
Abstract
Serogroup B invasive meningococcal disease (IMD) is increasing in China, but little is known about the causative meningococci. Here, IMD and carriage isolates in Shanghai characterised and the applicability of different vaccines assessed. Seven IMD epidemic periods have been observed in Shanghai since 1950, with 460 isolates collected including 169 from IMD and 291 from carriage. Analyses were divided according to the period of meningococcal polysaccharide vaccine (MPV) introduction: (i) pre-MPV-A, 1965–1980; (ii) post-MPV-A, 1981–2008; and (iii) post-MPV-A + C, 2009–2016. Over this period, IMD incidence decreased from 55.4/100,000 to 0.71 then to 0.02, corresponding to successive changes in meningococcal type from serogroup A ST-5 complex (MenA:cc5) to MenC:cc4821, and finally MenB:cc4821. MenB IMD became predominant (63.2%) in the post-MPV-A + C period, and 50% of cases were caused by cc4821, with the highest incidence in infants (0.45/100,000) and a case-fatality rate of 9.5%. IMD was positively correlated with population carriage rates. Using the Bexsero Antigen Sequence Type (BAST) system, fewer than 25% of MenB isolates in the post-MPV-A + C period contained exact or predicted cross reactive matches to the vaccines Bexsero, Trumenba, or an outer membrane vesicle (OMV)-based vaccine, NonaMen. A unique IMD epidemiology was seen in China, changing periodically from epidemic to hyperepidemic and low-level endemic disease. At the time of writing, MenB IMD dominated IMD in Shanghai, with isolates potentially beyond coverage with licenced OMV- and protein-based MenB vaccines.
Similar content being viewed by others


Introduction
The encapsulated bacterium Neisseria meningitidis is a leading cause of bacterial meningitis and septicaemia globally, causing over 1.2 million invasive meningococcal disease (IMD) cases annually1. Over 90% of IMD cases are caused by serogroups A, B, C, W, X, and Y2, which are defined by immunological variants of the capsular polysaccharide. Of these, five (serogroups A, C, W, X, and Y) are preventable by protein-conjugate polysaccharide vaccines3. Most IMD is caused by meningococci belonging to one of a limited number of hyperinvasive lineages, recognised as particular clonal complexes (ccs) by multilocus sequence typing (MLST), which are usually associated with a particular serogroup4.
In China, during the 1950s to 1980s, serogroup A (MenA) isolates were responsible for over 95% of cases5, with incidence peaking in 1967 (403/100,000)6. These were predominantly due to ST-5 clonal complex (cc5) and cc17, and in response, a non-conjugate MenA meningococcal polysaccharide vaccine (MPV) was routinely administered from 1980 onwards6,7. This was followed by a decrease in MenA incidence. From 2003–2005, serogroup C hypervirulent lineage ST-4821 complex (MenC:cc4821) caused outbreaks in Anhui province5, leading to the predominance of MenC IMD caused by MenC:cc48216,8. As a result, in 2008, a serogroup A and C bivalent MPV was introduced into the vaccination program6, followed by an overall IMD incidence decrease to 0.047/100,000, although this may have been underestimated7. From 2011 onwards, the proportion of MenC IMD began to decrease while MenB increased from 7.2% in 2006 to 26.5% in 2014 nationwide7,9, with a few regional MenW:cc11 cases10.
Prevention of MenB IMD is challenging due to the poorly immunogenic serogroup B capsular polysaccharide and concerns about autoimmunity, due to its structural similarity to structures occurring on human tissue. To address this deficit, two protein-based vaccines, Bexsero (4CMenB) and Trumenba (bivalent rLP2086), have been developed and licensed in Europe and the USA11,12. Bexsero is composed of factor H binding protein (fHbp), Neisserial heparin-binding antigen (NHBA), Neisseria adhesin A (NadA), and outer membrane vesicles (OMVs) containing the porin PorA (meningococcal subtyping antigen) as a major immunogen, whereas Trumenba contains two fHbp-subfamily variants13. Both Bexsero and Trumenba may elicit protective responses across meningococci of diverse serogroup expressing these or related protein variants14. Two methods have been established to predict Bexsero coverage. (i) The Meningococcal Antigen Typing System (MATS) combines phenotypic and functional assays12; however, it is time and labour intensive, requires toddler serum, and is only performed by specialist laboratories. (ii) The Bexsero Antigen Sequence Typing (BAST) which uses sequence data is a rapid, scalable, and portable genotypic approach, which catalogues deduced peptide sequences and matches to vaccine variants (BAST-1) or cross-reactive variants15. The meningococcal antigen surface expression (MEASURE) system is used to assess Trumenba coverage by flow cytometry16.
