
Abstract
It is increasingly recognized that synthesis and turnover of cardiac triglyceride (TG) play a pivotal role in the regulation of lipid metabolism and function of the heart. The last step in TG synthesis is catalyzed by diacylglycerol:acyltransferase (DGAT) which esterifies the diacylglycerol with a fatty acid. Mammalian heart has two DGAT isoforms, DGAT1 and DGAT2, yet their roles in cardiac metabolism and function remain poorly defined. Here, we show that inactivation of DGAT1 or DGAT2 in adult mouse heart results in a moderate suppression of TG synthesis and turnover. Partial inhibition of DGAT activity increases cardiac fatty acid oxidation without affecting PPARα signaling, myocardial energetics or contractile function. Moreover, coinhibition of DGAT1/2 in the heart abrogates TG turnover and protects the heart against high fat diet-induced lipid accumulation with no adverse effects on basal or dobutamine-stimulated cardiac function. Thus, the two DGAT isoforms in the heart have partially redundant function, and pharmacological inhibition of one DGAT isoform is well tolerated in adult hearts.
Similar content being viewed by others
Introduction
Cardiac triglyceride (TG) accumulation is a common clinical feature observed in older individuals and patients with diabetes1,2. Although increased cardiac TG content has previously been considered as a marker of lipotoxic cardiomyopathy, recent work showed that enhancing cardiac triglyceride production by diacylglycerol:acyltransferase 1 (DGAT1) overexpression was protective against lipotoxicity and ischemia-reperfusion injury3,4. These findings suggest that cardiac lipid turnover plays an important role in stress response. When cardiac TG lipolysis is blocked by deletion of adipose triglyceride lipase (ATGL) it results in a dramatic steatosis, cardiac dysfunction and early death5. Alternatively, increasing cardiac lipolysis by overexpression of ATGL protects against pressure overload and diabetic cardiomyopathy6,7. These data suggest that TG synthesis followed by breakdown, otherwise known as TG turnover, plays a pivotal role in cardiac physiology and diseases. Although studies targeting TG metabolism through ATGL or DGAT1 overexpression have been enlightening, few studies exist that have evaluated the effect of decreased TG production on cardiac metabolism and health. This is an important issue as inhibition of TG synthesis is being considered as a therapeutic measure for hyperlipidemia8,9,10.
The final step in TG synthesis is catalyzed by DGAT 1 and 2, both of which are present in the heart. The DGAT1-null mice were previously shown to be resistant to diet-induced obesity with no cardiac phenotype, mainly due to reduced lipid uptake in the intestine9,10,11,12. The DGAT1-null mice had normal levels of TG and diacylglycerol (DAG) in the heart10 while cardiac specific deletion of DGAT1 was sufficient to increase DAG levels without effecting TG levels13. The in vivo rate of TG synthesis or turnover in the heart has never been assessed in these models. The relative contribution of the two DGAT isoforms in cardiac triglyceride metabolism has not been addressed either.
In addition to protection against lipotoxicity, TG turnover has been suggested to be critical for modulation of fatty acid oxidation via regulating the transcriptional activity of peroxisome proliferator-activated receptor α (PPARα), possibly by contributing a fatty acid ligand. Previous studies have shown that either extracellular TG lipolysis by lipoprotein lipase (LPL) or intracellular TG lipolysis by ATGL can increase PPARα activity5,14,15. In rodent hearts, TG turnover rate was shown closely correlated with PPARα activity and cardiac function5,14,15. In failing human hearts, DGAT expression was shown to be decreased which was considered a pathogenic mechanism for decreased fatty acid oxidation and lipotoxicity16.
In the present study we determined the role of DGAT 1 and 2 in TG synthesis and turnover in the heart as well as their contributions to cardiac fatty acid metabolism. Utilizing an inducible cardiac-specific DGAT1 deletion mouse model (iKO) together with DGAT2-specific inhibitor, we were able to achieve graded inhibition of TG synthesis and turnover in adult mouse hearts. We observed that deletion of DGAT1 only modestly reduced TG synthesis from exogenous fatty acids which correlated with increased fatty acid oxidation and had no effect on cardiac function up to 1 year of age. Furthermore, we found that DGAT2 compensated for DGAT1 function in iKO and that inhibition of both isoforms abrogated TG synthesis but did not alter cardiac response to high fat diet.
Results
Deletion of DGAT1 has no effect on cardiac TG content and function
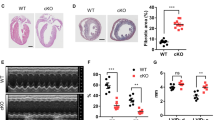
To investigate the role of DGAT1 in the heart, we generated cardiac-specific constitutive and inducible DGAT1 KO mouse models (cKO and iKO, respectively). Both models showed reduced DGAT1 mRNA (Fig. 1A) and protein (Fig. 1B) with no effects on DGAT2 mRNA expression or cardiac TG content (Fig. 1A,D). As evidenced by electron microscopy, there were no differences between genotypes for cardiac lipid droplet number and morphology (Fig. 1E–G). Relative to control mice, echocardiography analysis showed no differences in cardiac geometry or function of cKO and iKO mice for up to 12 months (Fig. 1H, Supplemental Table 2). To focus on the effects of DGAT inhibition in adult hearts, we used iKO model for the rest of the study.

