
Abstract
Plants retain rhythmic physiological responses when adapting to environmental challenges. However, possible integrations between drought conditions and those responses have not received much focus, especially regarding crop plants, and the relationship between abiotic stress and the diurnal cycle is generally not considered. Therefore, we conducted a genome-wide analysis to identify genes showing both diurnal regulation and water-deficiency response in rice (Oryza sativa). Among the 712 drought-responsive genes primary identified, 56.6% are diurnally expressed while 47.6% of the 761 that are down-regulated by drought are also diurnal. Using the β-glucuronidase reporter system and qRT-PCR analyses, we validated expression patterns of two candidate genes, thereby supporting the reliability of our transcriptome data. MapMan analysis indicated that diurnal genes up-regulated by drought are closely associated with the starch-sucrose pathway while those that are down-regulated are involved in photosynthesis. We then confirmed that starch-sucrose contents and chlorophyll fluorescence are altered in a diurnal manner under drought stress, suggesting these metabolic diurnal alterations as a novel indicator to evaluate the drought response in rice leaves. We constructed a functional gene network associated with the starch-sucrose KEGG metabolic pathway for further functional studies, and also developed a regulatory pathway model that includes OsbZIP23 transcription factor.
Similar content being viewed by others


Introduction
Plants, animals, and microorganism experience diurnal fluctuations in light and temperature that follow a 24-h cycle with diverse biological activities and physiological outputs1,2,3. This rhythmic pattern influences physiological behaviors including chronotype, diseases such as obesity and diabetes, the application of drug therapy, and cognitive functions in humans2, 4. Also, the diurnal rhythmicity of intestinal microbiota controls the transcription and epigenetic regulation of hosts5. In plants, it affects transcriptional regulation, hormone levels, stomatal conductance, photosynthetic carbon metabolism, nitrate and potassium uptake, metabolism, and overall growth and development1, 6,7,8,9,10,11. For example, bulk stomatal conductance is higher mid-day in the leaves of rice (Oryza sativa) and lower in the morning and evening due to variations in leaf chlorophyll concentration and solar radiation9. Research with Zea mays has demonstrated that the efficiency of diurnally varied uptake for nitrate, potassium, and phosphate differs among genotypes10. Promoter analyses have revealed that each phase in the regulatory module of ME/G-box, EE/GATA, and PBX/TBX/SBX controls transcriptional circuity12, 13. Rice is a major model system for which studies of functional genomics have made great advances. In fact, more than 50,000 genome-wide transcriptome data, 3000 whole-genome re-sequencing data, and 8646 QTLs have been obtained for those plants; its ease of transformation means that a gene-indexed mutant population is now available for almost half of this genome14,15,16,17,18,19,20,21. Other useful resources include datasets (GSE36040; GEO, http://www.ncbi.nlm.nih.gov/geo/) that cover diurnal rhythms under natural field conditions, with data collected from leaf blades at 2-h intervals over 2 d during each of nine developmental stages22.
Plants must also contend with abiotic stresses, such as water scarcities, that threaten global agriculture and food security due to a growing world population and climate change23. Crop productivity that solely depends upon rainfall is projected to decline as much as 50% by 2020, and yields in some countries might be reduced 90% by 210024. Drought events now affect 20% of the land used for rice cultivation, i.e., 19 to 23 million ha (IRRI, http://www.irri.org). Immense economic losses related to drought mean that research of stress-tolerant crops is a major issue. The functions of 105 drought stress related genes have been characterized in rice, and are now part of the Overview of functionally characterized Genes in Rice Online (OGRO) database25. Although some investigations have focused on elucidating the mechanisms of drought tolerance, functions have been identified for only 3% of the related genes in the entire rice genome26.
Researchers have proposed that crosstalk exists in the relationship between abiotic stress and the diurnal cycle. In Arabidopsis thaliana, ABA signaling plays a role in regulating numerous genes that are drought-responsive in later daylight hours27. Other studies have also indicated a reciprocal relationship exists between the Salt Overly Sensitive (SOS) pathway and diurnal rhythms in rice, based on diurnal patterns of expression for OsSOS1, OsSOS2, OsSOS3 and RD29A transcripts28, 29. In submerged rice plants, root extension and internal O2 concentrations in the roots exhibit relative fluctuations in their diurnal cycles when compared with plants that are not submerged. Ethanol production contributes to the survival of roots when those plants are submerged during the nighttime30. Moreover, approximately 75% more cold-responsive transcription factors (TFs) in Arabidopsis are induced in the morning than at night, and the diurnal cycle operates at a lower amplitude when temperatures are colder than normal31. That cycle also drives the highest cold-inducible Ca2+ signaling in guard cells and in whole plants in the middle of a photoperiod. The circadian clock is also engaged in whole-plant signaling32. Finally, rice Cold induced MYB 1 (CMYB1) is thought to induce cold-responsive and rhythmic gene expression under nighttime temperatures33.
Despite those research advances, however, little has been done to improve our understanding of the relationship between diurnal rhythm and drought stress, and only two reports have been made about genome-wide analyses that examined the link between those components in Arabidopsis and in Populus 27, 34. Because abiotic stress is strongly affected by changes in the light/dark cycle and the temperature cycle, it is essential that diurnal rhythms should be included in such investigations of mutual interactions. Therefore, our objective here was to develop an effective means for studying how drought stress influences crosstalk with the diurnal cycle in rice. In particular, we examined key regulators of the drought-stress mechanism, and included “time of day” as one of our controlling factors. Global transcriptome analyses of genes involved in the drought response and in diurnal rhythms were combined. This allowed us to identify 766 genes that were then characterized through Gene Ontology enrichment and MapMan analyses. Combining our understanding of the predicted protein–protein interaction network with that of the starch-sucrose KEGG metabolic pathway provides a useful framework for further functional studies to elucidate the crosstalk between diurnal rhythm and the drought response. We obtained transcriptome data from drought-tolerant transgenic plants that over-express the OsbZIP23 transcription factor. By incorporating those data, we developed a regulatory pathway model that includes OsbZIP23 to explain this crosstalk.
Results
Genome-wide identification of rice genes showing both a drought response and diurnal regulation
To determine which of our genome-wide candidate genes are drought-responsive, we used a meta-expression database for abiotic stress based on nine series of Affymetrix array data that comprised 131 samples (Fig. 1). Those data were downloaded from the National Center for Biotechnology Information (NCBI) Gene Expression Omnibus (GEO; http://www.ncbi.nlm.nih.gov/geo/) and were log2-normalized35. We then generated log2 fold-change data (expression in the stressed sample versus the MOCK control) for each replicate (Table S1). From this we identified 712 genes that were up-regulated under drought and 761 that were down-regulated, based on K-means clustering (KMC) in MultiExperiment Viewer (MeV) software36.

Identification and heatmap analysis of genes associated with both diurnal rhythm and drought response. Analysis (via KMC) of expression profiles for 766 genes with diurnal rhythm that are either up-regulated (403) or down-regulated (363) by drought. Average values for log 2 fold-changes under drought, cold, heat, or submergence conditions are shown in heatmap on left side. Drought-responsive genes are marked with yellow boxes. In two heatmaps on right side, log2-normalized intensity values indicate expression patterns for genes (red and green boxes) that were monitored at 2-h intervals (over 2-d period) using leaf blades from field-grown rice plants sampled during 9 developmental stages.
Diurnal patterns for these differentially expressed gene groups were further analyzed by using RXP_0002 data sets from the RiceXPro (RXP) website that were deposited as GSE36040 in the NCBI GEO. These data had been collected from the uppermost fully expanded leaf blades in the main stem of natural field grown rice plants at 2-h intervals for 2 d during nine developmental stages including the vegetative, reproductive, and ripening stages in 200822. These data are helpful to observe diurnal rhythmic expression patterns according to the nature environment through entire growth stages. Analysis of KMC identified 403 genes showing diurnal rhythm plus upregulation under drought, while 363 genes showed diurnal rhythm with downregulation under drought (Tables S2 and S3). In detail, 34.99% of 403 genes exhibit the highest expression in the predawn; 29.53% of these show the midday peaks; 18.61% of these express the highest in the midnight; and 16.87% of these show the expression peak in the late day. The proportion of genes showing the expression peak in the predawn is also the highest for the latter 363 genes as 38.29%. The second largest percentage is 24.52% with regards to the midday time. The proportions of genes highly expressed during the midnight and the late day are 20.66% and 16.53%, respectively.
Among the remaining up-regulated genes, 35 were more highly expressed during the vegetative growth stage than during the reproductive growth stage, 16 were more highly expressed during the reproductive growth stage, and 258 had very low levels of expression or less-obvious diurnal patterns (Fig. 1). Among the remaining down-regulated genes, 110 showed strong expression during the vegetative growth stage but significantly reduced expression during the reproductive stage, 43 were more highly expressed during the reproductive growth stage than at any other stage, and 210 had less-clear expression patterns for diurnal rhythm. These data indicated that 56.6% (403/712) of the genes up-regulated by drought stress were also regulated by diurnal rhythm, while 47.7% (363/761) of the genes down-regulated by drought were also diurnally controlled. Thus, both groups of genes are a useful source for studying crosstalk between drought stress and diurnal regulation.