Limited information is available documenting N. meningitidis isolates associated with IMD and carriage in China over the past 60 years. In this study, fluctuations of IMD and meningococcal carriage are described in association with the introduction of MPV vaccines in Shanghai, China since 1950. In addition, and in response to increasing MenB IMD7, we assessed the potential impact of protein-based vaccines to local prevalent serogroups and clonal complexes.
Results
Epidemiology and characterisation of N. meningitidis isolates associated with IMD and carriage in Shanghai
From 1950 to 2016, seven IMD epidemic periods were observed, each lasting 8–10 years (Fig. 1). Average incidence per 100,000 population was: 1953–1961, 21 (case-fatality rate, 8.5%); 1962–1972, 87 (3.3%); 1973–1981, 2.9 (5.7%); 1982–1990, 1.6 (6.3%); 1991–2000, 0.23 (3.7%); 2001–2008, 0.17 (9.8%); and 2009–2016, 0.02 (15%). Highest incidence occurred in children aged <5 years, decreasing with age, except in those aged 15–19 years from 2005 (Fig. 2). Seasonality of IMD rates was apparent: 50–70% of cases occurred between February and April, with fewer cases (8–23%) between June and October (Fig. 3). A positive correlation was observed between observed carriage rate and IMD incidence (Fig. 4).

Invasive meningococcal disease incidence in Shanghai, China during 1950–2016, as reported in National Notifiable Diseases Registry System. The times of introduction of serogroup A (1980) and serogroups A and C polysaccharide vaccines (2008) in Shanghai, China were indicated with red arrow. Inset figure shows the incidence after 1970. The highest incidences in different epidemic period were labelled. MenA, serogroup A meningococcus; MPV, meningococcal polysaccharide vaccine.

Analysis of invasive meningococcal disease incidence in Shanghai, China by age group.

Seasonality of invasive meningococcal disease in Shanghai, China.

Relationship between invasive meningococcal disease incidence and meningococcal carriage rate observed in Shanghai, China.
Based on the time of introduction of MPVs in China (1980 and 2008), three periods were defined: (i) pre-MPV-A, 1965–1980; (ii) post-MPV-A, 1981–2008; and (iii) post-MPV-A + C, 2009–2016 (Table 1 and Fig. 5).

Minimum-spanning tree analysis of multilocus sequence types of invasive and carriage N. meningitidis before and after introduction of meningococcal vaccines in China. Isolates were obtained during the pre-MPV-A (1965–1980), post-MPV-A (1981–2008), and post-MPV-A + C (2009–2016) periods. Sequence types (STs) are displayed as circles. The size of each circle indicates the number of isolates with this particular type. Serogroup is distinguished by different colours. The shaded halo surrounding the STs encompasses related sequence types that belong to the same clonal complex. Heavy solid lines represent single-locus variants, and light solid lines represent double-locus variants. Sequences types sharing no less than 4 loci, but not assigned to any clonal complexes in the PubMLST database were assigned to ST-clusters. NG, nongroupable.
(i) In the pre-MPV-A period, the average IMD incidence was 55.4/100,000. MenA isolates were predominant (84/117, 71.8%; Table 1), with most belonging to cc5 (48/84, 57.1%) and cc1 (36/84, 42.9%). Among MenA:cc5 isolates, ST-5 prevailed (37/48, 77.1%), with no ST-7 observed. All ST-5 contained PorA P1.20,9, while of the MenA:cc1 isolates, 34/36 (94.4%) were ST-3, with 32/34 (94.1%) P1.7-1,10. MenB isolates were assigned to: cc41/44 (6/20, 30%); cc32 (3/20, 15%); cc8 (1/20, 5%); cc35 (1/20, 5%); and cc198 (1/20, 5%), with eight STs unassigned to a cc. MenC isolates were assigned to ST-9514 cluster (44.4%, 4/9; Fig. 5 and S1), cc4821 (3/9, 33.3%), and cc231 (1/9, 11.1%). MenC:cc4821 isolates were all ST-3436 with P1.20-3,23 several variants including P1.20-3,23-1 and P1.20-3,23-3. The observed meningococcal carriage rate ranged from 24.1% in 1967 to 2.4% in 1972 (Table S1), with overall carriage rates of 4.4% (368/8,319) in children and 9.9% (888/8,956) in adults (≥15 years). In 1966–1967, high IMD incidence (>200/100,000) coincided with high carriage rates (>15%), of which a high proportion (>70%) was MenA. This decreased from 50% in 1970 to 1.1% in 1979. Among the 178 carriage isolates analysed, MenB (52.2%) was predominant (Table 1), with cc32 (17/93, 18.3%) the most prevalent.
