
Abstract
Classical Lotka-Volterra (LV) competition equation has shown that coexistence of competitive species is only possible when intraspecific competition is stronger than interspecific competition, i.e., the species inhibit their own growth more than the growth of the other species. Note that density effect is assumed to be linear in a classical LV equation. In contrast, in wild populations we can observed that mortality rate often increases when population density is very high, known as crowding effects. Under this perspective, the aggregation models of competitive species have been developed, adding the additional reduction in growth rates at high population densities. This study shows that the coexistence of a few species is promoted. However, an unsolved question is the coexistence of many competitive species often observed in natural communities. Here, we build an LV competition equation with a nonlinear crowding effect. Our results show that under a weak crowding effect, stable coexistence of many species becomes plausible, unlike the previous aggregation model. An analysis indicates that increased mortality rate under high density works as elevated intraspecific competition leading to the coexistence. This may be another mechanism for the coexistence of many competitive species leading high species diversity in nature.
Similar content being viewed by others

Introduction
Competition is one of the fundamental ecological interactions between species1. We can observe that coexisting species are competing for the same resources2. A typical resource competition model which has been recognized widely is the Classical Lotka-Volterra competition1,2. The analyses of this equation show that coexistence of two or more species becomes only possible if intraspecific competition is stronger than interspecific competition3. Otherwise, dynamics leads to the exclusion of one species among n species, known as the competitive exclusion principle1,4. However, in natural communities many competing species have been coexisting in the same habitat over time, resulting in a high species diversity. Hence, we suspect that there should be some mechanisms for coexistence of competitive species, e.g., spatial structures5,6. These models, however, introduce an additional complexity into the mathematical models of classical LV systems. Compared with these complex models, coexistence of multiple species in natural communities seems to be far more ubiquitous. Therefore, a more universal explanation may be worth considering.
Crowding effect is considered as one of the ubiquitous mechanisms in any biological populations7,8,9,10,11,12,13,14,15,16. A nonlinear density effect at high densities is called crowding effect, while that at low densities, Allee effect7,8,9,10,11,12,13,14,15,16,17. Unlike this nonlinear density effect, in the traditional mathematical models of population dynamics, density effect is usually treated as constant (i.e., linear), and crowding effect is not included. To consider the nonlinear density effects, aggregation models have been developed and studied extensively introducing ‘mean crowding’8,9,10,15,16. These models show the coexistence of a few species, but not many species. The ‘mean crowding’ is a statistical feature of crowding affecting population growth rate (both birth rate and mortality). Here, we develop a simple LV type competition model with crowding effect on mortality only. We assume that the mortality rate of an individual increases with the density of a population. Moreover, using our modified LV competition model with nonlinear crowding effect, we show that multiple species are generally possible to coexist using LV system with crowding effect. This coexistence dynamics should be applicable to insect or animal species.
Results
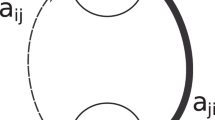
We consider the LV competition system where all interspecific competition rates are unity, i.e., α ij = α ji = 1∀i. Setting dx i /dt = 0, we obtain the zero isoclines for the modified Lotka-Volterra (LV) competition equations with nonlinear crowding effect rate m i . These isoclines are straight lines in the classical LV competition equation when we sketch the graph of the population density 1 with respect to the population density 2 (Fig. 1, dotted lines). However, we observed that these isoclines turn out to be curved when we include a nonlinear crowding effect m i (Fig. 1, solid lines). All four cases of Lotka-Volterra model show convergent-stable coexistence by adding a crowding effect, where an inferior (superior) species always increases (decreases) in densities (Fig. 1). The equilibrium point is moving from E1(d i = 0.0) to E2(d i = 0.5) which is also caused by the inclusion of a crowding effect m i , irrespective of other constant parameters (Fig. 1).