Analysis of candidate genes with known functions using literature searches emphasizes their significance in promoting or maintaining crop yields when plants are subjected to abiotic stresses
To search for some of our candidate genes that have already been characterized through genetic and molecular studies, we utilized the OGRO database, which contains a good summary of rice genes with known functions25. There, we found that 53 of those candidates were functionally characterized previously (Table 1). Among them, 27 appeared to be involved in stress responses, including 11 that conferred drought tolerance. Four genes had roles in starch or sucrose biosynthesis, or in determining starch contents, while another eight had functions in grain yield. This demonstrated that genes showing both diurnal and water-stress responses might play key roles in promoting or maintaining crop yields in a world that is undergoing abiotic stresses.
Validation of expression patterns for drought-inducible and diurnally rhythmic genes, using the GUS reporter system
To confirm the expression patterns of candidate genes up-regulated by drought, we used two strategies: screening of promoter trap lines (as we have described previously) and generation of transgenic plants carrying the β-glucuronidase (GUS) reporter gene vector under the control of the target gene promoter (Fig. 2)37, 38. For the former approach, we analyzed promoter trap lines for 70 candidate genes and found that only one had positive GUS expression that matched the activity of its co-segregating genotypes. PFG_3A-02918 is a promoter trap line of LOC_Os05g07810, which encodes a universal stress protein domain-containing protein. The T-DNA in this line is located in the first intron (Figure S1). Co-segregation tests of GUS activity and the T-DNA insertion confirmed that GUS expression was caused by the promoter trap of the target gene (Figure S1). In particular, GUS activity was observed in the shoots of 10-day-old seedlings that were subjected to drought stress for 6 h (Fig. 2a). Quantitative RT-PCR analysis revealed that LOC_Os05g07810 expression was approximately nine times in the stressed plants than in the MOCK. Meta-analysis also showed that this gene followed a diurnal pattern, with expression being highest at the end of the night and lowest at the end of the day (Fig. 2a).

Validation of differential expression patterns under drought for two candidate genes using GUS reporter system and qRT-PCR. Results of qRT-PCR analyses: (a) drought-inducible GUS expression of LOC-Os05g07810 by promoter trap system, and its expression level under drought stress and diurnal rhythm relative to OsUbi5; and (b) drought-inducible GUS expression of LOC_Os10g42610 by promoter-GUS vector system, and its expression level under drought stress and diurnal rhythm relative to OsUbi5. D0, untreated leaf sample; M3, MOCK (Control) monitored for 3 h after drought stress; M6, MOCK after 6 h; M12, MOCK after 12 h; D3, leaf sample under drought for 3 h; D6, leaf sample under drought for 6 h; D12, leaf sample under drought for 12 h. For analyses of diurnal expression: EN, leaf sample collected at end of night; Mid-D, mid-day; ED, end of day; and Mid-N, middle of night. OsUbi5 was used as internal control for qRT-PCR analysis. Error bars indicate standard errors of 3 replicates. *, **, ***, and ****: values are significantly different from each other at P ≤ 0.05, P ≤ 0.01, P ≤ 0.001, and P ≤ 0.0001, respectively.
For the latter approach, we constructed promoter-GUS reporter gene vectors under the control of the LOC_Os10g42160 promoter. Using T1 transgenic plants that expressed GUS, we tested inducible patterns under drought and found that, as we expected, GUS reporter activity was significantly stimulated by 6 h of stress (Fig. 2b). We also conducted quantitative RT-PCR analyses and confirmed that LOC_Os10g42610 expression was approximately three times higher in stressed plants than in the MOCK control. Expression was again strongest at the end of the night and lowest at the end of the day (Fig. 2b).
The expression patterns of additional drought-inducible and diurnally rhythmic candidate genes encoding glycogen branching enzyme (OsSBE3, LOC_Os02g32660), Osβ-amylase3 (OsBAM3, LOC_Os03g04770), Osβ-amylase2 (OsBAM2, LOC_Os10g32810), Osβ-amylase5 (OsBAM5, LOC_Os10g41550), neutral/alkaline invertase (LOC_Os04g33490), and sucrose synthase (LOC_Os03g22120) were also validated (Figures S4 and S5). As expected, they all exhibited diurnal rhythmic pattern and drought induced expression. These expression data further enhance the fidelity of the global transcriptome data used in this study.
Gene Ontology enrichment analysis reveals the significance of water-stress responses associated with diurnal regulation
We performed Gene Ontology (GO) enrichment analysis with the Rice Oligonucleotide Array Database (ROAD; www.ricearray.org/index.shtml) to identify the biological roles of candidate genes that are both drought-responsive and have a diurnal rhythm of expression. The GO terms were evaluated according to two thresholds (≥two-fold enrichment and P-value < 0.05)39. In all, 19 terms were significantly enriched for 403 genes. Among them, 15 were related to drought stress or diurnal rhythm (Fig. 3a, Table S4). Several of those terms could be organized into categories for carbohydrate metabolism, transcription and translation, transport, protein process, and auxin pathway. Highly overrepresented terms, such as those for embryo development (GO:0009790; by 55.8-fold), response to water (GO:0009415; by 20.6), starch biosynthetic process (GO:0019252; 12.4), polysaccharide catabolic process (GO:0000272; 10.9), glycogen biosynthetic process (GO:0005978; 9.3), and carbohydrate metabolic process (GO:0005975; 2.3) indicated that these biological processes were more likely associated with up-regulated drought responses and diurnal rhythms.

GO enrichment analysis of drought-responsive genes showing diurnal rhythm. Within category of Biological Process, GO enrichment terms are identified for 766 diurnally controlled genes that are either up-regulated (403; a) or down-regulated (363; b) by drought. Fold-enrichment values are shown along Y-axis; enriched GO terms, along X-axis.
In total, 20 GO terms were enriched for 363 genes that were down-regulated by drought and also expressed diurnally. These terms covered categories for organ development, light reaction, carbohydrate metabolism, and transport (Fig. 3b, Table S5). Leaf development (GO:00048366; by 39.2-fold), flower development (GO:0009908; by 10.3), chlorophyll biosynthetic process (GO:0015995; 14.0), photosynthesis light harvesting (GO:0009765; 11.5), glycine catabolic process (GO:0006546; 28.0), cellular carbohydrate metabolic process (GO:0044262; 10.9), malate metabolic process (GO:0006108; 8.5), extracellular polysaccharide biosynthetic process (GO:0045226; 8.5), and tricarboxylic acid cycle (GO:0006099; 5.9) appeared to be important participants in the potential crosstalk between the diurnal cycle and drought-responsive downregulation.
MapMan analysis reveals significant pathways associated with diurnal regulation and the drought response
To identify the metabolic pathways associated with drought and diurnal regulation, we conducted a MapMan Metabolism overview analysis for 766 candidate genes differentially expressed under drought stress as well as involved in diurnal rhythm. The Metabolism overview, installed in the MapMan toolkit (3.51R2), was executed as we have previously described (Fig. 4a, Table S6)40. From this we determined that pathways for starch biosynthesis and degradation, the sucrose degradation pathway, and lipid degradation metabolism were positively associated with drought and diurnal regulation. In contrast, light reactions and the Calvin Cycle in photosynthesis, pathways for ascorbate and glutathione, and the pathway for synthesis of cell wall precursors were negatively associated with this proposed crosstalk. We were also interested to find a strong positive association with the starch synthesis and degradation pathways. To get more detailed information about these pathways, we analyzed the Sucrose-Starch overview (Fig. 4b) and Plant Glycolysis overview (Fig. 4c) installed in the MapMan toolkit and identified detailed elements in the starch synthesis and degradation pathways that are associated with this crosstalk. Subsequently, we found up-regulated genes that encode ADP-glucose pyrophosphorylase (AGPase), starch synthase, and starch branching enzyme in the starch synthesis pathway, as well as upregulated genes for β-amlyases and disproportionation enzyme in the starch degradation pathway. Moreover, details about light reactions and the Calvin Cycle were identified in the Photosynthesis overview (Fig. 4d) and the Plant Glycolysis overview (Fig. 4c). We also discovered down-regulated genes, such as those for the light harvesting complex of Photosystem II (PSII) chlorophyll binding protein 1 (Lhcb1), chlororespiratory reduction 23 (CRR23), and proton gradient regulation 5 in photosystem light reactions, as well as glyceraldehyde 3-phosphate dehydrogenase A subunit (GAPA) and fructose-1,6-biophosphate in the Calvin Cycle. Based on these results, we speculated that the starch-sucrose pathway and photosynthesis are critical components of this crosstalk between drought stress and diurnal rhythm. Additional analyses covered overviews for regulation, transcription, large enzymes, cellular-responsive, secondary metabolism, receptor-like kinases, and proteasomes (Figures S7 and S8). Genes up-regulated by drought are marked in red while down-regulated genes are marked with green.