(ii) In the post-MPV-A period, the average IMD incidence was 0.71/100,000. Based on 61 IMD cases with available serogroup data, MenC 28/61 (45.9%) was the most frequent, in which isolates belonging to cc4821 (17/19, 89.5%) dominated, with the majority of these ST-4821 (15/17, 88.2%) and P1.7-2,14. MenA:cc5 (10/16, 62.5%) dominated the MenA isolates: of the eight collected during 2005–2008 6/8 (75%) were ST-7 and P1.20,9, and two isolated in 1985 were ST-5, P1.20,9. MenB isolates were assigned to cc4821 1/7 (14.3%) (ST-5798:P1.10,13-1), cc41/44 (1/7, 14.3%), and cc32 (1/7, 14.3%), with four isolates unassigned to a cc. The carriage rate from the 2007 survey was 11/553 (2.0%), with 9/369 (2.4%) in children and 2/184 (1.1%) in adults (15–46 years). MenB (16/24, 66.7%) was predominant in carriage (Tables 1), 31.3% of which belonged to cc4821, with 5 different STs each possessing a different PorA VR type.
(iii) In the post-MPV-A + C period, the average IMD incidence was 0.02/100,000. MenB (63.2%) isolates predominated, 50% of which were cc4821 and assigned to 5 STs each with a different PorA VR type (Fig. 5). All 7 MenC isolates were assigned to cc4821. Except one DNA sample with incomplete ST, other 6 MenC:cc4821 isolates were ST-4821, with 5 exhibiting PorA P1.7-2,14. The carriage rate ranged from 0.5% in 2011 to 1.6% in 2014, with 25/1,660 (1.5%) in children and 73/4,624 (1.6%) in adults (15–78 years). MenB (75/89, 84.3%) was the most frequent serogroup in carriage (Table 1), with 20/75 (26.7%) cc4821.
Features and seasonality of MenB IMD
From 1965 to 2016, 34/47 (72.3%) of all MenB IMD occurred in children, while 60/150 (40%) of non-B IMD cases occurred in this age group (p = 0.01). Since 2005, all MenB IMD cases were in children (19 days to 12 years), among which 13/20 (65%) were infants. During 2005–2008, MenB IMD incidence was 0.01/100,000, highest among infants (1.1/100,000) compared to 0.009/100,000 in children aged 1–15 years, with no reported deaths. During the post-MPV-A + C period, MenB IMD incidence was 0.007/100,000, the highest of which in infants (0.45/100,000) compared to 0.03/100,000 in children aged 1–15 years, with a case-fatality rate of 2/21 (9.5%). During the post-MPV-A + C period, MenB cases were observed from February to September, and in December while all MenB cases from 2005–2008 occurred from January to June.
BAST identification, prevalence of vaccine antigens and potentially cross-reactive variants
A total of 243 BASTs were identified with high diversity in each of the vaccine antigens: fHbp, 64 variants; NHBA, 95; NadA, 9 (the nadA gene was absent or had gene-silencing frameshift mutations in 367/460 or 82.0% of isolates); PorA VR1, 38; and PorA VR2, 64. A total of 56 BASTs were identified in the 169 IMD isolates (0.33 BASTs/isolates). The four most prevalent BASTs were BAST-13 (associated with cc5), BAST-794 (cc1), BAST-802 (cc4821) and BAST-22 (cc5), representing by 102/169 (60.4%) of the meningococcal disease isolates. In the 291 carriage isolates, 201 BASTs were identified (0.69 BASTs/isolates). The four most prevalent BASTs, BAST-2300 (ST-9514 cluster), BAST-13 (cc5), BAST-794 (cc1), and BAST-2262 (ST-5620 cluster), were represented by 40/267 (15.0%) of the isolates. BASTs fluctuated with ccs found in the pre- and post-MPV periods (Table 1).