Triglyceride storage is unaffected in DGAT1 inducible knockout (iKO) mice. Effective knockdown of DGAT1 in the heart was achieved as evidenced by significant reduction in DGAT1 mRNA (A) and protein (B), which was not accompanied by a concomitant increase in DGAT2 mRNA (A). Although DGAT1 mRNA was reduced, there was no change in cardiac triglyceride content in iKO hearts compared to control (D). Electron microscopic analysis also revealed the presence of lipid droplet in iKO hearts at comparable sizes to control (E–G). Fractional shortening and left ventricular posterior wall thickness demonstrated unaltered cardiac function in cardiac-specific constitutive (cKO) and iKO mice (H). Full-length blots are presented in Supplementary Figure 4. Data are presented as mean ± SEM (n = 3–11). *P < 0.05 vs. control.
Cardiac TG synthesis is modestly reduced with loss of DGAT1
Using 13C NMR spectroscopy we examined fatty acid incorporation rate into the TG pool of isolated hearts perfused with 13C labeled substrates. The iKO showed a 30% lower rate of TG synthesis from exogenous fatty acids than the control (Fig. 2A). GC-MS analysis of cardiac triglycerides extracted from the heart at the end of labeling confirmed reduction in 13C labeled fatty acids in the TG pool (Fig. 2B). Moreover, total cardiac TG content was unaffected by inducible DGAT1 deletion (Fig. 2C). Collectively, these data show a modest decrease in cardiac TG turnover following DGAT1 deletion in adult hearts.

TG formation in the perfused heart is significantly reduced in inducible DGAT1 knockout animals. 13C labeling of the triglyceride pool in perfused hearts from iKO animals revealed that loss of DGAT1 reduces TG synthesis rate (A). GCMS analysis of the triglyceride pool confirmed only a modest reduction of 13C labeling in iKO (B). This was observed while TG levels remained constant during the perfusion period (C). Data are presented as mean ± SEM (n = 3–9). *P < 0.05 vs. control at a given time point.
Increased oxidation of exogenous fatty acids in DGAT1 iKO hearts
To assess the effects of reduced TG turnover on substrate oxidation, the relative contribution of 13C labeled fatty acids and glucose along with unlabeled substrates to the tricarboxylic acid (TCA) cycle was assessed by 13C-NMR techniques. Compared to control, increased contribution from exogenous fatty acid and reduced glucose contribution to total oxidative metabolism was observed in iKO hearts (Fig. 3A). Moreover, there was a negative correlation between fatty acid oxidation and the incorporation of exogenous fatty acids into the TG pool (Fig. 3B), suggesting that decreased TG synthesis is associated with increased oxidation of exogenous fatty acids. These changes in fatty acid metabolism did not affect cardiac function or energetics in the perfused iKO hearts (Fig. 3C,D). Additionally, these hearts responded normally to increased workload in vivo induced by dobutamine (Fig. S1). These data suggest that adult hearts are able to adapt to a moderate decrease in TG turnover caused by DGAT1 deletion and are able to maintain cardiac energetics or function.

Inducible DGAT knockout alters cardiac metabolism and reduces triglyceride fatty acid incorporation. 13C isotopomer analysis of glutamate revealed increased fatty acid and decreased glucose oxidation in iKO hearts compared to control (A). A Pearson’s correlation coefficient shows a strong correlation between TG incorporation and fatty acid oxidation (B), suggesting these two pathways are interconnected in governing the overall fate of fatty acid in the heart. No alteration in cardiac function (C) or energetics (D) was observed in the perfused hearts in between genotypes. Data are presented as mean ± SEM (n = 4–8). In (A) *P < 0.05 vs. control.
DGAT1 iKO hearts respond normally to high fat diet
To evaluate the effect of reduced TG synthesis on lipid loading-induced metabolic remodeling, we subjected control and iKO mice to a high fat diet (HFD, 60% kcal from fat) for 5 weeks. Body weight gains were similar in control and iKO mice after the 5 week HFD (Fig. S2A-B). Cardiac TG or DAG content did not change in control or iKO hearts after 5 weeks of a high fat diet (Fig. 4A,B). However, the 5-week HFD was sufficient to stimulate PPARα activity and increase fatty acid oxidation as previously described17. Interestingly, increases in PPARα activity and fatty acid oxidation were similar in iKO and control hearts (Fig. 4C,D) suggesting that the modest reduction in triglyceride turnover did not affect hearts’ ability to activate PPARα and increase fatty acid oxidation in response to increased lipid load.