MapMan analysis of drought-responsive genes showing diurnal rhythm. Results of mapping 766 genes to Metabolism overview (a), Sucrose-Starch overview (b), Plant Glycolysis overview (c), and Photosynthesis overview (d). Red boxes, genes up-regulated under drought; green boxes, genes down-regulated under drought. Within Metabolism overview, red and green lines indicate MapMan functional groups selected for physiological validation.
Fluctuations in levels of soluble sugar and starch contents in rice leaves under drought stress during the day/night cycle
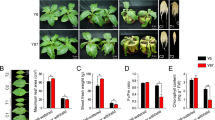
Rice plants were grown for five weeks in a controlled-climate chamber (28 °C/20 °C, 66% relative humidity, and 12-h photoperiod from incandescent lamps). Some of these plants were then exposed to drought stress for 9 d while the others remained under standard growing conditions (MOCK control). Leaves were sampled at the end of each dark period (night) and the end of each light period (day), and their soluble sugar and starch contents were measured by an enzymatic method, as described previously41. Under drought treatment, diurnal patterns of starch and soluble sugars, including sucrose, were altered, with soluble sugar contents tending to increase and starch levels decreasing in response to stress (Fig. 5)42. The soluble sugar contents peaked during the daytime43 and, in fact, were elevated even further as a result of drought treatment. By comparison, starch levels remained relatively low even at the end of the day, suggesting rapid turnover. These findings suggested that changes in sugar and starch contents could serve as a new indicator for evaluating the drought response associated with diurnal rhythm in rice leaves. When combined with the results from our genome-wide analysis presented above, the starch degradation pathway appears to have a particularly important role in altering the synthesis of those metabolites.

Diurnal alterations in soluble sugars and starch contents from rice leaves under drought stress. On Y axis: contents (mg g−1 FW of sample) of a) glucose; (b) fructose; (c) sucrose; and (d) starch. X-axis features time of sampling: EN, end of night; and ED, end of day. MOCK control, samples not subjected to stress treatment; Drought, samples from plants exposed to drought conditions. Error bars indicate standard errors of 3 replicates.
Changes in chlorophyll fluorescence in rice leaves under drought stress during the day/night cycle
Chlorophyll fluorescence (Fv/Fm)44 was measured mid-day and mid-night in 2-month-old rice plants that had been exposed to drought treatment for 0 to 5 d (Fig. 6). Although fluorescence is normally much lower mid-day than mid-night, the degree of difference is enlarged under a water deficiency45,46,47,48. We also found that mid-day values were drastically lower after 4 d of drought. When combined with the data gained in our analysis above, these findings led us to speculate that PSII in light reactions has a crucial role in this diurnal drought response, probably due to the water supply being limited.

Diurnal alterations in chlorophyll fluorescence and total chlorophyll content in rice leaves under drought stress. Chlorophyll fluorescence (Fv/Fm) was measured in leaf samples from non-stressed MOCK plants (0) and drought-stressed plants (2, 4, and 5), collected mid-day (Mid-D) and mid-night (Mid-N) for periods indicated: 2 Mid-D and 2 Mid-N, sampled for 2 d; 4 Mid-D and Mid-N, sampled for 4 d; and 5 Mid-D and 5 Mid-N, sampled for 5 d. Error bars indicate standard errors of 3 replicates. *, **, ***, and ****: values are significantly different from each other at P ≤ 0.05, P ≤ 0.01, P ≤ 0.001, and P ≤ 0.0001, respectively.
Construction of a functional interaction network focused on genes in the starch biosynthesis and degradation pathways for which expression shows both a drought response and diurnal rhythm
Because the starch biosynthesis and degradation pathways appear to be closely linked with the drought response and diurnal control, we explored the mechanism that might regulate this. To do so, we developed a functional interaction network mediated by 276 genes that encode the components of the entire starch biosynthesis and degradation pathways (Fig. 7, Table S7). These were derived from the RiceCyc Interaction Viewer (http://pathway.gramene.org/gramene/ricecyc.shtml) and the Rice Interaction Viewer (http://bar.utoronto.ca/interactions/cgi-bin/riceinteractionsviewer.cgi)49, 50. From this, we identified 69 interactions associated with six genes for starch biosynthesis and five for starch degradation. Within the network, 11 genes (nodes with bold-red borders in Fig. 7) were up-regulated by drought and also diurnally rhythmic. Six other genes had been functionally characterized previously51,52,53,54,55,56. Therefore, the interactions between them and genes not yet characterized might help us study the latter more closely. Through MapMan analysis of network components, we were able to assign functional roles for 15 genes associated with carbohydrate metabolism; eight for RNA processing, three for abiotic stress; four for transport metabolism; and 22 for protein degradation, folding, modification, or processing.

Analysis of functional gene network associated with pathways for starch biosynthesis and starch degradation. We used RiceCyc, Rice Interaction Viewer, and Cytoscape program to develop network, based on information derived from MapMan analysis. Nodes with bold-red borders, genes that are both diurnally rhythmic and up-regulated by drought; rounded rectangles, genes for which functions were reported previously. Colored nodes (circles) represent functional classification information from MapMan analysis (see Fig. 4), with color of edge indicating differential Pearson’s correlation coefficient (PCC) values that were calculated using anatomical microarray data from ROAD, and which ranged >0.5 (red). Thickness of edge was determined based on Interlog confidence value from Rice Interaction Viewer, with thicker edges indicating higher confidence. Dotted arrows with circle heads show genes encoding proteins bound by OsbZIP23; dotted lines with triangle heads, genes targeted by OsbZIP23. Blue-dotted lines, genes regulated negatively in overexpression of OsbZIP23; red-dotted lines, genes regulated positively in overexpression of OsbZIP23. Color indicates assigned function: yellow, carbohydrate metabolism; green, RNA processing; orange, abiotic-stress response; pink, protein degradation/folding/modification/processing; blue, transport; and white, other functions.
Among the genes assigned as TFs in the Transcription overview in MapMan analysis (Fig. 3b), three have known functions in drought tolerance: Oryza sativa basic leucine zipper 23 (OsbZIP23, LOC_Os02g52780), HD-Zip I homeobox 22 (HOX22, LOC_Os04g45810), and ABA responsive element binding factor 1 (ABF1, LOC_Os01g64730)57,58,59. Because we could utilize only earlier results from CHIP-seq and RNA-seq analyses of OsbZIP23 TF, we integrated only genes for drought tolerance that are either regulated by or bound by the bZIP23 TF, according to this network57. In all, 11 genes appeared to be bound by OsbZIP23, with one being a direct target. Four genes were up-regulated and three were down-regulated by overexpression of OsbZIP23 in the starch-sucrose metabolic pathway under drought conditions. Among them, five were diurnal and up-regulated by drought, similar to the pattern for OsbZIP23. These genes might be valuable for elucidating the roles of OsbZIP23 when linking the drought mechanism to starch-sucrose metabolism.
Discussion
Plants challenged by drought must respond if they are to survive and continue to grow. Before deeper functional analyses can be conducted, researchers must first understand the possible connections between water stress and the diurnal cycle. This can be accomplished by using meta-expression data to identify differentially expressed genes28. Results from previous genome-wide transcriptome analysis have helped us discover the entire molecular network and examine stress-target genes and their complex responses in plants60,61,62. Here, we used microarray data to determine that 56.6% (403) of our 712 candidate drought-responsive genes are also diurnally expressed, while 47.7% (363) of the 761 genes down-regulated by drought are also diurnal. Hence, these 766 candidate genes provide fundamental materials for investigations of the reciprocal network between drought stress and diurnal rhythm. Because a large number of these drought-responsive genes are also diurnally rhythmic, we must consider what role the diurnal cycle has in that drought response. Probe sets for leaves from rice seedlings grown under normal conditions have indicated that only 10.96% of the rice transcriptome shows diurnal oscillations6. Because the percentage of rice genes that are both diurnally regulated and drought-responsive is higher than those that fulfill only one of those criteria, the extent to which drought stress influences these transcriptome rearrangements appears to be dependent upon the time of day. Similar results have been found with Arabidopsis 27. Regarding time phasing patterns, it is clearly seen that more than one third of drought responsive genes relatively show higher expression level during predawn. The phase distribution of diurnal rhythm might not be influenced by drought stress because the period from 2:00 am to 4:00 am has the highest proportion of probe sets showing diurnal rhythmic distribution in rice seedling leaves under normal conditions6.