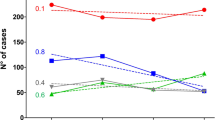
Combined exact matches and putative cross-reactive antigens, revealed that 15/221 (6.8%) of MenB isolates were potentially covered by Bexsero, and among IMD MenB isolates, these constituted: 3/20 (15%) in pre-MPV-A; none in post-MPV-A; and none in post-MPV-A + C periods (Fig. 6). For Trumenba, no exact antigen match was found, and putative cross-reactive variants were 90/221 (40.7%) among MenB isolates. In IMD MenB isolates this constituted: 10/20 (50%); 1/8 (12.5%), and 2/9 (22.2%) in each respective period. For NonaMen, the prevalence of vaccine antigens among MenB isolates was 34/221 (15.4%), and in IMD MenB isolates, 3/20 (15%), 2/8 (25%), and none, respectively.

Prevalence of peptide variants, and potentially immunologically cross-reactive variants, for three serogroup B-substitute vaccines (Bexsero, Trumenba, and NonaMen) among 460 invasive and carriage meningococci from Shanghai, China in the pre-MPV-A, post-MPV-A, and post-MPV-A + C periods. Bexsero and Trumenba are two protein-based serogroup B substitute meningococcal vaccines, which have been licensed in Europe and the USA, while NonaMen is a 9-valent investigational outer membrane vesicle (OMV) vaccine, which has undergone pre-clinical testing. Three periods were defined, pre-MPV-A (1965–1980), post-MPV-A (1981–2008), and post-MPV-A + C (2009–2016), according to the time of two meningococcal polysaccharide vaccines introduced in China (1980 serogroup A, 2008 A and C).
The prevalence of homologous and potentially cross-reactive peptide variants was noted for the 5 most prevalent ccs, which corresponded to 238/460 (51.7%) of the IMD meningococcal isolates, namely: cc1; cc5; cc32; cc41/44; and cc4821. With the exception of cc5 (73/74, 98.6%) the prevalence of such variants for Bexsero was low: cc1, none; cc4821, none; cc32, 1/23 (4.3%); and cc41/44, 9/21 (42.9%). Prevalence of similar antigens for Trumenba was: cc32, 5/23 (21.7%); cc4821, 29/72 (40%); cc41/44, 15/21 (71.4%); cc1 and cc5 < 2%. No NonaMen antigens were observed in isolates belonging to cc1, cc5, cc41/44 or cc4821, while 16/23 (69.6%) of cc32 isolates contained homologous PorA sequences. For cc4821 isolates, Trumenba contained homologous or potentially cross-reactive antigen sequences among IMD cc4821 isolates as follows: none in pre-MPV-A; 1/14 (7.1%) in post-MPV-A; and 2/10 (20%) in post-MPV-A + C periods. Among carriage cc4821 isolates, this was 10/15 (66.7%), 1/7 (14.3%) and 15/23 (65.2%), respectively.
Discussion
This study provides a comparative analysis of IMD and meningococcal carriage isolates obtained in Shanghai, China, and the likely impact of novel vaccines on disease and carriage. From 1965 to 1980, IMD in China was dominated by MenA meningococci belonging to the hyperinvasive lineages corresponding to cc5 (ST-5) and cc1 (ST-3), resulting in several epidemics (Figs 1 and 5), consistent with similar MenA epidemics seen elsewhere17,18. Introduction of serogroup A MPV vaccine in China in 1980 was followed by a decrease in IMD; however, this in turn may have contributed to expansion of MenC:cc48215. Clonal expansion following vaccine implementation was further observed with the implementation of serogroup A and C MPV in 2008 which was followed by an increase in MenB IMD, largely due to MenB:cc4821 isolates (Figs 1 and 5, Table 2)19. These data indicate that vaccine intervention may have promoted the emergence of different IMD-associated meningococci, consistent with the fluctuation of hyperinvasive lineages in the carried population; indeed, similar changes subsequent to vaccine implementation with MPV A + C were observed in Egypt and Morocco during 1992–199520.
Since the 1950s, the seasonal peak of IMD cases in Shanghai has been from February to April (Fig. 3), similar that seen nationally in China6. China is a large country, with differences seen for example in the peak influenza season between northern (January) and southern China (from June to July), with the latter characterised by a warmer, more humid climate21. This suggests that, besides climate and influenza incidence, national social factors should be considered when deploying preventative strategies. IMD outbreaks, such as the MenA:cc5 global pandemic and MenW:cc11 transmission, are often associated with mass gathering events17,22. For example, the 1967 MenA epidemics across China (403/100,000) occurred following the National Great Networking event during 1966–19676,23, which involved gatherings of millions of students from all over the country23. The observed seasonal peaks in IMD incidence (Fig. 3) may be associated with the Spring Festival travel rush for the Chinese New Year where, from January to March, over 200 million people travel across China for family reunions24. Poor sanitary conditions and overcrowded environments on public transport is likely to facilitate transmission of meningococci, indicating the need for enhanced understanding of meningococcal carriage before, during, and after the Chinese New Year.