DGAT1 deficiency shows no effect on cardiac lipid metabolism after a short term high fat diet (HFD) feeding. Cardiac TG (A) and DAG (D) contents were not different between control and iKO mice following HFD. PPARα target gene expression (C) and fatty acid contribution to total oxidative metabolism (D) were not different between WT and iKO mice in response to HFD. The dotted line indicates the level of chow-fed controls. Data are presented as mean ± SEM (n = 6–10). *P < 0.05 vs. control - Control Diet.
Role of DGAT2 in cardiac TG metabolism
Considering relatively mild effects of DGAT1 deficiency on cardiac lipid metabolism, we next sought to explore the relative contribution of the two DGAT isoforms in the heart. Quantitative measurements of the mRNA levels showed greater abundance of DGAT2 mRNA in the heart compared to DGAT1 (~1 vs. ~0.25 pmol/mg RNA respectively, Fig. 5A). To evaluate the contribution of DGAT2 to TG synthesis, we perfused hearts from control or iKO mice with DGAT2 inhibitor (5 µM)18. The DGAT2 inhibitor alone has very modest effect but inhibition of both isoforms substantially reduced 13C fatty acid incorporation into TG pool in the heart as measured by NMR spectroscopy and GCMS (Fig. 5B,C). Moreover, perfusion with DGAT2 inhibitor for 60 minutes did not alter cardiac TG levels in control or iKO hearts (Fig. S3A) suggesting that inhibition of both DGAT isoforms effectively abrogated TG turnover in the heart. Furthermore, we found that inhibition of TG turnover resulted in a rapid downregulation of PPARα target gene expression with no change in cardiac function (Fig. 5D and Fig. S3B), and DGAT2 inhibitor did not further change substrate oxidation in iKO during the perfusion period (Fig. S3C).

Contribution of individual DGAT isoforms to triglyceride metabolism. mRNA quantification of both DGAT isoforms in the mouse heart (A), 13C labeling of TG pool in the control and iKO hearts perfused with DGAT2 inhibitor (B), mass spectrometry analysis of 13C labeling of TG pool in control and iKO hearts perfused with DGAT2 inhibitor (C) and PPARα signaling between WT and iKO hearts perfused with 5 µM DGAT2 inhibitor (D). Data are presented as mean ± SEM (n = 3–7). *P < 0.05 vs. control
Cardiac effects of DGAT1/2 inhibition in vivo
To explore how DGAT1/2 coinhibition modulates in vivo cardiac lipid metabolism, we treated iKO mice with DGAT2 inhibitor for 5 days. Under fasting and fed conditions, both control and iKO mice treated with DGAT2 inhibitor had lower cardiac triglyceride content compared to vehicle treated controls (Fig. 6A). To investigate whether prolonged DGAT1/2 coinhibition modifies cardiac lipid metabolism and function during sustained lipid overload we subjected mice to 5-week HFD. Treatment with DGAT2 inhibitor in mice on control diet showed a modest effect on body weight with no significant change of blood TG level. In HFD-fed mice, however, DGAT2 inhibitor reduced body weight gain and protected against the increase of blood TG level (Fig. 6B,C). Cardiac TG was also lower in iKO mice treated with DGAT2 inhibitor (Fig. 6D). The downregulation of cardiac TG level did not affect cardiac PPARα signaling or its response to HFD (Fig. 6E). Interestingly, while basal cardiac function was unaltered between genotypes on control diet or HFD, dobutamine-induced increase in ejection fraction was attenuated in mice subjected to the HFD which was not altered by DGAT inhibition (Fig. 6F).

Coinhibition of DGAT1 and DGAT2 during high fat diet (HFD). Cardiac TG level was decreased with DGAT2 inhibition or with iKO + DGAT2 inhibition under fed and fasting conditions (A). Coinhibition of DGAT1 and DGAT2 attenuated body weight gain (B) and reduced serum (C) and cardiac (D) TG content following HFD. PPARα signaling (E) and basal cardiac function (F) were unaffected in iKO + DGAT2 mice on a HFD compared to control. Dobutamine-induced increase in cardiac function was attenuated by HFD with no differences between genotypes (F). Data are presented as mean ± SEM (n = 4–8). *P < 0.05, **P < 0.01, ***P < 0.001. †P < 0.05 vs. corresponding control group, §P < 0.05 mean effect of diet. Data analyzed by one-way ANOVA (A) and two-way ANOVA (B–F).