The zero isoclines and the long-term variation density of the classical LV competition equation with the inclusion of crowding effect for the four classical cases. Crowding effects (Solid line: d i = 0.5, equilibrium: E2) enable coexistence in all four cases of classical LV system (Broken line: d i = 0.0, equilibrium: E1). (a) \(({K}_{2} < \frac{{K}_{1}}{{\alpha }_{12}},{K}_{1} > \frac{{K}_{2}}{{\alpha }_{21}})\), (b) \(({K}_{2} > \frac{{K}_{1}}{{\alpha }_{12}},{K}_{1} < \frac{{K}_{2}}{{\alpha }_{21}})\), (c) \(({K}_{2} < \frac{{K}_{1}}{{\alpha }_{12}},{K}_{1} < \frac{{K}_{2}}{{\alpha }_{21}})\), and (d) \(({K}_{2} > \frac{{K}_{1}}{{\alpha }_{12}},{K}_{1} > \frac{{K}_{2}}{{\alpha }_{21}})\). Colors indicate x1(red) and x2 (blue). K i : carrying capacity of species i, and α ij : competition coefficient from species j to species i. Parameters value: b i = 1.0, d i = 0.5, mi0 = 0.1, 0 ≤ δ ≤ 5. (a) α12 = 0.8, α21 = 1.2. (b) α12 = 1.2, α21 = 0.8. (c) α12 = 0.5, α21 = 0.4. (d) α12 = 1.2, α21 = 1.3. Initial density x1 = 0.5, x2 = 0.2. We used Anaconda package of the software Python 3.6 for our simulation analysis22.
Curved lines in the \({x}_{1}-{x}_{2}\) plane imply that coexistence only take place when the value of δ → 1.0 for all d i > 0 (Fig. 2). However, by increasing δ, the competitive exclusion reappears in all exclusion cases, where the equilibrium state in \({x}_{1}-{x}_{2}\) plane is strongly curved to return to the originated axis (Fig. 2 except 2c). In contrast, an isocline is straight in \({x}_{1}-{x}_{2}\) plane, where the two species coexist without crowding effects, such that \({K}_{2} < \frac{{K}_{1}}{{\alpha }_{12}}\) and \({K}_{2} < \frac{{K}_{2}}{{\alpha }_{12}}\) (Fig. 2c). Furthermore, Fig. 3a shows that two competing species can coexist for any small positive value of δ for all d i > 0. Moreover, small positive real number δ will make coexistence possible but bigger value indicates the competitive exclusion principle again (Fig. 2 except 2c and Fig. 3). Figure 4 shows many-species Lotka-Volterra competition dynamics with crowding effects. Temporal dynamics of all species becomes convergent stable by adding crowding effects (Fig. 4a,b). The effect of δ in 5 or 10 species (Fig. 4c,d) is qualitatively same with the case of two species competition (Fig. 3b). Thus, many-species LV competition model with crowding effects leads to the stable coexistence of all species (Fig. 4).

Modified LV competition system with nonlinear crowding effect using the four possible cases of isoclines for the crowding strength factor δ. All non-coexistence patterns (a,b,d) move to coexistence as δ → 1.0 and return to competitive exclusion as δ → 5.0. The right column figures show the equilibrium points in the x1 − x2 phase plane. (a) \(({K}_{2} < \frac{{K}_{1}}{{\alpha }_{12}},{K}_{1} > \frac{{K}_{2}}{{\alpha }_{21}})\), (b) \(({K}_{2} > \frac{{K}_{1}}{{\alpha }_{12}},{K}_{1} < \frac{{K}_{2}}{{\alpha }_{21}})\), (c) \(({K}_{2} < \frac{{K}_{1}}{{\alpha }_{12}},{K}_{1} < \frac{{K}_{2}}{{\alpha }_{21}})\), and (d) \(({K}_{2} > \frac{{K}_{1}}{{\alpha }_{12}},{K}_{1} > \frac{{K}_{2}}{{\alpha }_{21}})\). Colors indicate x1(red) and x2 (blue). Parameter value and software used: see Fig. 1.

The effects of δ on population density in LV competition model with crowding effect. (a) Density ratio (x1/x2) of species 1 and 2 plotted on the crowding strength factor δ and the basic crowding component constant assuming equal between species 1 and 2 (d1 = d2). (b,c) Density of two species (x1: red, x2: blue) plotted against δ, where (b) 0 ≤ δ ≤ 1, (c) 0 ≤ δ ≤ 5, and d i = 0.3. Parameters value: b1 = 1.0, b2 = 1.8; α ij = 1.0; mi0 = 0.1. Initial density x i = 0.5. See Fig. 1 for software.