Few carriage surveys have been undertaken in China; however, two studies in the Shandong and Guangxi provinces identified high carriage rates of meningococci from hyperinvasive ccs in association with IMD outbreaks10,25. This is consistent with results from our study where carriage rates positively correlated with incidence (Fig. 4 and Table 1). In addition, MenB cases were not linked to a distinct seasonal pattern such that in the post-MPV-A + C period, MenB cases were found to occur throughout the year in contrast to that seen in the 1950s (Fig. 3).
Since the 1980s, three monovalent OMV-based MenB vaccines have been licensed for IMD epidemics, but these demonstrated clinical efficacy only against homologous meningococci13. Although a nonavalent OMV-based MenB vaccine has been evaluated26, the vaccine variants were found low prevalence among Chinese MenB IMD meningococcal isolates in this study (<5%) and from 27 provinces of China (<11%)19. Two protein-based MenB substitute vaccines have been licensed and implemented in vaccination interventions in Europe and the USA11,12. The coverage of MenB isolates by Bexsero in the UK during 2014/15 was predicted to be 60.8% using BAST15, and 66% by MATS27. For Trumenba, coverage rates of 78–100% to collections of diverse strains in Europe and the USA was estimated using serum bactericidal assay28,29,30. In this study, the prevalence of meningococcal antigens potentially covered by the variants present in the vaccine was low in Shanghai for both Bexsero (≤15%) and Trumenba (≤50%). Based on fHbp data from 30 provinces across China31, Trumenba was predicted to potentially cover 32.5% of IMD and 40% of MenB carriage isolates. This low prevalence is due to the predominance of cc4821 meningococci in China19, which share no homologous antigens with Bexsero and few with Trumenba (40.3%). In Europe and the USA, MenB IMD has been mainly caused by cc32, cc41/44, and cc269 over the past 20 years32, which exhibit distinct antigenic profiles to those found in China (Table 1). Therefore, the likely impact of Bexsero on Chinese cc32 (4.3%) and cc41/44 (42.9%) was lower than in Europe (93–100%)27. Alternative vaccination approaches include an OMV-based vaccine specific for MenB cc4821, and the characterisation of ST and antigen data, especially PorA variants, reported here will be invaluable in assessing vaccine coverage and future serogroup B-substitute vaccine development in China.
The interpretation of these data is limited by incomplete records of MenB IMD cases during 1950–2004 and the small number of isolates collected during 1965–1980 and 1981–2008; however, data from children’s hospitals in Shanghai and Beijing provide some insight into MenB IMD during 1976–2002. In Shanghai, from 1976 to 1985, 33.3% (20/60) of paediatric IMD cases were caused by MenB, with 45% (9/20) in infants (<1 year) and a fatality rate of 10% (2/20). During 1984–1993, 42% (21/50) of IMD cases were due to MenB, with 71.4% (15/21) occurring in children aged ≤2 years33,34. In Beijing, during 1984–2002, 51% (102/200) of paediatric IMD cases were caused by MenB with 51.0% (52/102) in infants and a fatality rate of 11.8% (12/102)35. Among these MenB cases in Shanghai and Beijing, 59.2% (84/142) occurred outside the “epidemic season” (from February to April). Therefore, it is likely the two features of MenB IMD, namely (i) the occurrence in young children and (ii) lack of seasonal variation in incidence, have persisted since the 1970s.
Our data suggest that vaccine coverage of MenB meningococci in Shanghai by licensed OMV and protein-based ‘serogroup B substitute’ MenB vaccines may be limited. Therefore, a cautious, region-specific approach to implementation of new protein-based meningococcal vaccines should be considered. Further, the temporal analysis suggests that vaccine implementation coinciding with the start of the calendar year, so as to disrupt transmission events during Spring Festival might have the highest impact on IMD incidence. In conclusion, our data indicate that IMD surveillance in China should be enhanced, combined with comprehensive carriage studies to assess the impact of vaccines and their likelihood inducing both effective direct protection and herd immunity.
Methods
IMD surveillance
IMD surveillance in Shanghai, implemented in the National Notifiable Diseases Registry System (NNDRS), began in 1950 and was based on monthly paper reports. Since 2004, it has become a web-based, real-time system6. All clinical specimens and meningococcal isolates from suspected IMD cases in Shanghai are sent to Shanghai CDC when they are reported in the NNDRS6. In China, a child is defined as an individual <15 years of age and an infant <1 year6.