Discussion
In this study we show that both isoforms of DGAT contribute to triglyceride synthesis in the heart with partially redundant function. Loss of DGAT1 in the adult mouse heart exerts a moderate inhibition on cardiac triglyceride synthesis accompanied by increased oxidation of exogenous fatty acids. Inhibition of both enzymes abrogates triglyceride synthesis and turnover in the perfused heart leading to the downregulation of PPARα target gene expression. Treatment with DGAT2 inhibitor in mice with cardiac-specific DGAT1 deficiency prevents TG accumulation in the heart during 5-week high fat diet. It however, does not change the PPARα signaling or cardiac performance under control or high fat diet conditions.
It has been well recognized that myocardial TG pool is highly dynamic although the heart has very limited capacity for lipogenesis. The exact role of the TG turnover is not entirely clear but several potential functions have been proposed including preventing the generation of lipotoxic species during lipid overload, and supply fuel for oxidation upon energy stress. Several recent studies have suggested that TG lipolysis may regulate fatty acid oxidation by supplying ligands for PPARα activation5. Critical evidence in support of this notion was derived from mice with ATGL deletion in which blockade of TG lipolysis resulted in decreased fatty acid oxidation that could be rescued by pharmacological activation of PPARα. However, blockade of TG lipolysis also causes substantial steatosis that confounds the ATGL deletion phenotype. In this study, we find that TG turnover can be inhibited by targeting TG synthesis with no change of TG pool size. Using this approach, we show that partial inhibition of TG turnover by deleting DGAT1 in adult mouse heart does not impair PPARα transcriptional activity. The heart adapts to the decreased TG synthesis by increasing oxidation of the exogenous fatty acids without changes in lipid metabolism homeostasis or myocardial energetics. In response to high fat diet, not only does DGAT1 deficiency not affect cardiac function, but the iKO heart retains its ability to upregulate PPARα activity and remodel metabolism similar to that of the WT. This is unexpected given the growing body of evidence suggesting a close association of TG metabolism and PPARα activity with either TG turnover regulating PPARα activity5,15,19, or PPARα activity itself increasing TG turnover20. Furthermore, contrary to the previous studies suggesting that reduced TG synthesis and turnover would increase lipotoxic species in the heart3,13,21, DAG content in the iKO hearts did not change at baseline or following a high fat diet. These observations have led us to conclude that inhibition of DGAT1 in the adult heart exert insufficient effect on TG synthesis rates to impair cardiac function, lipid homeostasis, or PPARα activation.
Previous studies have revealed important differences in structure and subcellular localization of the two DGAT enzymes, suggesting that DGAT1 and 2 participate in different pathways of TG synthesis22,23. A model based on this premise proposes that DGAT1 is primarily involved in esterifying exogenous fatty acids to replenish the TG pool while DGAT2 is responsible for incorporating endogenously synthesized fatty acids into TG22. Since the heart is not a lipogenic organ, DGAT1 would be expected to play a primary role in TG synthesis. Interestingly, DGAT2 is highly expressed in the heart but its function has not been assessed due to neonatal lethality of DGAT2 knockout mice24. Using dynamic 13C-NMR technique, here we show that the two enzymes have partially redundant function in the heart; inhibition of either enzyme only modestly affects the rate of fatty acids incorporation into the TG pool. Inhibition of both DGAT activities results in near complete loss of TG synthesis in the heart. This is consistent with the profound reduction of TG in fetal mice lacking both enzymes25, thus, supporting the notion that these two DGATs are jointly responsible for TG synthesis in the heart. Compensation by DGAT2 is likely an adaptive mechanism for DGAT1 deficiency in the heart. Although deletion of DGAT1 in the adult mouse hearts does not trigger any increases of DGAT2 mRNA level, we are unable to determine the protein level or post-translational modification of DGAT2 in the iKO hearts due to the lack of DGAT2-specific antibody. Recent studies have shown that ubiqutination or translocation of DGAT2 could change its stability or function in non-cardiac cells26, whether a similar mechanism contributes to cardiac triglyceride synthesis warrants further investigation.
Targeting TG synthesis has been proposed as a therapeutic strategy for hyperlipidemia for more than a decade9. Multiple small molecular inhibitors of DGAT, especially DGAT2, have been developed recently18,27,28. However, previous studies have suggested that inhibition of TG synthesis and turnover in the heart could contribute to lipotoxicity and hence cautioned the use of pharmacological inhibition of DGAT. In contrast, we find that inhibition of one isoform of DGAT in the adult heart is well tolerated. Once a near complete inhibition of TG synthesis and turnover is achieved by perfusing iKO hearts with DGAT2 inhibitor, expression of PPARα target genes is significantly downregulated. Inhibition of both DGAT isoforms in vivo, although effectively reduces TG content in the heart, do not affect PPARα activity or cardiac function under normal or high fat diet condition. We speculate that pharmacological inhibition of DGAT2 in the heart is only intermittent in vivo, thus resulting in a state of partial suppression of TG turnover despite the reduction of TG level. Collectively, these observations suggest that a low rate of TG turnover is sufficient to maintain PPARα activity, and furthermore, adult hearts can adapt to partial inhibition of DGAT activity. It remains to be determined whether this degree of DGAT inhibition affects cardiac responses to other chronic stresses such as pressure overload.