Long-term dynamics of many-species LV competition model with crowding effect. (a,b) Temporal dynamics with the basic crowding component constant (solid line: d i = 0.3; dotted line (no crowding): d i = 0.0) and the crowding strength factor δ = 0.3, (c,d) the equilibrium (final) states plotted against the effects of crowding strength factor δ (0 ≤ δ ≤ 1). (a,c) 5 species (birth rate: b i = 1.0 + 0.1(i − 1)); (b,d) 10 species (birth rate: b i = 1.0 + 0.05(i − 1)). Density of i-species: x i (i = 1, 2, …, n). Parameter value: α ij = 1.0; mi0 = 0.1. Initial density x i = 0.5. See Fig. 1 for software.
We also build LV competition models with aggregation effects on mortality that are qualitatively equivalent to the aggregation model of Hartley and Shorrocks8. We compare them with the current crowding models using the same parameter conditions (Fig. 5). In the 2-and 5-species dynamics, all species survive and converge to a stable equilibrium in both the crowding model (Fig. 5a,c) and aggregation model (Fig. 5b,d). In the 10-species dynamics, all species survive and converge to stable equilibrium in the crowding models (Fig. 5e), while only five species in the aggregation model.

Long-term dynamics of 2-, 5- and 10-species LV competition models with two types of nonlinear density effects. (a,c,e) Crowding effect (Eq. 1; d i = 0.3, δ = 0.4); (b,d,f) aggregation effect (Eq. 3; ε = 0.01). (a,b) 2 species (mi0 = 0.3); (c,d) 5 species (mi0 = 0.3); (e,f) 10 species (mi0 = 0.1). Parameter value: α ij = 1.0; b i = 1.0 + 0.01(i − 1). Initial density: (a,b) x1 = 0.15, x2 = 0.25, (c,d) x3 = 0.2, x4 = 0.12, x5 = 0.18, (e,f) x6 = 0.19, x7 = 0.22, x8 = 0.14, x9 = 0.13, x10 = 0.26. See Fig. 1 for software.
We also consider the effect of nonlinear competition terms21 with or without crowding effect. We compare them with the current crowding models using the same parameter conditions (Fig. 6). In their original model, Taylor and Crizer consider a modified Lotka-Volterra model introducing the effect of nonlinear competition on growth rate. Here, we introduce the nonlinear competition on birth rate alone, since crowding effect is introduced only on mortality rate. In this manner, we can compare the effect of these changes separately. In the 2-species dynamics, both species survive and converge to a stable equilibrium in the crowding model with linear or nonlinear competition terms (Fig. 6a,c). In the current parameter conditions, the LV competition model with nonlinear competition terms lead to the extinction of one species (Fig. 6b). Note that by changing the conditions, this model lead to the coexistence of the two species21. In the 5-species dynamics, all five species survive and converge to a stable equilibrium (Fig. 6d,e and f). Interestingly, the model with nonlinear competition terms converges to the same density for all five species (Fig. 6e), while the crowding effect lead to different densities among all species (Fig. 6d,f). In the 10-species dynamics, seven species survive and converge to stable equilibrium in the crowding models (Fig. 6g). In contrast, all ten species in the LV competition model with nonlinear terms lead to the coexistence of all species with an equal density (Fig. 6h). By combining the nonlinear competition and the crowding effect, all ten species survive and converge to different densities (Fig. 6i).

Long-term dynamics of 2-, 5- and 10-species LV competition models with crowding effect and/or nonlinear competition term. (a,d,g) Crowding effect (Eq. 1); (b,e,h) nonlinear competition term; (c,f,i) combination of both (Eq. 3). Parameter value: α ij = 1.0; b i = 1.0 + 0.1(i − 1). (a,c,d,f,g,i) d i = 0.3, δ = 0.4. (a–f) mi0 = 0.3; (g–i) mi0 = 0.1. Initial density: (a–c) x1 = 0.15, x2 = 0.25, (d–f) x3 = 0.2, x4 = 0.12, x5 = 0.18, (g–i) x6 = 0.19, x7 = 0.22, x8 = 0.14, x9 = 0.13, x10 = 0.26. All simulations run for the same time steps. See Fig. 1 for software.
Discussion
Many species compete for precisely the same limited resources to survive1,2. Gause’s exclusion principle show that multiple competing species cannot coexist in natural communities1,4,18,19. Only one species, the superior competitor, will survive and other competitors will eventually become extinct. We should note that frequency dependence does not promote the coexistence of multiple species20. Niche theory suggested that it will only become possible for the competing species to coexist if they have different niche2,3. Linear density effects show that coexistence becomes possible under very limited conditions. Hence, we search for mechanisms that will enable coexistence of competitive species5,6. As a universal and more biologically founded solution, we consider crowding effect, nonlinear density effects at high densities, in the LV competition systems.
