
Abstract
The Gulf of Naples is an example of the most beautiful and biodiverse marine regions of the Mediterranean Sea and of the most impacted areas in terms of industrial activities, large contaminated areas, resource exploitation, infrastructures at sea and maritime transportation. We conducted Remotely Operated Vehicle surveys in the Dohrn Canyon in the Tyrrhenian Sea at approximately 12 NM off Naples metropolitan area, and revealed a hotspot of deep-sea benthic biodiversity of sessile fauna at ca. 400 m depth. The hard bottoms are characterized by a high abundance of charismatic species, such as the habitat forming cold-water corals (CWC) Madrepora oculata, Lophelia pertusa, Desmophyllum dianthus in association with the large size bivalves Acesta excavata and Neopycnodonte zibrowii. This CWC-bivalve co-occurrence represents a novel biotope for the Mediterranean Sea, which coexists with the evidence of severe anthropogenic threats, such as illegal dumping and fishery malpractices that were visually documented during the survey. We recommend the adoption of specific protection measures to preserve these unique deep-sea assemblages showing the uncommon co-existence of such a number of deep-sea species in a single habitat.
Similar content being viewed by others


Introduction
In the 19th century, the beauty and biological richness of the Gulf of Naples (GoN) was such to convince Anton Dohrn to found there in 1872 the first marine station in the world to demonstrate the validity of the Darwin theories on evolution using marine organisms. The following 150 years have witnessed the unbridled growth of scientific studies in the gulf, along with the explosion of the industrial and urban development of the city of Naples and adjacent coast, especially after World War II1, which determined a progressive increase of the anthropogenic pressures. Today, the Naples metropolitan area accounts for ca. 4.5 million inhabitants and the entire population of the gulf is distributed along a mere 12 km coastal stretch. The presence of illegal dumping and untreated sewage, along with the presence of wide contaminated areas, intense maritime transportation, infrastructure at sea, other direct and indirect anthropogenic stressors2,3,4,5,6,7,8,9,10,11,12,13 have progressively threatened the coastal ecosystems of this gulf, determining also a progressive loss of marine habitats.
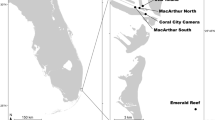
Despite a long tradition of biological studies that renders the GoN one of the most intensively investigated marine areas of the world, little is known about the ecology and environmental status of the deep-sea habitats of the gulf, whose hard bottom has been so far investigated only in terms of geological setting. A prominent submarine feature of the gulf is the Dohrn Canyon, a bifurcate structure (Fig. 1) that indents perpendicular to the coastline the continental shelf 12 NM from the Naples megalopolis, beginning at ca. −250 m and sharply declining down to ca 1300 m in the Tyrrhenian plain14,15. This inactive canyon, part of the Magnaghi-Dohrn canyon system16 and consisting of a western and an eastern branch, is thought to have been formed in response to relative sea-level fall during the last glaciation enhanced by local subsidence, cutting through Pleistocene sediments and the Campana Ignimbrite14. The water mass structure of the GoN is predominantly linked to the main circulation of the southern and mid-Tyrrhenian Sea, with influences from local factors, such as the wind stress and the river runoff. The two main water masses flowing in the GoN are the Modified Atlantic Water (MAW) that occupies the upper 50–100 m and the Levantine Intermediate Water (LIW) located below ~200–300 m, with salinity of 38.65, temperature of 14.2 °C and a potential density (σθ) of 29.0 kg m−3 17,18,19. Depending on the season, other water masses can be recognized within the GoN, such as the Tyrrhenian Intermediate Water (TIW) formed during winter mixing at depths down to ~150 m and the Tyrrhenian Surface Water (TSW) found above 75 m as the result of summer warming and freshening of the TIW20.

(A) Location map of the Gulf of Naples (white star in the inset. Basemap data: Google, SIO, NOAA, US Navy, NGA, GEBCO; Image Landsat/Copernicus) showing the three main canyons dissecting the Campanian shelf. Bathymetry from52 (http://www.emodnet-bathymetry.eu/), contour lines spacing 100 m; (B) detail of Dohrn Canyon (red square in A) and location of the four ROV dives performed in the area. (C) ROV tracks plotted over the morphobathymetric map. Note the sector of the canyon wall between 350 and 470 m depth where deep-water corals and large bivalves do occur. Contour line spacing 5 m. Maps were created using ArcGIS software by Esri (www.arcgis.com). ArcGIS and ArcMap are the intellectual property of Esri. Morphobathymetry were generated using CARIS HIPS and SIPS release 9 software (http://www.caris.com).
In 2016, the first ecological exploration of the deep-sea habitats of the Dohrn Canyon was launched by means of Remotely Operated Vehicle (ROV) focusing on the steep walls of the northern flank of its western branch (Fig. 1B,C). The survey unveiled for the first time the presence of charismatic benthic life in the canyon system, with biological traits previously unreported for the entire Mediterranean basin.