Conclusions
In the present study, we find that both isoforms of DGAT contribute to cardiac TG synthesis with partial redundancy. Coinhibition of DGAT1 and 2 in the heart is necessary to abrogate cardiac TG turnover. Moreover, inhibiting DGAT1 and 2 in the heart during chronic, sustained lipid loaded protects against cardiac lipid accumulation without significant changes of cardiac function.
Methods
Animal models
All animal procedures were approved by the University of Washington Institutional Animal Care and Use Committee (IACUC) and were performed in accordance with IACUC guidelines and regulations. DGAT1 flox/flox mice were purchased from Jackson Labs and crossed with either α-MHC-MerCreMer (MCM) transgenic mice to generate cardiac-restricted inducible DGAT1 knockout (iKO) or with α-MHC-CRE transgenic mice to generate cardiac-specific DGAT1 knockout (cKO). Age and sex-matched littermate DGAT1 flox/flox mice were used as controls. For iKO mice, tamoxifen (20 mg/kg/d, i.p.) was given for 5 days to 8-week old MCM-DGAT1 flox/flox mice and mice were analyzed 4 weeks after final tamoxifen injection. For high fat diet treatment, 3-month old mice were fed a diet with 60.3% kcal from fat (HFD, Research Diets TD.09766) or a control diet with 16.8% kcal from fat (CD, Research Diets TD.120455) for 5 weeks. For DGAT2 inhibitor treatment, control or iKO mice 4 weeks after tamoxifen treatment were orally gavaged (30 mg/kg, BID) of the DGAT2 inhibitor PF-06424439-27 in 0.5% methylcellulose for 5 days and tissues harvested with or without overnight fasting. Separate cohort of mice on HFD or CD were treated with the same dose of PF-06424439-27 for 5 weeks. All mice were kept on a 12 h light/dark cycle with ad libitum access to food and water.
Echocardiography
Transthoracic echocardiography was collected from mice anesthetized with isoflurane (1–2%) using a Vevo 2100 (VisualSonics) in short axis mode. Dobutamine stress echo was conducted by administration of dobutamine (3 mg/kg, i.p.) immediately after the baseline function assessment and repeating the echocardiographic measurements 10 minutes after dobutamine injection. Data for each mouse is an average of 3 cardiac cycles taken at comparable heart rates.
Perfusion and Substrate Utilization
Langendorf perfusion was carried out as previously described4,29,30. Mice were heparinized (100 U/mouse), anesthetized with pentobarbital (190 mg/kg, I.P.), hearts excised and perfused at 80 mmHg perfusion pressure at 37 °C with a KH buffer containing the following in mM: 0.5 EDTA, 5.3 KCl, 1.2 MgSO4, 118 NaCl, 25 NaHCO3, 2 CaCl2, 0.4 mixed long chain fatty acids, 5.5 glucose, 1.2 lactate and 50 µU/mL insulin. Ex vivo cardiac phosphocreatine (PCr) and ATP contents were determined using 31P NMR as previously described4. Contribution of individual substrates to total oxidative metabolism was determined using 13C NMR4. Specifically, hearts perfused with 1,6-13C glucose and U-13C mixed fatty acids were freeze-clamped following 40 min labelling perfusion, tissues were powdered and metabolites extracted with perchloric acid. Proton-decoupled 13C NMR spectra of tissue extracts were acquired on a 14 T magnet using TopSpin software (Bruker 600; Billerica, MA) and contributions of labeled (glucose and fatty acids) and unlabeled (exogenous lactate and endogenous glycogen and TG) substrates to total oxidative metabolism were determined by modeling of citric acid cycle flux using the glutamate C3 and C4 isotopomer peak areas (tcaCALC, Dallas, TX).
TG synthesis
Incorporation of 13C-labeled mixed fatty acids into the TAG pool was analyzed by monitoring the resonance of the methylene carbon at ~31 ppm via 13C NMR spectroscopy as previously described4. Proton-decoupled carbon spectra were determined on isolated hearts perfused with 13C-labeled mixed fatty acids over a 60-min period. Peak areas of the methylene resonance of each sequential spectrum were quantified using software from Advanced Chemistry Development Laboratories (ACD Laboratories, Toronto, Ontario, Canada) and corrected for 13C natural abundance.
Electron Microscopic Analysis
Thin heart sections were cut and rinsed in a KH buffer containing 20 mM KCl to arrest the muscle in diastole. Sections were immediately placed into a fixation solution containing 0.4% glutaraldehyde in 0.1 M sodium cacodylate buffer. Fixed tissue samples were processed and images taken using a Transmission Electron Microscope (JEOL1230). Lipid droplet number and morphology were quantified using ImageJ analysis software. Data for each heart were calculated as average value of 40 random images.