Classical LV competition model shows that it is only possible for two species to coexist together if intraspecific competition is stronger than interspecific competition3. However, our results have shown that inclusion of crowding effect to the classical Lotka-Volterra competition system guarantees the coexistence of two or more species. We have investigated that two species can coexist when we include crowding effect to the classical LV competition system (Fig. 1 (solid lines), Fig. 2 as δ → 1.0, Fig. 3a as δ → 1.0, and Fig. 4a,b (solid lines)) compare to the results of the classical LV competition model which show the competitive exclusion principle (Fig. 1 (dotted lines) and Fig. 4a,b (dotted lines)).
Crowding effect has been recognized in many natural and experimental populations7,8,9,10,11,12,13,14,15,16. Note that if the density of population is increased a lot more than the carrying capacity, then crowding effect will kill all competing individuals. We here, introduced the intraspecific crowding effect into the Lotka-Volterra competition model. We have shown that a weak crowding effect make it possible to achieve a stable coexistence of multiple species. Our analysis implies that increased mortality under high density works as elevated intraspecific competition leading to the coexistence. This may be another ubiquitous mechanism for the coexistence of multiple species leading species diversity in nature.
We compare the current model with the aggregation model of coexistence8 (Fig. 5). Unlike the original aggregation model of Hartley and Shorrocks in which the aggregation reduces growth rates, we only include the aggregation effect on mortality rate. These examples show that the aggregation model becomes difficult to maintain the coexistence of all or many species when the number of species is increased. In contrast, the coexistence of all or many species can be easily achieved in the current crowding model even when the number of species is increased (Fig. 5a,c and e). The reason why the coexistence becomes difficult when the number of species increased in the aggregation model seems to depend on the combination of the parameters, where the coexistence region is expressed as a polygon in the n-species parameter space which can disappear easily when the number of species is increased. The logic is same with linear programming in n-dimensional space. In contrast, in the crowding model, the mortality rate of any species increases when its density approaches its carrying capacity. Because of this, increasing mortality rate near carrying capacity will keep all the species at the densities much below their carrying capacity. Thus, the intraspecific competition becomes most severe at or near carrying capacity, resulting in the stable coexistence of all species.
We also compare the effects of the nonlinear competition terms with the current crowding effects (Fig. 6). Unlike the LV model with nonlinear competition effects on growth rates21, we only include the nonlinear competition effects on birth rate alone since crowding effect is included only on mortality rate, so that these effects can be easily distinguished. In the current parameter conditions, the 2-species model results in the exclusion of one species (Fig. 6b). However, when the number of species is increased, LV model with nonlinear competition terms will lead to the coexistence of all species with an equal density regardless of the parameter combinations (Fig. 6e,h). It is not sure whether the stability can be achieved easily when the number of species increased in the nonlinear competition model. However, the equilibrium density is identical for all coexisting species in the model with nonlinear competition terms. This means that the effects of other species-specific parameters are completely cancelled by the introduction of nonlinear competition effect. In contrast, the coexistence of all or many species can be easily achieved in the current crowding model even when the number of species is increased (Fig. 6a,d and g). By combining the nonlinear competition and the crowding effect, many species survive and converge to different densities (Fig. 6c,f and i). Thus, both mechanisms can promote the coexistence of many species differently.
The most important assumption in our model of crowding effect is the increase in mortality rate at high density. Here, d i represents the proportion of crowding mortality contribution and δ is the power of crowding effect. Therefore, when d i = 0, this model reduces to the classical LV competition model. This assumption should be one natural way to include the crowding effect. However, there may be many other natural ways to include crowding effect, e.g., the aggregation model of Hartley and Shorrocks8. We have to wait for empirical studies to verify which way is actually functioned in a natural ecosystem. Note that these functional mechanisms are not exclusive of each other. Thus, the valid mechanisms may be different depending on a natural ecosystem. We should also note that the nonlinearity in the functional responses should be an important factor driving population dynamics and resulting evolution. The nonlinearity in density effect may be biologically inherent and appears as crowding effect at high densities and as Allee effect at low densities. Our studies thus show that the real competitive communities have a much more complicated dynamical system than the classical LV competition system.