N. meningitidis carriage surveys
Twenty carriage studies were conducted during 1965–2016. In each study, three districts were chosen, including urban, suburban, and rural districts. Posterior oropharyngeal swabs were collected from preschool children (toddlers aged 3-6 years in childcare centres), students (aged 6-14 years in schools), and adults (staff in department stores, railway stations, army, and residents in communities), and cultured as previously described36.
Isolate collection
From 1965–2016, 460 meningococcal isolates were collected in Shanghai, excluding the period 1986–2004 when isolates were not stored. As a result, 169 IMD and 291 carriage isolates dating from 1965–1985 (n = 306) and 2005–2016 (n = 154) were available for study. Serogroup was determined by slide agglutination using monoclonal antiserum (BD, USA) and PCR detection of the appropriate capsule synthesis gene37. Isolate serogroup distribution was: A, 123 isolates; B, 221; C, 62; E, 13; W, 5; X, 3; Y, 9; and Z, 3; and 21 nongroupable (NG, negative by PCR and sera agglutination). Sequence type (ST), cc, porA and fetA variants were determined from nucleotide sequences by querying the PubMLST.org/Neisseria database38. Relationships between STs were analysed using BioNumerics software package (version 7.6.2; Applied Maths, Belgium).
BAST identification and vaccine coverage estimates
BAST was determined as described previously15. Briefly, genome sequences were used to deduce peptide sequences by in silico translation of the nucleotide sequences of fhbp, nhba, nadA, and porA variable regions 1 and 2 (VR1 and VR2). Variant numbers were assigned to unique peptide sequences using the established nomenclatures39,40,41. Each unique combination of the 5 antigenic variants was assigned a BAST number, in order of discovery; BAST-1 refers to the Bexsero vaccine constituents: fHbp 1, NHBA 2, NadA 8, and PorA P1.7–2,4. Exact matches and potential cross-reactive matches were combined to evaluate coverage of Bexsero, Trumenba, and NonaMen, a 9-valent OMV-based vaccine (Table S2)26,28,29,30,42,43.
Statistical analysis
Statistical analysis was performed using SPSS (version 20.0; IBM, USA). Fisher’s exact test was used to compare proportions of IMD occurring in children with causative serogroup. Statistical significance was assessed at p < 0.05. The correlation coefficient between carriage rate and IMD incidence was calculated using Microsoft Excel 2010.
Ethical Aspects
All the specimens from meningococcal patients and carriers were collected as part of the routine clinical management of invasive meningococcal cases, according to the national guidelines in China. Informed consent has been obtained, including that from a parent and/or legal guardian of the participants under the age of 18 years. All these experimental protocols were approved by the Institutional Review Board of Huashan Hospital, Fudan University.
Data Availability
All data generated or analysed during this study are included in this published article and its Supplementary Information files.
Change history
04 September 2018
A correction to this article has been published and is linked from the HTML and PDF versions of this paper. The error has not been fixed in the paper.
References
Jafri, R. Z. et al. Global epidemiology of invasive meningococcal disease. Popul Health Metr 11, 17 (2013).
Caugant, D. A. & Maiden, M. C. Meningococcal carriage and disease–population biology and evolution. Vaccine 27(Suppl 2), B64–70 (2009).
Rollier, C. S., Dold, C., Marsay, L., Sadarangani, M. & Pollard, A. J. The capsular group B meningococcal vaccine, 4CMenB: clinical experience and potential efficacy. Expert Opin Biol Ther 15, 131–142 (2015).
Read, R. C. Neisseria meningitidis; clones, carriage, and disease. Clin Microbiol Infect 20, 391–395 (2014).
Shao, Z. et al. Identification of a new Neisseria meningitidis serogroup C clone from Anhui province, China. Lancet 367, 419–423 (2006).
Li, J. et al. Prevalence of meningococcal meningitis in China from 2005 to 2010. Vaccine 33, 1092–1097 (2015).
Li, J. et al. Meningococcal disease and control in China: Findings and updates from the Global Meningococcal Initiative (GMI). J Infect 76, 429–437 (2018).
Zhou, H. et al. Distribution of serogroups and sequence types in disease-associated and carrier strains of Neisseria meningitidis isolated in China between 2003 and 2008. Epidemiol Infect 140, 1296–1303 (2012).