Results
The living community
An abundant and diverse sessile fauna was observed by ROV image analysis (Figs 2–4). Several macro-invertebrates species were concentrated on rocky bottoms at depths of 350–470 m (Fig. 5), corresponding to the depth of the LIW (Fig. 6). We report here the presence of the frame-building cold-water colonial scleractinian corals Lophelia pertusa and Madrepora oculata. These species are key ecosystem engineers in deep-sea habitats of the Atlantic Ocean and have been reported with a fragmented distribution in the Mediterranean Sea21,22 but were never reported previously in the GoN. Differently from what observed so far in the Mediterranean Sea, where cold-water coral habitats are predominantly characterized by M. oculata, in the Dohrn Canyon L. pertusa is the most abundant species, especially on the northwestern flank where as many as 10 colonies of each species have been counted on a distance of 235 m between 375–455 m water depth. Our ROV images and videos indicate that both species occur as small interspersed colonies on the rocky substrate (Figs 2A,B,E and 3B,E), isolated or admixed (Fig. 2F), and with polyps fully expanded (Figs 2A,B,F and 3B,E). On the canyon walls, we also found conspicuous populations of solitary corals (Figs 2C–E and 3B,D,F), including Javania cailleti and Desmophyllum dianthus (a target species for paleoclimatically-oriented studies23). Such solitary corals settle on bedrock overhangs forming dense aggregations of up to more than 700 ind. m−2, with the tentacles facing down, and often unevenly rimming the layered substrate (Fig. 2D). The sea anemone Protanthea simplex is equally abundant (Fig. 2C), often associated with solitary corals or settled on the dead frames of M. oculata and L. pertusa. This piezotolerant species, spanning from infralittoral to bathyal depths in the northern Atlantic, has been previously reported only once in the Mediterranean, in the Strait of Sicily24. Another unique finding is the discovery of the presence of large bivalves in association with such CWC (Fig. 5; Table 1), namely the limid Acesta excavata (Figs 2E and 3A,D), and the giant deep-sea oyster Neopycnodonte zibrowii (Fig. 4A), which colonize the vertical cliff of the canyon. These large bivalves are frequently reported in the Atlantic Ocean, but seldom documented alive in the Mediterranean Sea25,26,27, where their presence has been so far documented only in the western basin or as Pleistocene fossil in the eastern basin. Individuals attain a size of ca. 20 cm that is comparable with the NE Atlantic populations28. Individuals of A. excavata have been observed byssate on the wall at 380 m water depth down to the basal part of the canyon flank at 415 m. The long-lived oyster N. zibrowii is recorded in all ROV transects along the Dohrn Canyon (Figs 3A and 4A) but with variable density, occurring as individuals or aggregations (strings) in a depth range of 375–415 m. Remarkably, clusters of dead and black-coated N. zibrowii occur cemented at the canyon’s wall, some of which fouled by living epifauna that includes CWC. Overall we counted more than 120 living N. zibrowii reaching a maximum density of ca. 20 ind. m−2 in the northwestern part of the canyon between 375–395 m (Fig. 4B). The canyon’s wall is inhabited by a number of other mega- and macrobenthic taxa including further cnidarians, sponges, polychaetes, molluscs, bryozoans, echinoderms, crustaceans, adding to the total biodiversity (see species list in Table 2). Other cnidarians include rare specimens of the antipatharian Parantipathes larix (Fig. 3C), the sea fan anemone Amphianthus dohrnii (Fig. 2F) abundant on long lines entangled in the wall, and an unidentified species of Epizoanthus sp. (Fig. 3E). The sponge assemblages are dominated by the encrusting species Desmacella inornata and Hexadella sp., and by a few massive and tubular sponges (i.a., Spongosorites flavens). Serpulid polychaetes, encrusting and erect bryozoans, byssate (Asperarca nodulosa, Delectopecten vitreus) and cemented (Spondylus gussonii) bivalves contribute to the high biodiversity of hard substrates of the canyon. The investigated system hosts also some vagrant macro-invertebrate species, such as the pencil urchin Cidaris cidaris (Fig. 3A), Gracilechinus cf. acutus (Fig. 3B), the asteroid Astropecten sp., and the decapods Munida intermedia (Fig. 3F) and Anamathia rissoana (Fig. 2F). The colonial coral framework provides a cryptic habitat populated by a diversified fauna such as the pink fish Bellottia apoda and unidentified crustaceans.

Main frame coral builders and most noticeable fauna in the Dohrn Canyon: (A) the colonial scleractinian Lophelia pertusa with expanded polyps; arrows indicate Acesta excavata on the right foreground and the solitary coral Javania cailleti on the left foreground (st. MS16_7, −386 m) bar = 2 cm; (B) live colony of the scleractinian Madrepora oculata (st. MS16_7, −391 m, bar = 2 cm); (C) solitary coral community, with adult and juvenile Desmophyllum dianthus (d), J. cailleti (j), the sea anemone Protanthea simplex (arrows), plus on the left side a juvenile L. pertusa (l) (st. MS16_7, −385 m) bar = 1 cm; (D) curtains of solitary corals (Desmophyllum dianthus, Javania cailleti) rimming the layered bedrock (st. DOHRN_ROV_1, −385 m) bar = 1 cm; (E) the large limid A. excavata among solitary (D. dianthus: white arrow) and colonial (M. oculata) scleractinian corals, both alive and dead (st. MS16_7, −415 m) bar = 10 cm; (F) mud-draped substrate with small colonies of M. oculata (m) and L. pertusa (l), arrow indicates a group of small anemone (Amphianthus dohrnii) above the decapod Anamathia rissoana crawling on a undefined stalk (st. MS16_7, −412 m) bar = 1 cm.