As we have described for previous investigations, we identified meaningful biological processes and visualized significant metabolic pathways through GO enrichment and MapMan analyses26, 63. Results from the former suggested that biological processes mainly related to carbohydrate metabolism are important in the crosstalk between the drought up-regulation and diurnal rhythm. Other researchers have also indicated that carbohydrates are closely associated with water-deficit responses and the diurnal cycle. For example, levels of trehalose, a carbohydrate storage molecule, are elevated through manipulation of the intermediate trehalose-6-phosphate, which enables transgenic rice plants to become more drought-tolerant through sugar-signaling and carbohydrate metabolism64. However, only a few examinations have considered the strong association between drought stress and diurnal rhythm, as coupled with carbohydrate metabolism10, 43, 65. We also found that the GO term for embryo development had the highest fold-enrichment in genes that were up-regulated by both drought and diurnal rhythm. This connection might be explained by the processes of physiological desiccation and dormancy in rice that occur during embryo development prior to grain harvest66. In barley, the diurnal cycle has a critical role in the formation of caryopses67. The GO term with the second-highest fold-enrichment was related to the water-stress response.
Terms for light reactions and relative carbohydrate metabolism were also significantly represented in our analysis of genes that are both drought down-responsive and diurnally rhythmic. This is manifested in C3 plants by a reduction in CO2 diffusion through the stomata and mesophyll under drought conditions68,69,70. In Phaseolus vulgaris, the speed and the extent of photosynthetic recovery depends upon variations in the carbon balance68, 71, 72. Because 15% of relatively down-regulated genes in drought-tolerant Thellungiella are photosynthetic, it is highly probable that those genes also have key roles in light reactions73. However, future studies are required to determine whether these features are conserved in rice, and to learn how they might be connected to the drought mechanism via the diurnal cycle. In Ricinus communis, leaf development is influenced by the combination of drought stress and diurnal regulation70. Although those tissues grow more rapidly late at night than in the late afternoon, their diurnal amplitude in the rate of leaf growth decreases during periods of stress74.
Our MapMan results confirmed the findings from GO analysis that terms for the sucrose-starch pathway and light reactions are highly enriched. There, 10 genes allocated to the sucrose-starch pathway were up-regulated by drought while four genes related to photosynthesis were down-regulated (Fig. 5). Diurnal fluctuations in starch and sucrose contents under a water deficit are possibly influenced by the regulation of genes allocated to that pathway. Starch is accumulated during the daytime as a product of photosynthesis, but is then degraded at night to provide and export sugars to sink organs43, 75. Soluble sugars are accumulated in leaves when the water supply is limited in the growing environment. They can act as osmolytes to maintain normal transpiration and leaf-water content under drought conditions42. However, in periods of drought, starch is degraded to soluble sugars during daylight hours as well. Our findings here are supported by previous work with Arabidopsis thaliana, where the accumulation of sucrose at the end of the day is critical in protecting plants against xenobiotic and oxidative stresses76. We also speculated that the alteration of chlorophyll fluorescence under a water deficit is related to four genes allocated to photosynthesis (Fig. 6), possibly affecting PSII. Because photosynthesis is influenced by drought due to limitations in carbon uptake and CO2 diffusion, we might expect diverse metabolic changes to occur accordingly47. With regard to the diurnal cycle, photo-inhibition by strong light intensities is more like to happen mid-day, which then causes the photosynthesis potentiality in PSII to decline45,46,47. We might conclude that starch degradation in the daytime increases the level of soluble sugars, which then can compensate for drought-induced changes in photosynthate contents. Therefore, future experiments should also include the monitoring of changes in metabolites during daylight hours.
For our interaction network, we incorporated genes that are up-regulated by drought as well as those with diurnal patterns of expression. Several candidates showed the most potential for functional analysis, including OsBAM3, and OsIsoamylase1 (LOC_Os08g40930). These were selected because one homolog from Arabidopsis, AtBAM1 (AT3G23920), is down-regulated under drought stress and its expression is linked with diurnal degradation of starch to sustain proline biosynthesis during periods of osmotic stress. Furthermore, the atbam1 mutant is rendered drought-tolerant because starch does not break down in its guard cells and its rate of stomatal opening is lower when compared with normal plants77, 78. Rice granule-bound starch synthase1 (LOC_Os06g04220), which functions similarly to OsIsoamylase1 in the pathway for starch biosynthesis, has decreased expression in the ABA-independent pathway under a water deficit79. Based on our findings here, we propose that starch synthesis is greater under drought conditions due to the upregulation of ADP-glucose pyrophosphorylase large subunit 1 (OsAGPL1, LOC_Os05g50380), starch synthase I (OsSSI, LOC_Os06g06560), and OsSBE3. Consequently, higher levels of sucrose and soluble sugars might be turned over from enhanced starch contents through the expression of four candidate β-amylase genes (OsBAM2, OsBAM3, OsBAM5, and LOC_Os03g22790), plus an α-amylase precursor (LOC_Os06g49970), and 4-α-glucanotransferase (LOC_Os07g43390) (Fig. 4). The expression of OsBAM2, OsBAM3, and OsBAM5 were strongly induced in the end of night (Figures S4 and S5). In previous studies, expression of AtBAM1 was the highest during the shifting period from dark to light80. However, the expression phase of AtBAM1 is shifted to lateday in drought conditions27. Similarly, the expression of β-amylase genes in rice might have the shift of expression phase to daylight hours to produce increased amount of sucrose and soluble sugars in water deficient response. How the transcriptome of drought responsive genes in rice shifts according to diurnal pattern should be elucidated through further analyses.
OsbZIP23 could be responsible for regulating interactions between drought stress and diurnal rhythm. This TF contributes to drought tolerance, and it binds, targets, and regulates several genes encoding enzymes in the starch-sucrose pathway57, 81. In particular, drought-induced ABA inhibits OsPP2C49, and SAPK2 is then activated to phosphorylate OsbZIP2357, 82. Hence, the expression of drought-responsive genes is triggered by this activation of OsbZIP2357. Overexpression of OsbZIP23 in rice leads to a drought-tolerant phenotype and increased soluble sugar contents in the leaves of stressed plants83. The four β-amylases mentioned above are bound by OsbZIP23. Their function in breaking down starch might play a role in the drought response and could explain the elevated production of soluble sugars84. OsAGPL1 is also bound by OsbZIP23, and activation of the latter causes the former to be up-regulated. Thus, transitory starch synthesis in the leaves might be driven by OsAGPL1. A drought-related decline in chlorophyll fluorescence might be eased by the overexpression of OsbZIP23 under water stress81. Lhcb1 (LOC_Os01g41710) is bound by OsbZIP23 when water is limiting, and LOC_Os05g28090, which encodes chloroplast NAD(P)H dehydrogenase, is also regulated by OsbZIP23. Therefore, the downregulation of both of these genes, as controlled by OsbZIP23, might induce the photosynthetic and diurnal physiological differences in response to drought.
In conclusion, we exploited the advantages of meta-expression analysis to discover a functional gene network that explains the cross-talk between diurnal rhythm and drought stress. Our results indicate that the starch-sucrose pathway and light reactions might be its main targets. Under a water deficit, the daytime response is critical to the growth of rice plants because higher starch turnover, even in daylight hours, is to sustain limited production of photoassimilates during drought periods. The OsbZIP23 TF, which is involved in the drought-response mechanism, binds to four OsBAMs that are up-regulated by drought and also expressed diurnally. This TF also binds to OsAGPL1 in the starch-sucrose pathway and to the drought down-responsive and diurnal OsLHCB1 in PSII. Physiological and metabolic changes to starch/sucrose production and photosynthesis potentiality are stimulated by OsbZIP23 in response to drought stress. Based on these findings, we have developed a model to illustrate this regulatory mechanism (Fig. 8). Nevertheless, more functional and molecular investigations are required if we are to obtain deeper insight into this type of crosstalk.

Suggested model to describe how drought-inducible OsbZIP23 regulates both light reactions and starch-sucrose pathway. Red boxes, genes showing diurnal expression pattern and up-regulated by drought; green boxes, diurnal genes down-regulated by drought. Genes encoding proteins interacting with OsbZIP23 are linked with circle-head lines; those encoding proteins regulated by bZIP23 are connected with triangle-head arrows. Pathways in solid-line rectangles were previously elucidated. Pathways bordered by rectangles with dotted lines include genes proposed here as part of crosstalk between diurnal expression and drought response.