Li, J. et al. Epidemiological characteristics of meningococcal meningitis and switching trend of serogroups of Neisseria meningitidis in China, 2006–2014. Chin J Vacc Immun 21, 481–485 (2015).
Zhou, H. et al. Spread of Neisseria meningitidis serogroup W clone, China. Emerg Infect Dis 19, 1496–1499 (2013).
Shirley, M. & Dhillon, S. Bivalent rLP2086 Vaccine (Trumenba((R))): A Review in active immunization against invasive meningococcal group B disease in individuals aged 10–25 years. BioDrugs 29, 353–361 (2015).
Medini, D., Stella, M. & Wassil, J. MATS: Global coverage estimates for 4CMenB, a novel multicomponent meningococcal B vaccine. Vaccine 33, 2629–2636 (2015).
Poolman, J. T. & Richmond, P. Multivalent meningococcal serogroup B vaccines: challenges in predicting protection and measuring effectiveness. Expert Rev Vaccines 14, 1277–1287 (2015).
Ladhani, S. N. et al. Effectiveness of Meningococcal B Vaccine against Endemic Hypervirulent Neisseria meningitidis W Strain, England. Emerg Infect Dis 22, 309–311 (2016).
Brehony, C. et al. Distribution of Bexsero(R) Antigen Sequence Types (BASTs) in invasive meningococcal disease isolates: Implications for immunisation. Vaccine 34, 4690–4697 (2016).
McNeil, L. K. et al. Role of factor H binding protein in Neisseria meningitidis virulence and its potential as a vaccine candidate to broadly protect against meningococcal disease. Microbiol Mol Biol Rev 77, 234–252 (2013).
Zhu, P. et al. Fit genotypes and escape variants of subgroup III Neisseria meningitidis during three pandemics of epidemic meningitis. Proc Natl Acad Sci USA 98, 5234–5239 (2001).
Nicolas, P. et al. Clonal expansion of sequence type (ST5)- and emergence of ST-7 in serogroup A meningococci, Africa. Emerg Infect Dis 7, 849–854 (2001).
Zhou, H. et al. Clonal characteristics of invasive Neisseria meningitidis following initiation of an A + C vaccination program in China, 2005–2012. J Infect 70, 37–43 (2015).
Sridhar, S. et al. Global incidence of serogroup B invasive meningococcal disease: a systematic review. Lancet Infect Dis 15, 1334–1346 (2015).
Shu, Y. L. et al. Dual seasonal patterns for influenza, China. Emerg Infect Dis 16, 725–726 (2010).
Taha, M. K. et al. Serogroup W135 meningococcal disease in Hajj pilgrims. Lancet 356, 2159 (2000).
Fan, K. W. Epidemic cerebrospinal meningitis during the Cultural Revolution. Extrême-Orient Extrême-Occident 37, 197–232 (2014).
Wang, L., Zhang, Q., Cai, Y., Zhang, J. & Ma, Q. Simulation study of pedestrian flow in a station hall during the Spring Festival travel rush. Physica A: Statistical Mechanics and its Applications 392, 2470–2478 (2013).
Zhang, J. et al. Molecular haracteristics of Neisseria meningitidis isolated during an outbreak in a jail: Association with the spread and distribution of ST-4821 complex serogroup C clone in China. Biomed Environ Sci 26, 331–337 (2013).
Kaaijk, P. et al. Preclinical safety and immunogenicity evaluation of a nonavalent PorA native outer membrane vesicle vaccine against serogroup B meningococcal disease. Vaccine 31, 1065–1071 (2013).
Parikh, S. R. et al. Meningococcal serogroup B strain coverage of the multicomponent 4CMenB vaccine with corresponding regional distribution and clinical characteristics in England, Wales, and Northern Ireland, 2007-08 and 2014-15: a qualitative and quantitative assessment. Lancet Infect Dis 17, 754-762 (2017).
Taha, M. K. et al. Bactericidal activity of sera from adolescents vaccinated with bivalent rLP2086 against meningococcal serogroup B outbreak strains from France. Vaccine 35, 1530–1537 (2017).
Ostergaard, L. et al. A bivalent meningococcal B vaccine in adolescents and young adults. N Engl J Med 377, 2349–2362 (2017).
Lujan, E., Partridge, E., Giuntini, S., Ram, S. & Granoff, D. M. Breadth and duration of meningococcal serum bactericidal activity in healthcare workers and microbiologists immunized with the MenB-FHbp vaccine. Clin Vaccine Immunol 24, e00121–00117 (2017).
Shi, F. et al. Prevalence of factor H binding protein sub-variants among Neisseria meningitidis in China. Vaccine 35, 2343–2350 (2017).