Details of the fauna inhabiting the Dohrn Canyon: (A) the sea-urchin Cidaris cidaris crawling next to a live Acesta excavata, note a living Neopycnodonte zibrowii to the right, plus various solitary corals (Desmophyllum dianthus) and Madrepora oculata in the background (st. MS16_7, −386 m) bar = 5 cm; (B) live and dead M. oculata colonies, D. dianthus, A. excavata, and the echinoid Gracilechinus cf. acutus (st. MS16_7, −384 m) bar = 5 cm; (C) the antipatharian Parantipathes larix (st. MS16_7, −406 m) bar = 5 cm; (D) image documenting the taxonomic complexity of the sessile fauna in the canyon: A. excavata (a), M. oculata (m), L. pertusa (l), D. dianthus (d) and J. cailleti (j), arrow points the decapod Anamathia rissoana (st. DOHRN_ROV_1, −377 m) bar = 10 cm; (E) the solitary coral D. dianthus with fully expanded polyp next to a colony of L. pertusa, note also the colonial sea anemone Epizoanthus sp. colonizing a lost longline (st. MS16_7, −391 m) bar = 5 cm; (F) coral assemblage dominated by J. cailleti (j), small colony of M. oculata (m) and the decapod Munida intermedia sheltered among corals, arrow indicates juvenile specimens of M. intermedia (st. DOHRN_ROV_1, −384 m) bar = 2 cm.

Dohrn Canyon: (A) Neopycnodonte zibrowii string settling on the emerging layered bedrock (st. MS17_102) −446 m; (B) mud-draped bedrock with dead and blackened cemented shells of N. zibrowii, note an undetermined Ophidiasteridae seastar on top (st. MS16_7, −388 m) bar = 10 cm; (C) the sponge Stylocordyla pellita stemming from the canyon’s muddy bottom (st. MS16_7, −435 m) bar = 1 cm; (D) dense aggregation of Pachycerianthus cf. dohrni, note longline trapped among the cnidarians (st. MS16_7, −411 m) bar = 5 cm.

(A) Habitat map based upon four ROV surveys carried out in the western branch of the Dohrn Canyon with respect to the most relevant biological taxa and marine litter observed there; it documents a widespread distribution of Neopycnodonte zibrowii in the canyon head at steeper loci at the sites DOHRN-ROV_1/MS16_7 and MS17_102 but only sporadic at MS17_81; co-occurring CWC presence (Lophelia pertusa, Madrepora oculata, Desmophyllum dianthus) is less obvious loci at the site MS17_102; Acesta excavata is relatively frequent at the sites DOHRN-ROV_1/MS16_7. (B) Frequency of organisms and litter (x-axis) with respect to the depth range (y-axis); it documents a higher biodiversity between 370 and 390 m water depth; the stronger impact by fishing gears is located between 370 and 410 m, associated with hard substrates, whereas the maximum dumping impact is found between 430 and 460 m on muddy bottoms.

(A) Map of the Mediterranean Sea showing the main path of the LIW and the deep water currents (modified from)53,54; the major cold-water coral Provinces are reported27, upgraded to including the new Dohrn Canyon site: 1. South Sardinia, 2. South Malta, 3. Santa Maria di Leuca, 4. Bari Canyon, 5–6. Gulf of Lions, 7. Melilla, 8. Dohrn Canyon. (Map modified from)24. (B) T-S plot of 8 different depth profiles sourced from the World Ocean Atlas 2013 (WOA13) database55,56 in the area of the Gulf of Naples (area between 40°52′30″N and 40°22′30″N; 13°52′30″E and 14°52′30″E). Isopycnals are calculated at 0 m (σ0). MAW: Modified Atlantic Water, LIW: Levantine Intermediate Water, TDW: Tyrrhenian Deep Water. The plot was created using Ocean Data View (Schlitzer, R., Ocean Data View, http://odw.awi.de, 2015).
Nekton diversity during the ROV survey revealed the presence of a limited number of species. Shoals of Pagellus bogaraveo were observed swimming next to the canyon wall, and the silver scabbardfish Lepidopus caudatus was abundant near the wall and the bottom of the canyon. ROV explorations were extended also on the adjacent muddy bottoms of the canyon. Here we observed the presence of sparse cerianthiids, unidentified Paguridae, other benthopelagic fishes and sponges (Fig. 4C). In particular, the Ceriantharia Pachycerianthus cf. dohrni reached here densities up to 5–6 ind. m−2 (Fig. 4D). Finally, the ROV images documented a high zooplankton biomass, dominated by Euphausiacea and Chaetognata.