Methods
Analysis of drought-stress regulation and diurnal rhythms using publicly available microarray data
We downloaded nine raw data series containing 131 samples related to abiotic stress from the NCBI-GEO. This meta-expression database was then modified so we could focus on the effects of drought, cold, heat, and submergence. Using six dataseries (GSE6901, GSE21651, GSE24048, GSE25176, GSE26280, and E-MEXP-2401), we selected drought-responsive genes, then normalized the data using the Affy Package from R language and changed intensity values to a log2 scale35, 85 Average fold-change values, i.e., drought-stressed samples over control (MOCK) samples, were calculated. Genes that were up- or down-regulated by diverse drought conditions were sorted by KMC via MeV software. The average values computed for selected genes are imaged in the heatmap in Fig. 1 (left panel). We also conducted meta-expression analysis of genes that were either up-regulated (712) or down-regulated (761) according to a diurnal pattern. Using RXP_0002 data (GSE36040, GEO), we analyzed expression patterns of those genes in leaf blades sampled at 2-h intervals for 2 d during nine developmental stages for field-grown plants22. After the data were log2-normalized, KMC analysis was performed to delineate the genes that were differentially expressed diurnally. This produced seven gene groups, for which we produced heatmap images (MeV software) and then edited with Adobe Photoshop and Illustrator CS636. Moreover, we divided the genes to four groups according to their differential rhythmic patterns. The average of normalized values from the diurnal data of four time phases (Midday, 08:00–16:00; Lateday, 18:00–20:00; Midnight, 22:00–02:00; and Predawn, 04:00–06:00) were calculated to identify the peak of expression level.
Analysis of GUS expression
Screening of the promoter trap lines and GUS staining were performed as described previously37, 38, 86. We used a T-DNA-specific primer in the pGA2715 vector as well as gene-specific primer sequences to confirm, via PCR, the co-segregation of GUS expression and T-DNA insertion in rice Line PFG_3A-02918 (Table S8). The promoter for the GUS vector was constructed using the pGA3519 binary vector, as described previously87. For observing drought-inducible GUS activity in transgenic rice, we stained leaves from MOCK control and drought-stressed plants at the same time interval. Expression of GUS in the shoots was examined with an SZX16 microscope (Olympus, Japan). As negative controls for promoter trap systems of drought inducible expression patterns, we used T-DNA insertion in rice Line PFG_4A-04197 and PFG_5A-00191 to confirm the GUS activities controlled by the promoter trap of drought unresponsive genes. PFG 4A-04197 is a promoter trap line of LOC_Os03g08010 and PFG 5A-00191 is a promoter trap line of LOC_Os03g01910. Both genes showed ubiquitous expression and are not in our candidate gene list (Figure S3)38.
Plant materials and growing conditions
For RNA extractions and GUS assays, we germinated seeds of Oryza sativa L. ssp. japonica cv. Dongjin on Murashige and Skoog (MS) media and grew the seedlings for 10 d50. Conditions in the climate-controlled chamber included 28 °C/22 °C (day/night), a 12-h photoperiod, and light provided by incandescent lamps so we could monitor diurnal gene expression. Samples were collected at the end of the day (ED; 19:30), the middle of the night (MN; 1:30), the end of the night (EN; 7:30), and the middle of the day (MD; 13:30). Plants grown for 10 d under continuous light (28 °C) were then subjected to drought stress by withholding water for 3, 6, or 12 h while those assigned to the MOCK control continued to receive normal irrigation.
For the starch-sucrose assay, plants were grown for five weeks in a controlled-climate chamber from seeds germinated in MS media. After 9 d of exposure to drought conditions, samples were collected at the ED and EN time points. Two-month-old seedlings grown in a greenhouse were used for evaluating chlorophyll florescence. Afterward, the plants were moved to the climate-controlled chamber for acclimation before the stress treatments began. After 4 d of exposure to drought conditions, samples were collected at the MN and MD time points. At least three biological replicates were prepared and performed for each analysis.
Quantitative RT-PCR analysis
We extracted total RNA from whole seedlings and synthesized cDNA using methods introduced previously88, 89. The quality of cDNA samples was validated and evaluated with two drought-responsive marker genes (Oryza sativa dehydration-responsive element binding transcription factor 2b, or OsDREB2b, and OsbZIP23), the daytime-expressed Oryza sativa Late Elongated Hypocotyl (OsLHY), and the nighttime-expressed Heading Date 1 (HD1) (Figure S6)90,91,92,93. Quantitative real-time PCR analysis was conducted with Prime Q-Master Mix (2X, Real-time PCR with SYBR Green I) (GeNet Bio, Korea) and the Rotor-Gene 6000 (Qiagen, Hilden, Germany). The rice UBIQUITIN 5 (OsUbi5, LOC_Os01g22490) and rice UBIQUITIN 1 (OsUbi1, LOC_Os03g13170) served as a housekeeping gene for normalization, based on the comparative Ct (2−∆∆Ct) method50, 94, 95. All internal controls and gene-specific primer sequences are shown in Table S8. The P-values were calculated along a two-tailed distribution and two-sample unequal variance.
Analysis of gene ontology enrichment
The biological roles of selected genes were investigated through GO enrichment analysis of information from ROAD85. By using the GO enrichment tool installed in ROAD, GO terms were mapped on to the query genes with the detailed information. We generated the fold enrichment value by calculating the ratio of the query number value to the query expected value. Enriched GO terms that met our criteria of ≥two-fold enrichment and P ≤ 0.05 are shown in Fig. 3 and in Tables S4 and S5. Hyper P-values based on a hypergeometric distribution were provided by the GO enrichment tool installed in ROAD.
MapMan analysis
Candidate genes were analyzed through the Metabolism, Sucrose-Starch, Plant Glycolysis, and Photosynthesis overviews contained in the MapMan toolkit40 (Fig. 2). Additional analyses for regulation, transcription, large enzymes, cellular-responsive, secondary metabolism, receptor-like kinases, and proteasomes were also covered (Figures S7 and S8).
Measurements of starch and soluble sugar contents
We measured starch and soluble sugar contents by an enzymatic method41, 65. After fresh weights (FWs) were determined from each sampled leaf, they were ground with liquid nitrogen in a TissueLyser II (Qiagen, Germany). Extractions were performed with 10% perchloric acid. Following neutralization and centrifugation (13,000 rpm, 15 min), the aqueous phase was used for measuring soluble sugars while the sediment was used for starch. All values were calculated to the nearest mg g−1 FW of sample, based on the tissue weight and solution volume that was used with the enzymatic method.
Measurement of chlorophyll fluorescence
Chlorophyll fluorescence was evaluated using Handy PEA (Hansatech Instruments, England). The leaf samples were first dark-adapted for 15 min by closing the shutter plates of the leaf clips. Values were measured at a maximum light intensity of 3000 μm olm−2 s−1, and P-values were calculated based on a two-tailed distribution and two-sample unequal variance.
Analysis of interaction network
We proposed a functional gene network of 276 genes that encode enzymes for whole starch synthesis and the degradation pathway, using information from the RiceCyc and the Rice Interaction Viewers. Genes were ordered according to steps in each pathway. Any genes in the pathways that showed diurnal rhythm and drought-responsiveness but which were not part of these interaction results were added to the network as nodes (see representations in Fig. 7). We then added genes that were bound, targeted, and/or regulated under drought stress by OsbZIP23 TF (GEO; GSE81462)57. The network was edited with Cytoscape program (http://www.cytoscape.org/).
Data Availability
All data analyzed during this study are included in this published article and its supplementary information files.
References
Nozue, K. & Maloof, J. N. Diurnal regulation of plant growth. Plant Cell Environ. 29, 396–408 (2006).
Boyle, G. et al. Comparative analysis of vertebrate diurnal/circadian transcriptomes. PLoS One 12, e0169923 (2017).
Sato, T., Ida, T. & Kojima, M. Role of biological rhythms in the performance of physical activity. J. Phys. Fitness Sports Med. 6, 125–134 (2017).
Leone, M. J., Slezak, D. F., Golombek, D. & Sigman, M. Time to decide: Diurnal variations on the speed and quality of human decisions. Cognition. 158, 44–55 (2017).
Thaiss, C. A. et al. Microbiota diurnal rhythmicity programs host transcriptome oscillations. Cell 167, 1495–1510 (2016).
Xu, W. et al. Transcriptome phase distribution analysis reveals diurnal regulated biological processes and key pathways in rice flag leaves and seedling leaves. PLoS ONE 6, e17613 (2011).
Steer, B. T. Integration of photosynthetic carbon metabolism and nitrogen metabolism on a daily basis. In Photosynthesis and Plant Developement (ed. Marcelle, R., Clijsters, H. & van Poucke, M.) 309–320 (Junk, The Hague, 1979).
Pearson, C. J. & Steer, B. T. Daily changes in nitrate uptake and metabolism in Capsicum annuum. Planta 137, 107–112 (1977).
Maruyama, A. & Kuwagata, T. Diurnal and seasonal variation in bulk stomatal conductance of the rice canopy and its dependence on developmental stage. Agricult. Forest. Meteorol. 148, 1161–1173 (2008).
Pan, W., Teyker, R., Jackson, W. & Moll, R. Nutrient elements: Diurnal variation in nitrate, potassium, and phosphate uptake in maize seedlings: Considerations in screening genotypes for uptake efficiency. J. Plant Nutr. 10, 1819–1833 (1987).
Jiao, Y., Lau, O. S. & Deng, X. W. Light-regulated transcriptional networks in higher plants. Nat. Rev. Genet. 8, 217–230 (2007).
