct tectonic setting, which led to different environmental conditions27 (Fig. 1). The fine-grained strata of the Maricá Group record a short-distance transport of immature, angular to sub-angular sediments deposited under wave action in a shallow marine setting (See Supplementary File). No micro (hematite coating) or macroscopic (desiccation cracks) features related to subaerial exposure were identified. Similarly, both Bom Jardim and Santa Bárbara groups also comprise immature, short-distance transported sediments deposited in a subaqueous realm. However, a much larger alluvial influence (relative to the Maricá Group), and associated hyperpycnal turbidity currents, and rare (Bom Jardim Group) to common (Santa Bárbara Group) subaerial exposure of the depositional surface suggest continental, deep- to shallow lacustrine settings (See Photographs in Supplementary File). Accordingly, eodiagenetic hematite coatings are widespread in both units.
Camaquã Basin microfossils can be associated with Ediacaran Complex Acanthomorph Palynoflora (ECAP)28 in aspects of time range and with Late Ediacaran Leiosphere Palynoflora (LELP)29 when observed morphological aspects. This association still needs an improvement because nor the time range distribution or the morphology of the organisms are equal of the described assemblages. Camaquã Basin microfossils seems to represent a new group of microfossils (Fig. 3).

Acritarchs view under transmitted light microscopy. (A–C) Leiosphaeridia sp. from (A) Maricá (sample ULVG 12518), (B) Bom Jardim (sample ULVG 12495), and (C) Santa Bárbara (sample ULVG 12511) groups; (D,E) Lophosphaeridium sp. from (D) Maricá (sample ULVG 12518) and (E) Bom Jardim (sample 12495) groups; (F), Tanarium irregulare from Santa Bárbara Group (sample 12487); (G–J), Germinosphaera sp. from (G,H) Maricá (sample ULVG 12509), original opaque (G) and illuminated (H) image, (I) Bom Jardim (sample ULVG 12506), and (J) Santa Bárbara (sample ULVG 12515) groups.
The same assemblage was recovered in both marine (Maricá Group) and lacustrine settings (Bom Jardim and Santa Bárbara groups). Although acritarchs have usually been associated with marine settings, this taxonomic overlap is frequent among microfossil taxa from marine and non-marine setting in the Precambrian9,10,30 and reinforces the idea that morphological simple genera such as Leiosphaeridia not necessarily indicate any particular environment11. Some species of Leiosphaeridia may be related to the Prasinophyceae31,32, but the simple morphology of this genus does not preclude a wide variety of natural affinities and sources, both marine and non-marine30. Besides, organic-walled sphaeromorphs were also found in non-marine Torridonian rocks of Scotland9,11. Other works described similar organisms in non-marine strata and proposed a Proterozoic terrestrial colonization7,8,33 as well as a continental evolution for the Eukarya during the Late Ediacaran11.
The recovered microfossils were always associated with microbial mat fragments, as demonstrated in palynological slides (Fig. 4) and thin sections (Fig. 2). This coincidence reinforces previous suggestions that many Ediacaran acritarchs could represent benthic and even heterotrophic life stages30,31,32. We understand that this close association is not a definitive proof. There is the possibility of planktonic organisms sinking on the bottom and being trapped by bacterial mats. However, why microfossils do not occur when microbial mats are absent? Therefore, this close association does suggest a co-existence of both microfossils and microbial mats. Searching for favourable conditions and food, in a stable sea or lacustrine floor, with no signs of transport, these microorganisms inhabit the same subaqueous environment where microbial colonies build up mats.

Acritarchs view on scanning electron microscopy. (A,B) Leiosphaeridia sp. from Maricá Group (sample ULVG 12509); (C,D) Leiosphaeridia sp. (C) and Lophosphaeridium sp. (D) with filamentous bacteria from Bom Jardim Group (sample ULVG 12506); (E,F) Germinosphaera sp. (E) and Leiosphaeridia sp. (F) from Santa Bárbara Group (sample ULVG 12508) with bacterial mats fragments. Note folded microfossils due to compaction (B,C images).