Dating the Canyon occupancy by the coral-bivalve biotope
We have observed the common presence of subfossil oysters cemented to the wall (Fig. 4A,B). Some dead D. dianthus and Caryophyllia calveri appear also completely patinated by ferromanganese oxides, serving as attachment bases for living individuals of D. dianthus and J. cailleti, respectively (Fig. 7B,C). Noticeably, incipient oxide patination was observed on some live D. dianthus corals (Fig. 7D), although such polymetallic patina often testifies at a prolonged post-mortem exposure of the calcareous skeleton within the water column29,30. One dead valve of N. zibrowii coated by a thick black ferromanganese patina (Fig. 7A) was collected from the wall and radiocarbon dated at the Poznan Radiocarbon Laboratory in Poland, providing a calibrated age of 5496 yr cal BP (2σ range: 5329–5588) (Table 3). We hypothesize that this age might represent the time of coral-bivalve settlement of the Dohrn Canyon. The date matches well with the timing of re-colonization that followed the collapse of CWC-growth in the eastern Mediterranean Sea due to oxygen deficiency in intermediate and deep waters between 10.8–6.1 kyr BP31.

Samples taken by the ROV robotic arm at st. DOHRN_ROV_1: (A) dead and partly broken shell of Neopycnodonte zibrowii that provided a calibrated age of 5496 yr cal BP; note the black oxide patina which also affects numerous fouling organisms as solitary corals and serpulid polychaetes (−373 m); (B) a juvenile live-collected Javania cailleti on the side of a dead and patinated Caryophyllia calveri coral (−372 m); (C) juvenile live-collected Desmophyllum dianthus growing on the calyx of an older specimen (−371 m); (D) this Desmophyllium dianthus was collected alive, but its skeleton was already stained by an incipient oxide patina (−377 m). Scale Bars = 1 cm.
Thus, to the best of our knowledge, the Dohrn Canyon became home of this peculiar biotope since the mid Holocene at least, along with the onset of the LIW at the end of the sapropel S1 event at 6.1 kyr BP. We have no indication of a previous presence of CWC in this site, although it cannot be excluded a-priori given the Late Pleistocene L. pertusa recorded from off the Gulf of Naples32.
Anthropogenic impact
The ROV survey provided evidence of severe anthropogenic impacts on the Dohrn Canyon (Figs 5 and 8). Dohrn_01, MS17-81 and MS17_102 ROV tracks on the canyon’s bottom documented the abundant presence of marine litter even of large size (Fig. 8A,C), as well as lost nets and longlines seen enveloping also epifaunal megabenthos. A high number of longlines was reported entangled on substrate asperities (Fig. 8B,D) often close to colony of cold-water corals and bivalves. Longlines served also as attachment substrate for cnidarians, including M. oculata and D. dianthus (Fig. 8B,D). Our data pointed out the presence on an impressive amount of illegal dumping of garbage bags, so intense to completely drape the sea-bottom at places (Fig. 8E). Marine litter has been classified following the MSFD guidance provided by33, with density indicated as items/100 m based on the total length of the ROV tracks (Fig. 5B). Marine litter attains an overall density of 5.03 items/100 m, considering a total of 136 different objects (93 plastic objects: 3.44 items/100 m; 43 lost fishing gears: 1.59 items/100 m). Plastic objects are computed only for muddy bottoms (4.91 items/100 m), whereas lost fishing gears (5.32 items/100 m) are evident on hard substrates (Figs 5 and 8B,D; Table 4).

Typology of macroscopic anthropogenic impacts in the Dohrn Canyon including litter and lost fishing gears: (A) dumped mattress and plastic bag (st. ROV_DOHRN_1, −430 m); (B) colony of Madrepora oculata grown on lost longline (st. ROV_DOHRN_1, −375 m); (C) accumulation of dumped bags and other plastic objects completely covering the bottom (st. MS17-102, −453 m); (D) ghost net on a relatively bare substrate, note the solitary coral Desmophyllum dianthus (arrow) sitting on the same net (st. MS17_102, −453 m).
Discussion
Despite the fact that intensive biological investigations have been conducted in the GoN since the foundation of the Stazione Zoologica in 1872, information on deep-sea benthos has remained surprisingly very scant and limited to mobile bottoms34,35,36. The hard bottoms of the Dohrn Canyon reveal further elements of seafloor heterogeneity in the Gulf, which increase the incredibly rich topographic features of the coastal habitats, with all predictable ecological consequences37. It is surprising, however, that the presence of such a highly diverse habitat was so close to the shoreline and remained unnoticed until present. This finding further strengthens the evidence of the still insufficient knowledge of the deep-sea realm even in proximity of the coast38,39, and of the deep-sea Mediterranean biodiversity in particular40,41.