Methods
Mathematical models
We consider the modified Lotka-Volterra (LV) competition equations with crowding effect rate m i for species i. In our model, we only consider competition between two species. In addition, carrying capacity K i is set to be equal to 1, i.e., K i = 1. The modified LV competition model is shown on the following equations:
where x i represents the population density of species i where i = 1, 2. In this model, parameter b i represents the birth rate of species i while α ij represents the effect of species j on i where i,j = 1, 2 and i ≠ j. The crowding effect rate m i is given by
where parameter mi0 represents the initial mortality factor of species i. Parameter d i represents the density-dependent factor of species i. In addition, the sum of the initial mortality and density-dependent factor of species i must be greater than 0 but less than or equal to 1, i.e.,0 < mi0 + d i ≤ 1. Note that, if the initial mortality factor mi0 is zero then nonlinear crowding effect rate m i will imply that the intraspecific competition is perfectly density-dependent. In addition, if d i = 0 \({\rm{\forall }}i\) then m i = m i 0 \({\rm{\forall }}i\) which will imply that the modified LV competition model is the same with the classical LV competition model.
Following equations 4 and 11 in the paper of Hartley and Shorrocks8, we arrived with the Lotka-Volterra competition model adding the effect of a few more individuals, shown on the following equations:
where ε is any positive real number and \({m}_{i0}{{x}_{i}}^{1+\varepsilon }\) is the effect of a few more individuals for all species i. Note that, we do not include the crowding effect on birth rates unlike the aggregation model of Hartley and Shorrocks8.
In addition, we also used the modified LV competition model of Taylor and Crizer21 with the inclusion of nonlinear crowding effect for two species. In their model, they add nonlinear competition terms to prevent the population of species 2 to have a smaller effect on the population of species 1 when the population density of species 1 is very small compare to the population density of species 2 and vice versa. Taylor and Crizer’s competition model with nonlinear crowding effect is shown on the following equations:
Numerical simulations
In order to determine the impact of the inclusion of nonlinear crowding effects to the classical Lotka-Volterra equation we simulate the modified LV competition equations using Anaconda package of the software Python 3.622. Initially, we determine its effect if there are two competing species in a community and later extend it up to 10 competing species. We also determine the effect when we use small and large values of δ. Moreover, we identify the right combination of d i and δ that will allow competing species to coexist. Without losing essential qualitative dynamics, we considered the following parameter ranges in our numerical simulations:
-
0 ≤ Initial of x i ≤ 1 for all i;
-
0 < α ij ≤ 1 for all i, j;
-
1 ≤ b i ≤ 2 for all i
-
mi0 = 0.1 or 0.3 for all i;
-
d i = 0,0.1,0.3 or 0.5 for all i;
-
0 ≤ δ ≤ 5; and
-
K i = 1
In addition, we compare the results of the LV competition equation (1) with nonlinear crowding effect (\({m}_{i}={m}_{i0}+{d}_{i}{x}_{i}^{\delta }\)) and without nonlinear crowding effect (m i = mi0) using the four possible cases of isoclines. The four isocline cases are:
-
\({K}_{1} < \frac{{K}_{2}}{{\alpha }_{21}},{K}_{2} < \frac{{K}_{1}}{{\alpha }_{12}}\)
-
\({K}_{1} < \frac{{K}_{2}}{{\alpha }_{21}},{K}_{2} > \frac{{K}_{1}}{{\alpha }_{12}}\)
-
\({K}_{1} > \frac{{K}_{2}}{{\alpha }_{21}},{K}_{2} < \frac{{K}_{1}}{{\alpha }_{12}}\)
-
\({K}_{1} > \frac{{K}_{2}}{{\alpha }_{21}},{K}_{2} > \frac{{K}_{1}}{{\alpha }_{12}}\)
where K i is the carrying capacity of species i and α ij represents the effect of species j on i where i, j = 1, 2 and i ≠ j.
References
Gause, G. F. Experimental studies on the struggle for existence. J. Exp. Biol. 9, 389–402 (1932).
Grover, J. P. Resource Competition (Springer, New York, 1997).
MacArthur, R. & Levins, R. The limiting similarity, convergence, and divergence of coexisting species. Am. Nat. 101, 377–385 (1967).
Hardin, G. The competitive exclusion principle. Science 131, 1292–1297 (1960).