Biochemical Assays
Total lipids were extracted using the Folch extraction procedure and quantified using the sigma colorimetric assay kit as previously described31. Briefly, heart tissues were weighed and homogenized in 2:1 chloroform:methanol, incubated on ice for 30 min, then centrifuged at 3000 rpm for 12 minutes. The top layer was discarded, the bottom layer was dried under N2 gas and reconstituted in 1% Triton X-100:Isopropanol. Individual lipid classes were extracted as described above were loaded onto an NH2 column (Agilent BondElut) preconditioned with hexane and neutral lipids were extracted with 2:1 Chlorofom:Isopropanol, dried under N2 gas, reconstituted in hexane and loaded onto another preconditioned NH2 column4,32. TG were eluted from the column by the addition of 1% diethyl ether:10% methylene chloride in hexane and DAG with 15% ethyl acetate in hexane dried under N2 gas. DAG was quantified using the triglyceride assay kit with a DAG standard curve. Fatty acid composition and 13C labeling patterns was assessed using a gas chromatograph mass spectrometer (GCMS, Shimadzu QP2010) with a 0.25 mm inner diameter column (Restek Rtx-225) on fatty acid methyl esthers (FAMES) made from the triglyceride fraction obtained using solid phase separation as previously described33.
Real Time PCR
RNA from frozen heart tissue was isolated using the RNeasy fibrous extraction kit (Qiagen) and cDNA was synthesized using the Omniscript (Qiagen) reverse transcriptase according to the manufacturer’s recommendations. PCR reactions containing equal cDNA concentrations were set up in duplicate using Sybr green (BioRad) on a rotogene using concentrated cDNA as a standard. All values were normalized to 18 S concentration and expressed as a fold change from control. Primer sequences for all genes are described in Supplemental Table 1. Quantitative measurements of DGAT1 and DGAT2 levels in mouse heart tissue were performed based on standard curves generated using full-length amplicons supplied by Integrated DNA technologies.
Western blotting
Whole heart proteins were extracted from flash frozen hearts using 10–20 mg of wet weight tissue using RIPA buffer containing 150 mM NaCl, 50 mM Tris-HCl, 1% Triton X-100, 0.1% SDS, 0.1% sodium deoxycholate and 1 mM EDTA, pH 7.4. Briefly, frozen hearts were homogenized in RIPA buffer using bullet blender and placed on ice for 30 min. Next, samples were centrifuged for 10 min at 4 °C at 20,000 g. Supernatant was collected and its protein content determined using BCA assay kit (BCA protein assay kit, Pierce Biotechnology, USA). Protein samples were run a 10% Bis-Tris acrylamide gel for 2 h at constant 100 V and transferred on a polyvinylidenedifluoride membrane (PVDF) for 2 h at constant 250 mA in an ice-chilled transfer tank. The membrane was blocked and incubated overnight at 4 °C with mouse anti-DGAT1 antibody (Santa Cruz, #sc-271934) and rabbit anti-GAPDH antibody (Cell Signaling, #D16H11). Membranes were then washed and incubated with corresponding anti-mouse or goat anti-rabbit HRP-conjugated secondary antibody (R&D Systems). After washing membranes were incubated with enhanced chemiluminesence (ECL) detection solution (Supersignal Westpico Chemiluminescent Substrate, Thermo, #34080) and exposed to X-ray film (HyBlot CL, Denville Scientific, Inc., NJ, US). Intensity was measured by ImageJ densitometry software.
Statistics
All data are presented as mean ± SEM. Statistical analysis was conducted using a student’s t-test for experiments with 2 groups, one-way ANOVA was used for experiments of three or more groups. Two-way ANOVA was used for experiments with repeated measures as specified in the figure legend. Graphs and statistical analyses were performed using Graph Pad Prism 6.0. Statistical significance was set at P < 0.05.
Data availability
All data generated or analyzed during this study are included in this published article (and its Supplementary Information files).
References
McGavock, J. M. et al. Cardiac steatosis in diabetes mellitus: a 1H-magnetic resonance spectroscopy study. Circulation 116, 1170–1175, https://doi.org/10.1161/CIRCULATIONAHA.106.645614 (2007).
van der Meer, R. W. et al. The ageing male heart: myocardial triglyceride content as independent predictor of diastolic function. European heart journal 29, 1516–1522, https://doi.org/10.1093/eurheartj/ehn207 (2008).
Liu, L. et al. DGAT1 expression increases heart triglyceride content but ameliorates lipotoxicity. J Biol Chem 284, 36312–36323, https://doi.org/10.1074/jbc.M109.049817 (2009).
Kolwicz, S. C. Jr., Liu, L., Goldberg, I. J. & Tian, R. Enhancing Cardiac Triacylglycerol Metabolism Improves Recovery From Ischemic Stress. Diabetes 64, 2817–2827, https://doi.org/10.2337/db14-1943 (2015).