With respect to the cold-water coral habitats of the Mediterranean basin, the Dohrn Canyon is of particular oceanographic and ecological interest as it represents an element of potential connectivity with other deep-water coral populations of the basin. The GoN indeed is peripheral, but still on the track of the LIW (Fig. 6A), which is hypothesized to control the distribution of major deep-water coral grounds in the Mediterranean basin42.
The deep-sea assemblages revealed by this study indicate that the Dohrn Canyon is a highly diverse cold-water coral site of the eastern Tyrrhenian Sea, for which available information previously confirmed the occurrence of living specimens of M. oculata, L. pertusa and D. dianthus (pers. unpublished data), but never the contextual presence of N. zibrowii and A. excavata except in the case of subfossil Pleistocene records30,43.
A unique trait of the faunal assemblage of the Dohrn Canyon, however, is the tight co-existence of living specimens of the bivalves A. excavata and N. zibrowii with the cold-water corals, which has never been previously reported in the Mediterranean Basin. The assemblages of the Dohrn Canyon share several common traits with those inhabiting the Whittard Canyon of the eastern Atlantic Ocean44,45,46. Both canyons share, indeed, the presence of the same large bivalves of A. excavata and N. zibrowii, although with a much higher bivalve density in the Whittard than in the Dohrn Canyon. However, in both areas N. zibrowii appears to preferentially settle at slightly deeper depths than A. excavata. Conversely to what reported from the Whittard Canyon, solitary corals predominate in the Dohrn Canyon.
The new finding reported here from the Dohrn Canyon clearly indicates that the GoN is a hot spot of deep-sea benthic diversity coexisting with high levels of anthropogenic impact. The canyon head, indeed, is very close to the Naples megalopolis, and is subjected to a severe dumping and ghost- fishing impact, that were clearly documented by ROV images. Data on the number of large litter, and plastic debris, and of fishing nets and lines in the deep are still very scant47,48, so that remains difficult to provide sound comparisons. However, we did not find evidence of trawling and topographic elements of physical damage and habitat destruction (e.g., dredging). Data reported in the present study, thus suggest that hot spot of deep-sea benthic biodiversity can coexist with these impacts (litter, long lines) if the latter are not coupled with the habitat destruction. We call for the preservation of these unique deep-sea habitats by adopting protection measures from bottom contact fishing and other sources of impact. This goal could be partially achieved by granting to the Dohrn Canyon the status of Site of Community Importance (SCI) within the European Commission Habitats Directive, and by starting action to establish a deep-sea Marine Protected Area in this Mediterranean area.
Methods
A new swath bathymetry on a sector of Dohrn Canyon was acquired during the second cruise using Kongsberg Simrad 8160 Multibeam Echosounder hull-mounted with a frequency of 44 kHz. All data were plotted in the Transverse Mercator – UTM33N-WGS84 Coordinate System. Morphobathymetric maps were elaborated, with a cell size of 5 × 5 m, using CARIS HIPS and SIPS and Teledyne RESON PDS2000 software. The survey area extends from 140 to 634 m depth and covered a total area of 104 km2.
All four ROV surveys have been conducted using a Pollux III ROV, equipped with low definition CCD video-camera for navigation and general description, and high definition (2034 × 1296 pixels) video-camera (SonyHDR-HC7) for detailed description. Three laser beams at distance of 20 cm each other, provided the scale bar on the videos. The ROV was equipped with an acoustic position system which provided the exact geographical and depth position every 1 s. Videos have been analyzed with Adelie Video and Adelie GIS based on ArcMap® Geographic Information System (distributed by ESRI). The tool “minifilms” was used to extract high-resolution still images from ROV footage every 10 seconds: 2048 photographs have been obtained with each image covering ca. 6 m2. A total length of 2703.36 m (more than 15,000 m2) was surveyed during the four dives, of which 1895.58 m pertain to mobile substrates (such as mud) and 807.77 to hard substrates (e.g., rocky outcrops).
Taxa recognition was based on still HD images analysis and on a limited number of samples obtained by the ROV’s robotic arm. Sampled organisms were first photographed, and then fixed on board in a buffered 4% formaldehyde solution or preserved in proof ethanol for genetic analyses. Taxonomic identification was conducted in the laboratory using the proper techniques of each taxonomic group, and conforms to the World Register of Marine Species49.
A first ROV exploration of the Dohrn Canyon habitat was conducted in June 2016 during the oceanographic cruise ANOMCITY16 onboard R/V Minerva Uno (station code DOHRN_ROV_1), followed by others explorations in July 2016 and July 2017 for the initial assessment and monitoring of the new site within the frame of the Italian Marine Strategy Framework Directive (station codes MS16_ and MS17_, respectively). In total, four ROV dives explored three distinct sectors of the canyon head’s NW branch, along transects perpendicular to the axis of the channel and ending on the steep rocky margin of the channel. Dives DOHRN_01 and MS16_7 have been the first two exploratory dives in the canyon revealing high biodiversity level (Fig. 1B,C, and Table 2). Both dives explored the northwestern flank of the canyon at depth comprised between 375 and 455 m. Deeper parts, from 420 to 455 m are characterized by gentle topography and muddy substrate with no evident mega- and macrofauna. The two dives explored in detail the canyon wall from 375 to 410 m where rocky substrate outcrop. To the west of this sector (MS17_81), the area is characterized by a relatively gentle morphology between 360–450 m water depth, with muddy sediment prevailing over scattered outcropping bedrock occurrences at 380 and 450 m respectively. The easternmost ROV survey (MS17_102) explored the canyon’s head between 345–470 m. This side of the canyon is characterized by gentle muddy topographies alternating with rocky outcrops and cliffs (vertical walls) between 440 and 460 m and 390 and 430 m respectively.