Michael, T. P. et al. Network discovery pipeline elucidates conserved time-of-day–specific cis-regulatory modules. PLoS Genet. 4, e14 (2008).
Mockler, T. C. et al. The diurnal project: Diurnal and circadian expression profiling, model-based pattern matching, and promoter analysis. Cold Spring Harb. Symp. Quant. Biol. 72, 353–363 (2007).
Eckardt, N. A. Sequencing the rice genome. Plant Cell 12, 2011–2017 (2000).
Huang, X., Lu, T. & Han, B. Resequencing rice genomes: An emerging new era of ricegenomics. Trends Genet. 29, 225–232 (2013).
Hirochika, H. et al. Rice mutant resources for gene discovery. Plant Mol. Biol. 54, 325–334 (2004).
Ni, J. et al. Gramene QTL database: Development, content and applications. Database 2009, bap005 (2009).
Barrett, T. et al. NCBI GEO: Archive for functional genomics data sets—update. Nucleic Acids Res. 41, D991–D995 (2013).
Chandran, A. K. N. & Jung, K.-H. Resources for systems biology in rice. J. Plant Biol. 57, 80–92 (2014).
Jung, K.-H., Jeon, J.-S. & An, G. Web tools for rice transcriptome analyses. J. Plant Biol. 54, 65–80 (2011).
Alexandrov, N. et al. SNP-Seek database of SNPs derived from 3000 rice genomes. Nucleic Acids Res. 43, D1023–D1027 (2015).
Sato, Y. et al. RiceXPro version 3.0: Expanding the informatics resource for rice transcriptome. Nucleic Acids Res. 41, D1206–D1213 (2013).
Steduto, P., Faurès, J.-M., Hoogeveen, J., Winpenny, J. & Burke, J. Introduction. In Coping with Water Scarcity: An Action Framework for Agriculture and Food Security (eds Lawrence, T. & Zanolli, G.) 1–4 (Food and Agriculture Organization of the United Nations, Rome, Italy, 2012).
Shanahan M., Shubert W., Scherer C. & Corcoran T. What’s at stake? In Climate Change in Africa: A Guidebook for Journalists (ed. Banda, F.) 10 (AMCEN Secretariat, 2009).
Yamamoto, E., Yonemaru, J.-I., Yamamoto, T. & Yano, M. OGRO: The Overview of functionally characterized Genes in Rice Online database. Rice 5, 1–10 (2012).
Chandran, A. K. N. et al. Functional classification of rice flanking sequence tagged genes using MapMan terms and global understanding on metabolic and regulatory pathways affected by dxr mutant having defects in light response. Rice 9, 17 (2016).
Wilkins, O., Bräutigam, K. & Campbell, M. M. Time of day shapes Arabidopsis drought transcriptomes. Plant J. 63, 715–727 (2010).
Soni, P., Kumar, G., Soda, N., Singla-Pareek, S. L. & Pareek, A. Salt overly sensitive pathway members are influenced by diurnal rhythm in rice. Plant Signal. Behav. 8, e24738 (2013).
Park, H. J., Qiang, Z., Kim, W.-Y. & Yun, D.-J. Diurnal and circadian regulation of salt tolerance in Arabidopsis. J. Plant Biol. 59, 569–578 (2016).
Waters, I. et al. Diurnal changes in radial oxygen loss and ethanol metabolism in roots of submerged and non-submerged rice seedlings. New Phytol. 113, 439–451 (1989).
Bieniawska, Z. et al. Disruption of the Arabidopsis circadian clock is responsible for extensive variation in the cold-responsive transcriptome. Plant Physiol. 147, 263–279 (2008).
Dodd, A. N. et al. Time of day modulates low-temperature Ca2+ signals in Arabidopsis. Plant J. 48, 962–973 (2006).
Duan, M. et al. CMYB1 encoding a MYB transcriptional activator is involved in abiotic stress and circadian rhythm in rice. Sci. World J. 2014, 178038 (2014).
Wilkins, O., Waldron, L., Nahal, H., Provart, N. J. & Campbell, M. M. Genotype and time of day shape the Populus drought response. Plant J. 60, 703–715 (2009).
Jeong, H. J. & Jung, K. H. Rice tissue-specific promoters and condition-dependent promoters for effective translational application. J. Integr. Plant Biol. 57, 913–924 (2015).
Howe, E. et al. MeV: MultiExperiment Viewer. In Biomedical Informatics for Cancer Research (ed. Ochs, M. S., Casagrande, J. T. & Davuluri, R. V.) 267–277 (Springer, 2010).
Jeon, J. S. et al. T-DNA insertional mutagenesis for functional genomics in rice. Plant J. 22, 561–570 (2000).
Jung, K.-H. et al. Genome-wide identification and functional analysis of genes expressed ubiquitously in rice. Mol. Plant 8, 276–289 (2015).
Jung, K.-H. et al. Refinement of light-responsive transcript lists using rice oligonucleotide arrays: Evaluation of gene-redundancy. PLoS One 3, e3337 (2008).
Jung, K.-H. & An, G. Application of MapMan and RiceNet drives systematic analyses of the early heat stress transcriptome in rice seedlings. J. Plant Biol. 55, 436–449 (2012).
Stitt, M., Lilley, R. M., Gerhardt, R. & Heldt, H. W. Metabolite levels in specific cells and subcellular compartments of plant leaves. Methods Enzymol. 174, 518–552 (1989).
Mostajeran, A. & Rahimi-Eichi, V. Effects of drought stress on growth and yield of rice (Oryza sativa L.) cultivars and accumulation of proline and soluble sugars in sheath and blades of their different ages leaves. American-Eurasian J. Agric. Environ. Sci. 5, 264–272 (2009).
Zeeman, S. C., Smith, S. M. & Smith, A. M. The diurnal metabolism of leaf starch. Biochem. J. 401, 13–28 (2007).
Genty, B., Briantais, J. M. & Baker, N. R. The relationship between quantam yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochimica. et Biophysica. Acta. 990, 87–92 (1989).
Liu, J.-Y., Qiu, B.-S., Liu, Z.-L. & Yang, W.-N. Diurnal photosynthesis and photoinhibition of rice leaves with chlorophyll fluorescence. Acta Bot. Sin. 46, 552–559 (2004).
Huang, L. et al. Diurnal variations in gas exchange, chlorophyll fluorescence quenching and light allocation in soybean leaves: The cause for midday depression in CO2 assimilation. Sci. Hortic. 110, 214–218 (2006).
Pinheiro, C. & Chaves, M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 62, 869–882 (2011).
Jiang, D. & Xu, Y. Diurnal changes of photosynthetic rate, stomatal conductance and Rubisco in rice leaf. Acta Phytophysiol. Sin. 22, 94–100 (1996).
Nguyen, V. N. T., Moon, S. & Jung, K. H. Genome-wide expression analysis of rice ABC transporter family across spatio-temporal samples and in response to abiotic stresses. J. Plant Physiol. 171, 1276–1288 (2014).
Jung, K.-H., Gho, H.-J., Nguyen, M. X., Kim, S.-R. & An, G. Genome-wide expression analysis of HSP70 family genes in rice and identification of a cytosolic HSP70 gene highly induced under heat stress. Funct. Integr. Genom. 13, 391–402 (2013).
Utsumi, Y., Utsumi, C., Sawada, T., Fujita, N. & Nakamura, Y. Functional diversity of isoamylase oligomers: the ISA1 homo-oligomer is essential for amylopectin biosynthesis in rice endosperm. Plant Physiol. 156, 61–77 (2011).
Xu, D.-Q. et al. Overexpression of a TFIIIA-type zinc finger protein gene ZFP252 enhances drought and salt tolerance in rice (Oryza sativa L.). FEBS Lett. 582, 1037–1043 (2008).
Abe, N. et al. Relationships between starch synthase I and branching enzyme isozymes determined using double mutant rice lines. BMC Plant Biol. 14, 1–12 (2014).
Fujita, N. et al. Function and characterization of starch synthase I using mutants in rice. Plant Physiol. 140, 1070–1084 (2006).
Li, H. W., Zang, B. S., Deng, X. W. & Wang, X. P. Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta 234, 1007–1018 (2011).
Lua, C.-A. et al. The SnRK1A Protein Kinase Plays a Key Role in Sugar Signaling during Germination and Seedling Growth of Rice. Plant Cell 19, 2484–2499 (2007).
Zong, W. et al. Feedback regulation of ABA signaling and biosynthesis by a bZIP transcription factor targets drought resistance related genes. Plant Physiol. 171, 2810–2825 (2016).
Zhang, S. et al. Function of the HD-Zip I gene Oshox22 in ABA-mediated drought and salt tolerances in rice. Plant Mol. Biol. 80, 571–585 (2012).