We demonstrate that the first find of body fossils in the Camaquã Basin add new information about the Ediacaran fossil record of South America and fill some gaps about microfossils existence and distribution in a Proto-Gondwana context. Several Ediacaran basins as Nama (Namibia), Arroyo del Soldado (Uruguay), Corumbá (Brazil) and others register body fossils related to ECAP and ELP assemblages. Even simpler and less diversified that those assemblages, the Camaquã Basin microfossils reported here include Camaquã Basin as hostess of Ediacaran life at southwestern Proto-Gondwana.
The recovery of microfossils in marine and lacustrine strata and the profusion of Leiosphaerids suggest that these polyphyletic long-ranging taxa are cosmopolitan, as suggested by previous works11,33,34. Besides, the lack of macrofossils of the Ediacara biota suggests restraining ecological conditions (large fluvial inflow) for both marine and lacustrine settings developed during basin evolution. In addition, the Ediacara macrofossils occur in Late Ediacaran strata (575–542 Ma) around the world1. This time interval is compatible with the Santa Bárbara Group, which reflect shallow-lacustrine conditions, favourable for cosmopolitan microrganisms.
Conclusions
The first finding of body fossils in the Camaquã Basin adds new evidence about the Ediacaran fossil record of South America, improves the dataset and complements the scenario of life within the Proto-Gondwana. Based on their similar morphology and assigned time interval the reported microfossils could be ascribed to the Ediacaran Complex Acanthomorph Palynoflora (ECAP) and Late Ediacaran Leiosphere Palynoflora (LELP), even though the observed specimens are simpler and less diversified.
The finding of microfossils in marine and lacustrine strata and the profusion of Leiosphaerids suggest that both habitats were already colonized by cosmopolitan eukaryotes by the Late Ediacaran. On the other hand, the lack of typical components of the Ediacara fauna suggest restrictive ecological conditions in both marine and lacustrine realms during the Camaquã Basin evolution. At last, the close relationship between microfossils and bacterial mats suggest a possible link between both components of the Camaquã Basin biota and their living conditions.
Material and Methods
This study includes forty-three palynological slides obtained from outcrop samples. The outcrops record distinct intervals sampled at different locations (see Supplementary File - ST1). All samples are stored in the collection of the Museu de História Geológica do Rio Grande do Sul of the Universidade do Vale do Rio dos Sinos (São Leopoldo, Brazil). The palynological preparation technique was used for the extraction of acid-insoluble microfossils. Following Grey’s technique35, raw samples were mechanically disaggregated. After, they were digested with HCl and HF for carbonate and silicate removal, respectively. Boiling HCl was used for the removal of clay minerals. Strewed kerogen was oxidized with concentrated HNO3. After filtration (10 µm filter size) and swirling to separate heavy minerals, strew slides were prepared and examined under transmitted light microscope with interference contrast (Zeiss Axio Imager-A2). SEM-EDS (scanning electron microscopy plus energy-dispersive X-ray spectroscopy) analyses were performed on gold-coated samples obtained from representative microfossil specimens. SEM studies were executed at Instituto Tecnológico de Micropaleontologia - itt FOSSIL - of the Universidade do Vale do Rio dos Sinos – UNISINOS (São Leopoldo City/Rio Grande do Sul State), using a Zeiss EVO/MA15 SEM equipment, at Laboratório de Conformação Nanométrica – Instituto de Física of the Universidade Federal do Rio Grande do Sul – UFRGS (Porto Alegre/Rio Grande do Sul), using a JIP-4500 MultiBeam SEM-FIB equipment and at Laboratório Nacional de Nanotecnologia – LNNano - of the Centro Nacional de Pesquisa em Energia e Materiais – CNPEM (Campinas/São Paulo), using a Quanta 650FEG SEM equipment.
Data Availability
All data generated or analysed during this study are included in this published article and in the Supplementary File.
References
Schopf, J. W. & Klein, C. The Proterozoic Biosphere. 348p. (Cambridge Univ. Press., 1992).
Grotzinger, J. P. W. & Knoll, A. H. Stromatolites in Precambrian Carbonates: Evolutionary Mileposts or Environmental Dipsticks? Annu. Rev. Earth Planet. Sci. 1999, 27:313–58 (1999).