A fragment (~20 mg) of a fossil shell of N. zibrowii was carefully cleaned using a small diamond blade to remove any visible contamination, leached with diluted HCl and H3PO4 and analyzed for AMS-14C at the Poznan Radiocarbon Laboratory in Poland. The radiocarbon age was converted into calendar years (cal. yr BP, BP = AD 1950) using the Marine1350 calibration curve and the Calib7.0.4 program51. Prior to calibration, the age was corrected for an extra 49 ± 42 years regional reservoir effect (ΔR) using the value reported in the Marine Reservoir Correction Database (http://calib.qub.ac.uk/marine/).
References
Pacione, M. Socio-spatial development of the South Italian City: The case of Naples. T. I. Brit. Geogr. 12, 433–450 (1987).
Mendia, L. et al. Pollution control and quality levels of the Bay of Naples Mediterranean Coastal Pollution (ed. Jenkins, S. H.), Prog Wat Tech 12, 615–631, Pergamon Press Ltd (1980).
Oppenheimer, C. H., Oppenheimer, D. P. & Blundo, R. An ecological survey of the Gulf of Naples area, conducted during September 15 to October 15, 1976, for the Region of Campania, in Marine Environmental Pollution, 2 Dumping and Mining (ed. Geyer, R. A.) 67–142 (Elsevier Scientific Publishing Company, Amsterdam, 1981).
Volterra, L. et al. Microbiological pollution of marine sediments in the southern stretch of the Gulf of Naples. Water Air Soil Pollut. 26, 175–184 (1985).
De Pippo, T. et al. Seabed morphology and pollution along the Bagnoli coast (Naples, Italy): a hypothesis for environmental restoration. PSZN Mar. Ecol. 23(Suppl 1), 154–168 (2002).
Naso, B. et al. Persistent organic pollutants in edible marine species from the Gulf of Naples, Southern Italy. Sci. Tot. Environ. 343, 83–95 (2005).
Zingone, A. et al. Potentially toxic and harmful microalgae from coastal waters of the Campania region (Tyrrhenian Sea, Mediterranean Sea). Harmful Algae 5, 321–337 (2006).
Uttieri, M. et al. Multiplatform observation of the surface circulation in the Gulf of Naples (Southern Tyrrhenian Sea). Ocean Dynamics 61, 779–796 (2011).
Quaranta, F. et al. The environmental impacts of cruise ships in the Port of Naples: Analysis of the pollution level and possible solutions. J. Mar. Res. 9, 81–86 (2012).
Wang, M. et al. Investigation on inorganic pollution level in surface sediments of Naples and Salerno Bay. Computational Water, Energy, and Environmental Engineering 2, 36–40 (2013).
Tornero, V. & Ribera d’Alcalà, M. Contamination by hazardous substances in the Gulf of Naples and nearby coastal areas: A review of sources, environmental levels and potential impacts in the MSFD perspective. Sci. Tot. Environ. 466–467, 820–840 (2014).
Angiolillo, M. et al. Distribution and assessment of marine debris in the deep Tyrrhenian Sea (NW Mediterranean Sea, Italy). Mar. Poll. Bull. 92, 149–159 (2015).
Scarpato, A. et al. Western Mediterranean coastal waters - Monitoring PCBs and pesticides accumulation in Mytilus galloprovincialis by active mussel watching: the Mytilos project. J. Environ. Monit. 12, 924–935 (2010).
Milia, A. The Dohrn canyon: a response to the eustatic fall and tectonic uplift of the outer shelf along the eastern Tyrrhenian Sea margin, Italy. Geo-Mar. Lett. 20, 101–108 (2000).
D’Argenio, B. et al. Digital elevation model of the Naples Bay and adjacent areas, Eastern Tyrrhenian sea. Atlante di Cartografia Geologica scala 1:50,000 (progetto CARG), Servizio Geologico d’Italia (APAT). 32° International Congress “Firenze 2004”, Editore De Agostini, Italy (2004).
Di Fiore, V. et al. Gravity instabilities in the Dohrn Canyon (Bay of Naples, Southern Tyrrhenian Sea): potential wave and run-up (tsunami) reconstruction from a fossil submarine landslide. Geologica Carpatica 62, 55–63 (2011).
Carrada, G. C. et al. Variability in the hydrographic and biological features of the Gulf of Naples. PSZN Mar. Ecol. 1, 105–120 (1980).