Hossain, M. A. et al. The bZIP transcription factor OsABF1 is an ABA responsive element binding factor that enhances abiotic stress signaling in rice. Plant Mol. Biol. 72, 557–566 (2010).
Denby, K. & Gehring, C. Engineering drought and salinity tolerance in plants: Lessons from genome-wide expression profiling in Arabidopsis. Trends Biotechnol. 23, 547–552 (2005).
Urano, K., Kurihara, Y., Seki, M. & Shinozaki, K. ‘Omics’ analyses of regulatory networks in plant abiotic stress responses. Curr. Opin. Plant Biol. 13, 132–138 (2010).
Wang, Y. et al. Genome-wide identification of heat shock proteins (Hsps) and Hsp interactors in rice: Hsp70s as a case study. BMC Genom. 15, 344 (2014).
Nguyen, T. D. et al. Genome-wide identification and analysis of rice genes preferentially expressed in pollen at an early developmental stage. Plant Mol. Biol. 92, 71–88 (2016).
Redillas, M. C. et al. Accumulation of trehalose increases soluble sugar contents in rice plants conferring tolerance to drought and salt stress. Plant Biotechnol. Rep. 6, 89–96 (2012).
Geiger, D. R. & Servaites, J. C. Diurnal regulation of photosynthetic carbon metabolism in C3 plants. Annu. Rev. Plant Biol. 45, 235–256 (1994).
Hoshikawa K. Ripening. in The Growing Rice Plant (ed. Hoshikawa, K.) 258–259 (Tokyo, Nobunkyo, 1989).
Mangelsen, E. et al. Significance of light, sugar, and amino acid supply for diurnal gene regulation in developing barley caryopses. Plant Physiol. 153, 14–33 (2010).
Chaves, M., Flexas, J. & Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 103, 551–560 (2009).
Flexas, J., Bota, J., Loreto, F., Cornic, G. & Sharkey, T. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol. 6, 269–279 (2004).
Flexas, J. et al. Rapid variations of mesophyll conductance in response to changes in CO2 concentration around leaves. Plant Cell Environ. 30, 1284–1298 (2007).
Grzesiak, M., Grzesiak, S. & Skoczowski, A. Changes of leaf water potential and gas exchange during and after drought in triticale and maize genotypes differing in drought tolerance. Photosynthetica 44, 561–568 (2006).
Miyashita, K., Tanakamaru, S., Maitani, T. & Kimura, K. Recovery responses of photosynthesis, transpiration, and stomatal conductance in kidney bean following drought stress. Environ. Exp. Bot. 53, 205–214 (2005).
Wong, C. E. et al. Transcriptional profiling implicates novel interactions between abiotic stress and hormonal responses in Thellungiella, a close relative of Arabidopsis. Plant Physiol. 140, 1437–1450 (2006).
Schurr, U., Heckenberger, U., Herdel, K., Walter, A. & Feil, R. Leaf development in Ricinus communis during drought stress: Dynamics of growth processes, of cellular structure and of sink–source transition. J. Exp. Bot. 51, 1515–1529 (2000).
Geigenberger, P. & Stitt, M. Diurnal changes in sucrose, nucleotides, starch synthesis and AGPS transcript in growing potato tubers that are suppressed by decreased expression of sucrose phosphate synthase. Plant J. 23, 795 (2000).
Ramel, F., Sulmon, C., Gouesbet, G. & Couée, I. Natural variation reveals relationships between pre-stress carbohydrate nutritional status and subsequent responses to xenobiotic and oxidative stress in Arabidopsis thaliana. Ann. Bot. 104, 1323–1337 (2009).
Prasch, C. M. et al. β-amylase1 mutant Arabidopsis plants show improved drought tolerance due to reduced starch breakdown in guard cells. J. Exp. Bot. 66, 6059–6067 (2015).
Zanella, M. et al. β-amylase 1 (BAM1) degrades transitory starch to sustain proline biosynthesis during drought stress. J. Exp. Bot. 67, 1819–1826 (2016).
Wang, S., Liu, L., Chen, C. & Chen, L. Regulations of granule-bound starch synthase I gene expression in rice leaves by temperature and drought stress. Biol. Plant. 50, 537–541 (2006).
Smith, S. M. et al. Diurnal changes in the transcriptome encoding enzymes of starch metabolism provide evidence for both transcriptional and posttranscriptional regulation of starch metabolism in Arabidopsis leaves. Plant Physiol. 136, 2687–2699 (2004).
Park, S.-H. et al. OsbZIP23 and OsbZIP45, members of the rice basic leucine zipper transcription factor family, are involved in drought tolerance. Plant Biotechnol. Rep. 9, 89–96 (2015).
Kim, H. et al. A rice orthologue of the ABA receptor, OsPYL/RCAR5, is a positive regulator of the ABA signal transduction pathway in seed germination and early seedling growth. J. Exp. Bot. 63, 1013–1024 (2012).
Dey, A., Samanta, M. K., Gayen, S., Sen, S. K. & Maiti, M. K. Enhanced gene expression rather than natural polymorphism in coding sequence of the OsbZIP23 determines drought tolerance and yield improvement in rice genotypes. PLoS ONE 11, e0150763 (2016).
Thalmann, M. R. et al. Regulation of leaf starch degradation by abscisic acid is important for osmotic stress tolerance in plants. Plant Cell 28, 1860–1878 (2016).
Cao, P. et al. The rice oligonucleotide array database: An atlas of rice gene expression. Rice 5, 17 (2012).
Jefferson, R. A., Kavanagh, T. A. & Bevan, M. W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 6, 3901–3907 (1987).
Cho, L.-H., Yoon, J., Pasriga, R. & An, G. Homodimerization of Ehd1 is required to induce flowering in rice. Plant Physiol. 170, 2159–2171 (2016).
Kim, S.-W., Jeong, H.-J. & Jung, K.-H. Integrating omics analysis of salt stress-responsive genes in rice. Genes Genom. 37, 645–655 (2015).
Zulfugarov, I. S. et al. Enhanced resistance of PsbS-deficient Rice (Oryza sativa L.) to Fungal and Bacterial Pathogens. J. Plant Biol. 59, 616–626 (2016).
Nguyen, Q.-N. et al. Genome-wide identification and analysis of Catharanthus roseus RLK1-like kinases in rice. Planta 241, 603–613 (2015).
Chen, J.-Q., Meng, X.-P., Zhang, Y., Xia, M. & Wang, X.-P. Over-expression of OsDREB genes lead to enhanced drought tolerance in rice. Biotechnol. Lett. 30, 2191–2198 (2008).
Xiang, Y., Tang, N., Du, H., Ye, H. & Xiong, L. Characterization of OsbZIP23 as a key player of the basic leucine zipper transcription factor family for conferring abscisic acid sensitivity and salinity and drought tolerance in rice. Plant Physiol. 148, 1938–1952 (2008).
Yano, M. et al. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS. Plant Cell 12, 2473–2483 (2000).
Izawa, T., Takahashi, Y. & Yano, M. Comparative biology comes into bloom: Genomic and genetic comparison of flowering pathways in rice and Arabidopsis. Curr. Opin. Plant Biol. 6, 113–120 (2003).
Lee, Y.-S., Yi, J., Jung, K.-H. & An, G. Comparison of rice flowering-time genes under paddy conditions. J. Plant Biol. 59, 238–246 (2016).
Liu, W. Y. et al. The OsDHODH1 gene is involved in salt and drought tolerance in rice. J. Integr. Plant Biol. 51, 525–533 (2009).
Fukuda, A. et al. Function, intracellular localization and the importance in salt tolerance of a vacuolar Na+/H+ antiporter from rice. Plant Cell Physiol. 45, 146–159 (2004).
Diédhiou, C. J., Popova, O. V., Dietz, K.-J. & Golldack, D. The SNF1-type serine-threonine protein kinase SAPK4 regulates stress-responsive gene expression in rice. BMC Plant Biol. 8, 49 (2008).
Huang, X. et al. OsSLI1, a homeodomain containing transcription activator, involves abscisic acid related stress response in rice (Oryza sativa L.). Sci. World J. 2014, 809353 (2014).
You, J. et al. The SNAC1-targeted gene OsSRO1c modulates stomatal closure and oxidative stress tolerance by regulating hydrogen peroxide in rice. J. Exp. Bot. 64, 569–583 (2013).
Zhang, H. et al. Cloning and characterization of two rice long-chain base kinase genes and their function in disease resistance and cell death. Mol. Biol. Rep. 40, 117–127 (2013).
Chou, W.-L. et al. Divergence of the expression and subcellular localization of CCR4-associated factor 1 (CAF1) deadenylase proteins in Oryza sativa. Plant Mol. Biol. 85, 443–458 (2014).
Huang, J. et al. A TFIIIA-type zinc finger protein confers multiple abiotic stress tolerances in transgenic rice (Oryza sativa L.). Plant Mol. Biol. 80, 337–350 (2012).