Tubay, J. M. et al. The paradox of enrichment in phytoplankton by induced competitive interactions. Sci. Rep. 3, 2835 (2013).
Kakishima, S. et al. The contribution of seed dispersers to tree species diversity in tropical rainforests. R. Soc. Open. Sci. 2, 150330 (2015).
Ekaka-a, E. N. & Nafo, N. M. Stability analysis of predator-prey interaction with a crowding effect. Scientia Africana 12, 23–27 (2013).
Hartley, S. & Shorrocks, B. A general framework for the aggregation model of coexistence. J Anim Ecol 71, 651–662 (2002).
Sevenster, J. G. Aggregation and coexistence 1. Theory and analysis. J. Anim. Ecol. 65, 297–307 (1996).
Shorrocks, B., Rosewell, J., Edward, K. & Atkinson, W. Interspecific competition is not a major organizing force in many insect communities. Nature 310, 310–312 (1984).
Hara, T., Koike, F. & Matsui, K. Crowding effect in marine macrophytic algae population. Bot. Mag. Tokyo 99, 319–321 (1986).
Suehiro, K., Ogawa, H. & Hozumi, K. The crowding effect in an artificially mixed population of three species. Bot. Mag. Tokyo 98, 347–357 (1985).
Ikeshoji, T. Self-limiting ecomones in the population of insects and some aquatic animals. J. Pesticide Sci. 2, 77–89 (1977).
Ghazal, A. M. & Avery, R. A. Population dynamics of Hymenolepis nana in mice: fecundity and the ‘crowding effect’. Parasitology 69, 403–415 (1974).
Iwao, S. Analysis of contagiousness in the action of mortality factors on the western tent caterpillar population by using the m*-m relationship. Res. Popul. Ecol. 12, 100–110 (1970).
Lloyd, M. ‘Mean crowding’. J. Anim. Ecol. 36, 1–30 (1967).
Tanaka, Y., Yoshimura, J., Simon, C., Cooley, J. R. & Tainaka, K. Allee effect in the selection for prime-numbered cycles in periodical cicadas. PNAS 106, 8975–8979 (2009).
Schreiber, S. J. & Kelton, M. Sink habitats can alter ecological outcomes for competing species. J. Anim. Ecol. 74, 995–1004 (2005).
Levin, D. A. Reinforcement of reproductive isolation: Plants versus animals. Am. Nat. 104, 571–581 (1970).
Yoshimura, J. & Clark, C. W. Population dynamics of sexual and resource competition. Theoret. Pop. Biol. 45, 121–131 (1994).
Taylor, A. & Crizer, A. A Modified Lotka-Volterra competition model with a non-linear relationship between species. Rose-Hulman UMJ 6, 1–14 (2005).
Anaconda, Inc. Anaconda Python 3. 6. Available at https://www.anaconda.com/download/#windows (Downloaded: June 15, 2017) (2016).
Acknowledgements
This work was partly supported by grants-in-aid from the Japan Society for Promotion of Science (nos 22255004, 22370010, 26257405 and 15H04420 to JY; no. 26400388 to SM; nos 14J02983, 16H07075, 17J06741 and 17H04731 to HI; nos 25257406 and 16H04839 to TTo); and the Mitsubishi Scholarship (MISTU1722) to MKAG.
Author information
Authors and Affiliations
Contributions
M.K.A.G., T.T.a., T.N. and J.Y. conceived the study. M.K.A.G., T.T.a., T.N. and J.Y. built and analyzed the model. M.K.A.G. and T.T.a. built a program and ran the numerical simulations. H.I., S.M., G.I. and T.O. verified the mathematical properties of the models. H.I., T.T.o. and J.Y. developed biological interpretations of the model. M.K.A.G., T.T.a. and J.Y. wrote the manuscript. M.K.A.G. and T.T.a. as the lead authors. All authors reviewed the manuscript and gave final approval for publication.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gavina, M.K.A., Tahara, T., Tainaka, Ki. et al. Multi-species coexistence in Lotka-Volterra competitive systems with crowding effects. Sci Rep 8, 1198 (2018). https://doi.org/10.1038/s41598-017-19044-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-19044-9
This article is cited by
-
Lotka–Volterra model with Allee effect: equilibria, coexistence and size scaling of maximum and minimum abundance
Journal of Mathematical Biology (2023)
-
Evolvability and Macroevolution: Overview and Synthesis
Evolutionary Biology (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