De Maio, A. et al. Dinamica delle acque del Golfo di Napoli. Parte Prima. Annali Ist. Univ. Nav. 47–48, 201–213 (1978–1979a).
De Maio, A. et al. Dinamica delle acque del Golfo di Napoli. Parte Seconda. Annali Ist. Univ. Nav. 47–48, 215–231 (1978–1979b).
Hopkins, T. S. et al. Atti AIOL 10, 375–387 (1994).
Freiwald, A. et al. Cold-water coral reefs. Cambridge: UNEP-WCMC Biodiversity Series 22, 1–88 (2004).
Taviani, M. et al. Deep coral growth in the Mediterranean Sea: An overview. in Cold-water corals and ecosystems (eds Freiwald, A. & Roberts, J. M.), 137–156 Springer Berlin Heidelberg (2005).
Montagna, P. et al. Phosphorus in cold-water corals as a proxy for seawater nutrient chemistry. Science 312, 1788–1791 (2006).
Zibrowius, H. & Taviani, M. Remarkable sessile fauna associated with deep coral and other calcareous substrates in the Strait of Sicily, Mediterranean Sea in Cold-water corals and ecosystems (eds Freiwald, A. & Roberts, J. M.), 807–819, Springer Berlin Heidelberg (2005).
López Correa, M. et al. Distribution and habitats of Acesta excavata (Bivalvia: Limidae) with new data on its shell ultrastructure in Cold-water corals and ecosystems (eds Freiwald, A. & Roberts, J. M.) 173–205, Springer Berlin Heidelberg (2005).
Beuck, L. et al. Biotope characterisation and compiled geographical distribution of the deep-water oyster Neopycnodonte zibrowii in the Atlantic Ocean and Mediterranean Sea. Rapp. Comm. int. Mer Médit. 41, 462 (2016).
Taviani, M. et al. The “Sardinian Cold-water Coral Province” in the context of Mediterranean coral ecosystems. Deep-Sea Res. Pt. II 145, 61–78 (2017).
Wisshak, M. et al. Shell architecture, element composition, and stable isotope signature of the giant deep-sea oyster Neopycnodonte zibrowii sp. n. from the NE Atlantic. Deep-Sea Res. Pt. I 56, 374–407 (2009).
Allouc, J. et al. Phénomènes biosédimentaires et genèse des croutes et enduits polymétalliques. L’exemple des dépots hydrogenètiques de la marge ouest africaine et de la Méditerranée. Geobios 32, 769–790 (1999).
Taviani, M. et al. Pleistocene to Recent scleractinian deep-water corals and coral facies in the Eastern Mediterranean. Facies 57, 579–603 (2011).
Fink, H. G. et al. Oxygen control on Holocene cold-water coral development in the eastern Mediterranean Sea. Deep-Sea Res. Pt. I 62, 89–96 (2012).
McCulloch, M. et al. Proliferation and demise of deep-sea corals in the Mediterranean during the Younger Dryas. Earth Planet. Sc. Lett. 298, 143–152 (2010).
Galgani, F. et al. Guidance on Monitoring of Marine Litter in European Seas. Luxembourg: Publications Office of the European Union, 128 pp., https://doi.org/10.2788/99475 (2013).
Lo Bianco, S. Le pesche abissali eseguite da F.A. Krupp col Yacht Puritan nelle adiacenze di Capri ed in altre località del Mediterraneo. Mittheilungen aus der Zoologischen Station zu Neapel zugleich ein repertorium für mittelmeerkunde 16, Berlin (1903–1904).
Bonaduce, G. & Pugliese, N. Benthic ostracods as depth indicators. Rapp. Comm. int. Mer Médit. 25-26, 167–169 (1979).
Sgarrella, F. & Moncharmont-Zei, M. Benthic Foraminifera of the Gulf of Naples (Italy): systematics and autoecology. Boll. Soc. Paleontol. It. 32, 145–264 (1993).
Zeppilli, D., Pusceddu, A., Trincardi, F. & Danovaro, R. Seafloor heterogeneity influences the biodiversity–ecosystem functioning relationships in the deep sea. Sci. Rep. 6, 26352 (2016).
Ramirez-Llodra, E. et al. Deep, diverse and definitely different: unique attributes of the world’s largest ecosystem. Biogeosciences 7, 2851–2899 (2010).
Danovaro, R., Snelgrove, P. V. R. & Tyler, P. Challenging the paradigms of deep-sea ecology. Trends Ecol. Evol. 29, 465–475 (2014).
Danovaro, R. et al. Deep-sea biodiversity in the Mediterranean Sea: The known, the unknown, and the unknowable. PloS One 5, e11832 (2010).
Coll, M. et al. The biodiversity of the Mediterranean Sea: estimates, patterns, and threats. PloS One 5, e11842 (2010).
Taviani, M. et al. Reprint of ‘On and off the beaten track: Megafaunal sessile life and Adriatic cascading processes’. Mar. Geol. 375, 146–160 (2016).