Lin, M.-Y. et al. A positive feedback loop between HSP101 and HSA32 modulates long-term acquired thermotolerance illustrating diverse heat stress responses in rice varieties. Plant Physiol. 164, 2045–2053 (2014).
Du, H. et al. Characterization of the β-carotene hydroxylase gene DSM2 conferring drought and oxidative stress resistance by increasing xanthophylls and abscisic acid synthesis in rice. Plant Physiol. 154, 1304–1318 (2010).
Saika, H., Matsumura, H., Takano, T., Tsutsumi, N. & Nakazono, M. A point mutation of Adh1 gene is involved in the repression of coleoptile elongation under submergence in rice. Breeding Sci. 56, 69–74 (2006).
You, J., Hu, H. & Xiong, L. An ornithine δ-aminotransferase gene OsOAT confers drought and oxidative stress tolerance in rice. Plant Sci. 197, 59–69 (2012).
Tang, Y. et al. Knockdown of OsPAO and OsRCCR1 cause different plant death phenotypes in rice. J. Plant Physiol. 168, 1952–1959 (2011).
Park, S.-Y. et al. The senescence-induced staygreen protein regulates chlorophyll degradation. Plant Cell 19, 1649–1664 (2007).
Iwamoto, M., Higo, K. & Takano, M. Circadian clock- and phytochrome-regulated Dof-like gene, Rdd1, is associated with grain size in rice. Plant Cell Environ. 32, 592–603 (2009).
Deng, Z.-Y. & Wang, T. OsDMC1 is required for homologous pairing in Oryza sativa. Plant Mol. Biol. 65, 31–43 (2007).
Lo, S.-F. et al. A novel class of gibberellin 2-oxidases control semidwarfism, tillering, and root development in rice. Plant Cell 20, 2603–2618 (2008).
Zhao, Z. et al. A role for a dioxygenase in auxin metabolism and reproductive development in rice. Dev. Cell 27, 113–122 (2013).
Cook, F. R., Fahy, B. & Trafford, K. A rice mutant lacking a large subunit of ADP-glucose pyrophosphorylase has drastically reduced starch content in the culm but normal plant morphology and yield. Funct. Plant Biol. 39, 1068–1078 (2012).
Santana, J. O. et al. Characterization of the legumains encoded by the genome of Theobroma cacao L. Plant Physiol. Biochem. 98, 162–170 (2016).
Liu, C., Wu, Y. & Wang, X. bZIP transcription factor OsbZIP52/RISBZ5: A potential negative regulator of cold and drought stress response in rice. Planta 235, 1157–1169 (2012).
Yara, A. et al. Disease resistance against Magnaporthe grisea is enhanced in transgenic rice with suppression of ω-3 fatty acid desaturases. Plant Cell Physiol. 48, 1263–1274 (2007).
Joo, J., Lee, Y. H., Kim, Y. K., Nahm, B. H. & Song, S. I. Abiotic stress responsive rice ASR1 and ASR3 exhibit different tissue-dependent sugar and hormone-sensitivities. Mol. Cells 35, 421–435 (2013).
Sun, S. et al. A constitutive expressed phosphate transporter, OsPht1; 1, modulates phosphate uptake and translocation in phosphate-replete rice. Plant Physiol. 159, 1571–1581 (2012).
Yoshie, Y. et al. Function of the rice gp91phox homologs OsrbohA and OsrbohE genes in ROS-dependent plant immune responses. Plant Biotechnol. 22, 127–135 (2005).
Kim, S. G. et al. Overexpression of rice isoflavone reductase-like gene (OsIRL) confers tolerance to reactive oxygen species. Physiol. Plant. 138, 1–9 (2010).
Li, W. et al. Rice RING protein OsBBI1 with E3 ligase activity confers broad-spectrum resistance against Magnaporthe oryzae by modifying the cell wall defence. Cell Res. 21, 835–848 (2011).
Yamaji, N., Mitatni, N. & Ma, J. F. A transporter regulating silicon distribution in rice shoots. Plant Cell 20, 1381–1389 (2008).
Chen, Z. et al. Mn tolerance in rice is mediated by MTP8. 1, a member of the cation diffusion facilitator family. J. Exp. Bot. 64, 4375–4387 (2013).
Jung, K.-H. et al. Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol. 44, 463–472 (2003).
Nishikawa, Y. PGR5-dependent cyclic electron transport around PSI contributes to the redox homeostasis in chloroplasts rather than CO2 fixation and biomass production in rice. Plant Cell Physiol. 53, 2117–2126 (2012).
Yan, W.-H. et al. A major QTL, Ghd8, plays pleiotropic roles in regulating grain productivity, plant height, and heading date in rice. Mol. Plant 4, 319–330 (2011).
Zheng, L., Yamaji, N., Yokosho, K. & Ma, J. F. YSL16 is a phloem-localized transporter of the copper-nicotianamine complex that is responsible for copper distribution in rice. Plant Cell 24, 3767–3782 (2012).
Lee, S., Kim, J., Han, J. J., Han, M. J. & An, G. Functional analyses of the flowering time gene OsMADS50, the putative SUPPRESSOR OF OVEREXPRESSION OF CO 1/AGAMOUS-LIKE 20 (SOC1/AGL20) ortholog in rice. Plant J. 38, 754–764 (2004).
Lee, S. K. et al. Loss of cytosolic fructose‐1, 6-bisphosphatase limits photosynthetic sucrose synthesis and causes severe growth retardations in rice (Oryza sativa). Plant Cell Environ. 31, 1851–1863 (2008).
Li, R.-Q. et al. A down-regulated epi-allele of the genomes uncoupled 4 gene generates a xantha marker trait in rice. Theor. Appl. Genet. 127, 2491–2501 (2014).
Lu, Y., Song, Z., Lü, K., Lian, X. & Cai, H. Molecular characterization, expression and functional analysis of the amino acid transporter gene family (OsAATs) in rice. Acta Physiol. Plant. 34, 1943–1962 (2012).
Masumoto, C. et al. Phosphoenolpyruvate carboxylase intrinsically located in the chloroplast of rice plays a crucial role in ammonium assimilation. Proc. Natl. Acad. Sci. USA 107, 5226–5231 (2010).
Zhou, Q. et al. Knockdown of GDCH gene reveals reactive oxygen species-induced leaf senescence in rice. Plant Cell Environ. 36, 1476–1489 (2013).
Todaka, D. et al. Rice phytochrome-interacting factor-like protein OsPIL1 functions as a key regulator of internode elongation and induces a morphological response to drought stress. Proc. Natl. Acad. Sci. USA 109, 15947–15952 (2012).
Ishii, T. et al. OsLG1 regulates a closed panicle trait in domesticated rice. Nat. Genet. 45, 462–465 (2013).
Rong, H. et al. The Stay-Green Rice like (SGRL) gene regulates chlorophyll degradation in rice. J. Plant Physiol. 170, 1367–1373 (2013).
Li, Y. et al. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat. Genet. 43, 1266–1269 (2011).
Yamaguchi, T. et al. The YABBY gene DROOPING LEAF regulates carpel specification and midrib development in Oryza sativa. Plant Cell 16, 500–509 (2004).
Hu, X. et al. The U-box E3 ubiquitin ligase TUD1 functions with a heterotrimeric G α subunit to regulate brassinosteroid-mediated growth in rice. PLoS Genet. 9, e1003391 (2013).
Chen, W. et al. Genome-wide association analyses provide genetic and biochemical insights into natural variation in rice metabolism. Nat. Genet. 46, 714–721 (2014).
Acknowledgements
This work was supported by grants from the Next-Generation BioGreen 21 Program (PJ01182602 to KHJ), the Rural Development Administration; and from the Basic Science Research Program (NRF-2016R1D1A1A09919568 to KHJ) and the Mid-Career Researcher Program (NRF-2017R1A2B4009687 to JSJ), National Research Foundation, Republic of Korea.
Author information
Authors and Affiliations
Contributions
K.H.J., G.A. and J.S.J. designed the research. S.W.K., H.J.J. and S.K.L. performed the experiments. S.W.K. and S.K.L. analyzed the data. S.W.K., S.K.L., K.H.J., and J.S.J. wrote the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kim, SW., Lee, SK., Jeong, HJ. et al. Crosstalk between diurnal rhythm and water stress reveals an altered primary carbon flux into soluble sugars in drought-treated rice leaves. Sci Rep 7, 8214 (2017). https://doi.org/10.1038/s41598-017-08473-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-08473-1
This article is cited by
-
Chalkiness and premature controlled by energy homeostasis in OsNAC02 Ko-mutant during vegetative endosperm development
BMC Plant Biology (2024)
-
Transcriptome and WGCNA reveal hub genes in sugarcane tiller seedlings in response to drought stress
Scientific Reports (2023)
-
CO2 uptake and chlorophyll a fluorescence of Suaeda fruticosa grown under diurnal rhythm and after transfer to continuous dark
Photosynthesis Research (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