Haemmerle, G. et al. ATGL-mediated fat catabolism regulates cardiac mitochondrial function via PPAR-alpha and PGC-1. Nature medicine 17, 1076–1085, https://doi.org/10.1038/nm.2439 (2011).
Kienesberger, P. C. et al. Myocardial ATGL overexpression decreases the reliance on fatty acid oxidation and protects against pressure overload-induced cardiac dysfunction. Molecular and cellular biology 32, 740–750, https://doi.org/10.1128/MCB.06470-11 (2012).
Pulinilkunnil, T. et al. Myocardial adipose triglyceride lipase overexpression protects diabetic mice from the development of lipotoxic cardiomyopathy. Diabetes 62, 1464–1477, https://doi.org/10.2337/db12-0927 (2013).
Futatsugi, K. et al. Discovery and Optimization of Imidazopyridine-Based Inhibitors of Diacylglycerol Acyltransferase 2 (DGAT2). Journal of medicinal chemistry 58, 7173–7185, https://doi.org/10.1021/acs.jmedchem.5b01006 (2015).
Smith, S. J. et al. Obesity resistance and multiple mechanisms of triglyceride synthesis in mice lacking Dgat. Nature genetics 25, 87–90, https://doi.org/10.1038/75651 (2000).
Liu, L. et al. DGAT1 deficiency decreases PPAR expression and does not lead to lipotoxicity in cardiac and skeletal muscle. Journal of lipid research 52, 732–744, https://doi.org/10.1194/jlr.M011395 (2011).
Ables, G. P. et al. Intestinal DGAT1 deficiency reduces postprandial triglyceride and retinyl ester excursions by inhibiting chylomicron secretion and delaying gastric emptying. Journal of lipid research 53, 2364–2379, https://doi.org/10.1194/jlr.M029041 (2012).
Lee, B., Fast, A. M., Zhu, J., Cheng, J. X. & Buhman, K. K. Intestine-specific expression of acyl CoA:diacylglycerol acyltransferase 1 reverses resistance to diet-induced hepatic steatosis and obesity in Dgat1−/− mice. Journal of lipid research 51, 1770–1780, https://doi.org/10.1194/jlr.M002311 (2010).
Liu, L. et al. Cardiomyocyte-specific loss of diacylglycerol acyltransferase 1 (DGAT1) reproduces the abnormalities in lipids found in severe heart failure. J Biol Chem 289, 29881–29891, https://doi.org/10.1074/jbc.M114.601864 (2014).
Duncan, J. G. et al. Rescue of cardiomyopathy in peroxisome proliferator-activated receptor-alpha transgenic mice by deletion of lipoprotein lipase identifies sources of cardiac lipids and peroxisome proliferator-activated receptor-alpha activators. Circulation 121, 426–435, https://doi.org/10.1161/CIRCULATIONAHA.109.888735 (2010).
Kienesberger, P. C. et al. Early structural and metabolic cardiac remodelling in response to inducible adipose triglyceride lipase ablation. Cardiovasc Res 99, 442–451, https://doi.org/10.1093/cvr/cvt124 (2013).
Chokshi, A. et al. Ventricular assist device implantation corrects myocardial lipotoxicity, reverses insulin resistance, and normalizes cardiac metabolism in patients with advanced heart failure. Circulation 125, 2844–2853, https://doi.org/10.1161/CIRCULATIONAHA.111.060889 (2012).
Wright, J. J. et al. Mechanisms for increased myocardial fatty acid utilization following short-term high-fat feeding. Cardiovasc Res 82, 351–360, https://doi.org/10.1093/cvr/cvp017 (2009).
Imbriglio, J. E. et al. Discovery and Pharmacology of a Novel Class of Diacylglycerol Acyltransferase 2 Inhibitors. Journal of medicinal chemistry 58, 9345–9353, https://doi.org/10.1021/acs.jmedchem.5b01345 (2015).
Lahey, R., Wang, X., Carley, A. N. & Lewandowski, E. D. Dietary fat supply to failing hearts determines dynamic lipid signaling for nuclear receptor activation and oxidation of stored triglyceride. Circulation 130, 1790–1799, https://doi.org/10.1161/circulationaha.114.011687 (2014).
Banke, N. H. et al. Preferential oxidation of triacylglyceride-derived fatty acids in heart is augmented by the nuclear receptor PPARalpha. Circulation research 107, 233–241, https://doi.org/10.1161/CIRCRESAHA.110.221713 (2010).
Liu, L. et al. Diacylglycerol acyl transferase 1 overexpression detoxifies cardiac lipids in PPARgamma transgenic mice. Journal of lipid research 53, 1482–1492, https://doi.org/10.1194/jlr.M024208 (2012).