Hagadorn, J. W. & Bottjer, D. J. Wrinkle structures: microbially mediated sedimentary structures common in subtidal siliciclastic settings at the Proterozoic Phanerozoic transition. Geology 25, 1047–1050 (1997).
Gehling, J. G. Microbial mats in terminal Proterozoic siliciclastics: Ediacaran death masks. PALAIOS 14, 40–57 (1999).
Beeunas, M. A. & Knauth, L. P. Preserved stable isotopic signature of subaerial diagenesis in the 1.2-b.y. Mescal Limestone, central Arizona: Implications for the timing and development of a terrestrial plant cover. Geological Society of America Bulletin 96, 737–745 (1985).
Strother, P. K. & Wellman, C. H. Palaeocology of a billion-year-old non-marine cyanobacterium from the Torridon Group and Nonesuch Formation. Palaeontology 59, 89–108 (2016).
Horodyski, R. J. & Knauth, L. P. Life on land in the Precambrian. Science 263, 494–498 (1994).
Prave, A. R. Life on land in the Proterozoic: evidence from the Torridonian rocks of northwest Scotland. Geology 30, 811–814 (2002).
Teall, J. J. H. The Geological Structure of the North-west Highlands of Scotland (eds Peach, B. N. et al.) 288, plate LII (Memoirs of the Geological Society of Great Britain, 1907).
Cloud, P. & Germs, A. New Pre-Paleozoic Nannofossils from the Stoer Formation (Torridonian), Northwest Scotland. GSA Bulletin 82(12), 3469–3474 (1971).
Strother, P. K., Battison, L., Brasier, M. D. & Wellman, C. H. Earth’s earliest non-marine eukaryotes. Nature 473, 505–509 (2011).
Netto, R. G., Paim, P. S. G. & Martini-da-Rosa, C. L. Informe preliminar sobre a ocorrência de traços fósseis nos sedimentitos basais das bacias do Camaquã e Santa Bárbara. In Workshop Sobre Bacias Molássicas Brasilianas, 1, 1992. Anais, São Leopoldo, 90–96 (1992).
Paim, P. S. G., Faccini, U. F., Netto, R. G. & Nowatzki, C. H. Estratigrafia de sequências e sistemas deposicionais das bacias do Camaquã e Santa Bárbara, Eo-Paleozóico do RS (Brasil). INSUGEO, Serie Correlación Geológica 9, 41–55 (1992).
Netto, R. G. Preliminary report about trace fossils in Precabrian/Cambrian deposits of Southern Brazil. In Circum-Atlantic Circum-Pacific Terrane Conference 2, 1994, Program and Abstracts, p. 30 (1994).
Netto, R. G. Paleoicnologia do Rio Grande do Sul. In: M. Holz and L. F. De Ros. CIGO/UFRGS, 25–43 (2000).
Evitt, W. R. A discussion and proposals concerning fossil dinoflagellates, hystrichospheres and acritarchs - I, II. Proceedings of the Natural Academy of Sciences of the United States of America 49, 298–302 (1963).
Eisenack, A. Tasmanites Newton 1875 und Leiosphaeridia n. g. als gattungen der Hystrichosphaeridea. Palaeontographica 110, 1–19 (1958).
Turner, R. E. Acritarchs from the type area of the Ordovician Caradoc Series, Shropshire, England. Palaeontographica 190, 87–157 (1984).
Javaux, E. J., Knoll, A. H. & Walter, M. R. TEM evidence for eukaryotic diversity in mid-Proterozoic oceans. Geobiology 2, 121–132 (2004).
Kolosova, S. P. Late Precambrian thorny microfossils of the east of the Siberian Platform. Algologia 39, 53–59 (1991).
Moczydłowska, M., Vidal, G. & Rudavskaya, V. A. Neoproterozoic (Vendian) phytoplankton from the Siberian Platform, Yakutia. Palaeontology 36, 495–521 (1993).