Remia, A. & Taviani, M. Shallow-buried Pleistocene Madrepora-dominated coral mounds on a muddy continental slope, Tuscan Archipelago, NE Tyrrhenian Sea. Facies 50, 419–425 (2005).
Johnson, M. P. et al. A vertical wall dominated by Acesta excavata and Neopycnodonte zibrowii, Part of an undersampled group of deep-sea habitats. PLoS ONE 8, e79917 (2013).
Amaro, T. et al. The Whittard Canyon–A case study of submarine canyon processes. Progr. Oceanogr. 146, 38–57 (2016).
Daly, E. et al. Bottom trawling at Whittard Canyon: Evidence for seabed modification, trawl plumes and food source heterogeneity. Progr. Ocean . 169, 227-240 (2018)
Galgani, F., Souplet, A. & Cadiou, Y. Accumulation of debris on the deep sea floor off the French Mediterranean coast. Mar. Ecol. Prog. Ser. 142, 225–234 (1996).
Miyake H., Haruka S. & Furushima Y. Deep-Sea Litter Study Using Deep-Sea Observation Tools. Interdisciplinary Studies on Environmental Chemistry—Marine Environmental Modeling & Analysis, Eds., K. Omori, X. Guo, N. Yoshie, N. Fujii, I. C. Handoh, A. Isobe and S. Tanabe, pp. 261–269 (2010).
WoRMS Editorial Board. World Register of Marine Species. Available from, http://www.marinespecies.org at VLIZ. Accessed 2016-11-29, https://doi.org/10.14284/170 (2016).
Reimer, P. J. et al. IntCal13 and Marine13 Radiocarbon Age Calibration Curves 0-50,000 Years cal BP. Radiocarbon 55, 1869–1887 (2013).
Stuiver, M., Reimer, P. J. & Reimer, R. W. CALIB 7.1 [WWW program] at, http://calib.org, accessed2018-5-9 (2018).
EMODnet Bathymetry Consortium: EMODnet Digital Bathymetry (DTM), https://doi.org/10.12770/c7b53704-999d-4721-b1a3-04ec60c87238 (2016)
Pinardi, N. & Masetti, E. Variability of the large scale general circulation of the Mediterranean Sea from observations and modeling: a review. Palaeogeogr. Palaeoclimat. Palaeoecol. 158, 153–174 (2000).
Schroeder, K. et al. Circulation of the Mediterranean Sea and its variability in The climate of the Mediterranean region: from the past to the future (ed. Lionello, P.), 187–256, Elsevier (2012).
Locarnini, R. A. et al. World Ocean Atlas 2013 Volume 1: Temperature (ed. Levitus S.) 1–40, (ed., A. Mishonov Technical Ed.; NOAA Atlas NESDIS 73) (2013).
Zweng, M. M. et al. World Ocean Atlas 2013, Volume 2: Salinity(ed. Levitus, S.) 1–39, (ed., A. Mishonov Technical Ed.; NOAA Atlas NESDIS 74) (2013).
Acknowledgements
Captain, crew and shipboard staff of RV Minerva Uno are acknowledged for their efficient and skilful cooperation at sea during ANOMCITY2016 (Chief Scientist Elvira Oliveri), Strategia Marina Ligure/Tirreno (Chief Scientist Lorenzo Angeletti) and MSFD-IT-2-2017 (Chief Scientist Alessandro Remia) oceanographic cruises. Mariacristina Prampolini and Valentina Grande are especially acknowledged for data acquisition and elaboration of the morphobathymetric maps. We thank François Galgani for advice on marine litter typology. We thank two anonymous reviewers for positive comments that helped improving the clarity of the text. Partial funding was provided by CNR, the Italian Flag Project Ritmare and the Italian Marine Strategy Framework Directive Programmes. RD received funding from the EU Horizon 2020 project MERCES (Marine Ecosystem Restoration in Changing European Seas; grant agreement No 689518). This is ISMAR-CNR Bologna Scientific contribution n. 1909.
Author information
Authors and Affiliations
Contributions
M.T. and R.D. conceived the project and the idea; L.A., F.C. acquired the data at sea; L.A., F.C., P.M. and M.T. analyzed the data; M.T. wrote the manuscript with the assistance of L.A., F.C., R.D. and P.M. All of the authors contribute to the discussion and preparation of the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Taviani, M., Angeletti, L., Cardone, F. et al. A unique and threatened deep water coral-bivalve biotope new to the Mediterranean Sea offshore the Naples megalopolis. Sci Rep 9, 3411 (2019). https://doi.org/10.1038/s41598-019-39655-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-39655-8
This article is cited by
-
The paleo-community of the Sciacca red coral
Facies (2023)
-
Deep-sea fauna at a former red-mud disposal site (Cassidaigne Canyon, NW Mediterranean)
Marine Biodiversity (2023)
-
New morpho-bathymetric data on marine hazard in the offshore of Gulf of Naples (Southern Italy)
Natural Hazards (2022)
-
Massive bioconstructions built by Neopycnodonte cochlear (Mollusca, Bivalvia) in a mesophotic environment in the central Mediterranean Sea
Scientific Reports (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