Stone, S. J., Levin, M. C. & Farese, R. V. Jr. Membrane topology and identification of key functional amino acid residues of murine acyl-CoA:diacylglycerol acyltransferase-2. J Biol Chem 281, 40273–40282, https://doi.org/10.1074/jbc.M607986200 (2006).
Yen, C. L., Stone, S. J., Koliwad, S., Harris, C. & Farese, R. V. Jr. Thematic review series: glycerolipids. DGAT enzymes and triacylglycerol biosynthesis. Journal of lipid research 49, 2283–2301, https://doi.org/10.1194/jlr.R800018-JLR200 (2008).
Stone, S. J. et al. Lipopenia and skin barrier abnormalities in DGAT2-deficient mice. J Biol Chem 279, 11767–11776, https://doi.org/10.1074/jbc.M311000200 (2004).
Harris, C. A. et al. DGAT enzymes are required for triacylglycerol synthesis and lipid droplets in adipocytes. Journal of lipid research 52, 657–667, https://doi.org/10.1194/jlr.M013003 (2011).
Wilfling, F. et al. Triacylglycerol synthesis enzymes mediate lipid droplet growth by relocalizing from the ER to lipid droplets. Developmental cell 24, 384–399, https://doi.org/10.1016/j.devcel.2013.01.013 (2013).
Kim, M. O. et al. Discovery of a novel class of diacylglycerol acyltransferase 2 inhibitors with a 1H-pyrrolo[2,3-b]pyridine core. Biological & pharmaceutical bulletin 37, 1655–1660 (2014).
Tep, S. et al. Rescue of Mtp siRNA-induced hepatic steatosis by DGAT2 siRNA silencing. Journal of lipid research 53, 859–867, https://doi.org/10.1194/jlr.M021063 (2012).
Kolwicz, S. C. Jr et al. Cardiac-specific deletion of acetyl CoA carboxylase 2 prevents metabolic remodeling during pressure-overload hypertrophy. Circulation research 111, 728–738, https://doi.org/10.1161/circresaha.112.268128 (2012).
Yan, J. et al. Increased glucose uptake and oxidation in mouse hearts prevent high fatty acid oxidation but cause cardiac dysfunction in diet-induced obesity. Circulation 119, 2818–2828, https://doi.org/10.1161/CIRCULATIONAHA.108.832915 (2009).
O’Donnell, J. M. et al. Accelerated triacylglycerol turnover kinetics in hearts of diabetic rats include evidence for compartmented lipid storage. Am J Physiol Endocrinol Metab 290, E448–455, https://doi.org/10.1152/ajpendo.00139.2005 (2006).
Carley, A. N. et al. Multiphasic triacylglycerol dynamics in the intact heart during acute in vivo overexpression of CD36. Journal of lipid research 54, 97–106, https://doi.org/10.1194/jlr.M029991 (2013).
Kaluzny, M. A., Duncan, L. A., Merritt, M. V. & Epps, D. E. Rapid separation of lipid classes in high yield and purity using bonded phase columns. Journal of lipid research 26, 135–140 (1985).
Acknowledgements
This work was supported in part by grants from the National Institution of Health, HL129510 (to RT), T32DK007247 (to NR), S10RR029021 (to 14T HRIM Facility), P30 EY01730 (to Vision Research Center Core). We thank Dale Whittington at University of Washington School of Pharmacy mass spectrometry core for the help with GC-MS analysis, and Pfizer for its gracious donation of the DGAT2 inhibitor.
Author information
Authors and Affiliations
Contributions
N.D.R., M.K.H., T.L. and R.T. conceived and planned the experiments. N.D.R., M.K.H., T.L. performed experiments and analyzed the data. N.D.R., M.K.H. and R.T. interpreted the data. N.D.R., M.K.H. and R.T. drafted and edited the manuscript. N.D.R., M.K.H., T.L. and R.T. approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Roe, N.D., Handzlik, M.K., Li, T. et al. The Role of Diacylglycerol Acyltransferase (DGAT) 1 and 2 in Cardiac Metabolism and Function. Sci Rep 8, 4983 (2018). https://doi.org/10.1038/s41598-018-23223-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-23223-7
This article is cited by
-
Cardiomyocyte-specific disruption of the circadian BMAL1–REV-ERBα/β regulatory network impacts distinct miRNA species in the murine heart
Communications Biology (2023)
-
SARS-CoV-2 exploits host DGAT and ADRP for efficient replication
Cell Discovery (2021)
-
Treatment of human skeletal muscle cells with inhibitors of diacylglycerol acyltransferases 1 and 2 to explore isozyme-specific roles on lipid metabolism
Scientific Reports (2020)
-
DGAT1 Inhibitor Suppresses Prostate Tumor Growth and Migration by Regulating Intracellular Lipids and Non-Centrosomal MTOC Protein GM130
Scientific Reports (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