Timofeev, B. V. The ancient Baltic flora and its stratigraphical significance: Vsesoyuznyi Neftyanoi Nauchno-Issledovatelskii Geologorazvedochnyi Institut, Leningrad (VNIGRI). Trudy 129, 136 (1959).
Downie, C. & Sarjeant, W. A. S. On the interpretation and status of some hystrichosphere genera. Palaeontology 6, 88–96 (1963).
Lister, T. The acritarchs and chitinozoa from the Wenlock and Ludlow series of the Ludlow and Millichope areas, Shropshire. Monograph of the Palaeontographical Society 124(528), 1–100 (1970).
Mikhailova, N. S. New finds of the microfossils from the Upper Riphean deposits of the Krasnoyarsk region, p. 31–37. In: Current Problems of Modern Paleoalgology (Nauka, Kiev., 1986).
Butterfield, N., Knoll, A. H. & Swett, K. Paleobiology of the Neoproterozoic Svanbergfjellet Formation, Spitsbergen. Fossils and Strata 34, 1–84 (1994).
Paim, P. S. G., Wildner, W. & Chemale, F. Jr. Estágios evolutivos da Bacia do Camaquã (RS). Ciência e Natura 1–36 (2014).
Grey, K., Walter, M. R. & Calver, C. R. Neoproterozoic biotic diversification: Snowball Earth or aftermath of the Acraman impact? Geology 31, 459–462 (2003).
Gaucher, C. & Sprechmann, P. Neoproterozoic acritarch evolution. In: Gaucher, C., Sial, A. N., Halverson, G. P., Frimmel, H. E. (Eds): Neoproterozoic-Cambrian Tectonics, Global Change and Evolution: a focus on southwestern Gondwana. Developments in Precambrian Geology, 16, 319–326 (2009).
Strother, P. K. Acritarchs. In: Jansonius, J. and McGregor, D. C. (eds), Palynology: principles and applications. American Association of Stratigraphic Palynologists Foundation 1, 81–106 (1996).
Wicander, E. R. Upper Devonian-Lower Mississippian acritarchs and prasinophycean algae from Ohio. USA Palaeontographica, Abteilung B. 148(9–43), 5–19 (1974).
Tappan, H. The paleobiology of plant protists. San Francisco. 1028 p. (W.H. Freeman & Co., 1980).
Porter, S. M. & Knoll, A. H. Testate amoebae in the Neoproterozoic Era: evidence from vase-shaped microfossils in the Chuar Group, Grand Canyon. Paleobiology 26, 360–385 (2000).
Moczydłowska, M. Algal affinities of Ediacaran and Cambrian organic-walled microfossils with internal reproductive bodies: Tanarium and other morphotypes. Palynology 40(1), 83–121 (2016).
Grey, K. A modified palynological preparation technique for the extraction of large Neoproterozoic acanthomorph acritarchs and other acid insoluble microfossils. 23 p. (Geological Survey of Western Australia Record, 1999).
Acknowledgements
This research was supported by the FAPERGS 01/2017 ARD Project entitled Geobiology of the Ediacaran Camaquã Basin (Brazil) (Grant Number 17/2551-0000 824-3). The first author thanks CAPES for her scholarship (Grant Number 88887.150701/2017-00). The third author acknowledges CNPq for the long-term support (Grant Number 308140/2014-1). The authors are grateful to UNISINOS, UFRGS and CNPEM for providing lab facilities (SEM and X-ray) and field equipment. We thank Dr. Sebastian Willman (Uppsala University) for the discussions about the palynological material; Dr. Mírian Pacheco (UFSCar) and Dr. Farid Chemale Jr. for valuable assistance and review of a first draft of this MS. The authors also thank the editor for the assistance.
Author information
Authors and Affiliations
Contributions
I.L. contributed with sedimentology and palaeontology research, R.S.H. contributed with palaeontology data and P.S.G.P. contributed with geology and sedimentology research and review.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lehn, I., Horodyski, R.S. & Paim, P.S.G. Marine and non-marine strata preserving Ediacaran microfossils. Sci Rep 9, 9809 (2019). https://doi.org/10.1038/s41598-019-46304-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-46304-7
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