Vogel, U. et al. Predicted strain coverage of a meningococcal multicomponent vaccine (4CMenB) in Europe: a qualitative and quantitative assessment. Lancet Infect Dis 13, 416–425 (2013).
Li, Y., Zeng, M., Wang, X. & Wang, D. Clinical epidemiology of epidemic cerebrospinal meningitis in children in Shanghai area from 1984 to 2003. Chin. J Infect Chemother 4, 321–323 (2004).
Feng, L., Chengshu, D. & Daiming, W. Clinical epidemiology and clinical characterization of serogroup B meningococcal infection cases. J Practi Pediatr 2, 22–23 (1987).
Ma, Y., Shen, X., Yang, Y. & Luo, Z. Clinical analysis on children with group B epidemic meningitis. J Clin Pediatr 21, 461–463 (2003).
Chen, M. et al. Shifts in the antibiotic susceptibility, serogroups, and clonal complexes of Neisseria meningitidis in Shanghai, China: A time trend analysis of the pre-Quinolone and Quinolone Eras. PLoS medicine 12, e1001838, https://doi.org/10.1371/journal.pmed.1001838 (2015).
Zhu, H. et al. Development of a multiplex PCR assay for detection and genogrouping of Neisseria meningitidis. J Clin Microbiol 50, 46–51 (2012).
Jolley, K. A. & Maiden, M. C. BIGSdb: Scalable analysis of bacterial genome variation at the population level. BMC bioinformatics 11, 595 (2010).
Brehony, C., Wilson, D. J. & Maiden, M. C. Variation of the factor H-binding protein of Neisseria meningitidis. Microbiology 155, 4155–4169 (2009).
Russell, J. E., Jolley, K. A., Feavers, I. M., Maiden, M. C. & Suker, J. PorA variable regions of Neisseria meningitidis. Emerg Infect Dis 10, 674–678 (2004).
Bambini, S. et al. Neisseria adhesin A variation and revised nomenclature scheme. Clin Vaccine Immunol 21, 966–971 (2014).
Harris, S. L. et al. Neisseria meningitidis serogroup B vaccine, bivalent rLP2086, induces broad serum bactericidal activity against diverse invasive disease strains including outbreak strains. Pediatr Infect Dis J 36, 216–223 (2017).
Charlene, M. C. R. et al. Genomic surveillance of 4CMenB vaccine antigenic variants among disease-causing Neisseria meningitidis isolates, United Kingdom, 2010–2016. Emerg Infect Dis 24, 673–682 (2018).
Acknowledgements
This study was supported by grants from National Natural Science Foundation of China (81601801), Shanghai Rising-Star Program and Natural Science Foundation of Shanghai from Shanghai Municipal Science and Technology Commission (17QA1403100 and 16ZR1433300), a Municipal Human Resources Development Program for Outstanding Young Talents in Medical and Health Sciences in Shanghai (2017YQ039), the 4th Three-year Action Plan for Public Health of Shanghai (15GWZK0101) from Shanghai Municipal Commission of Health and Family Planning, and the 13th Five-Year Project of National Health and Family Planning Commission of the People’s Republic of China (2017ZX10303405004 and 2017ZX10103009-003). CMCR was supported by a Wellcome Trust Clinical Fellowship (087622/Z/08/2) and OBH and MCJM by the Wellcome Trust Institutional Strategic Support Fund (H2RXJo00). The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: M.C.J.M. and M.C. Performed the experiments: M.L.C. and C.Z. Analysed the data: M.L.C., C.M.C.R. and O.B.H. Contributed reagents/materials/analysis tools: T.T., J.C. and M.C. Wrote the first draft of the manuscript: M.L.C. Contributed to the writing of the manuscript: C.M.C.R., O.B.H. and M.C.J.M. Agree with manuscript results and conclusions: M.L.C., C.M.C.R., O.B.H., C.Z., T.T., J.C., X.Z., M.C. and M.C.J.M. All authors have read and confirm that they meet I.C.M.J.E. criteria for authorship.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chen, M., Rodrigues, C.M.C., Harrison, O.B. et al. Invasive meningococcal disease in Shanghai, China from 1950 to 2016: implications for serogroup B vaccine implementation. Sci Rep 8, 12334 (2018). https://doi.org/10.1038/s41598-018-30048-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-30048-x
This article is cited by
-
Impfstoffe gegen Meningokokken für das Kindesalter
Monatsschrift Kinderheilkunde (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
