
Abstract
Methane concentration in caves is commonly much lower than the external atmosphere, yet the cave CH4 depletion causal mechanism is contested and dynamic links to external diurnal and seasonal temperature cycles unknown. Here, we report a continuous 3-year record of cave methane and other trace gases in Jenolan Caves, Australia which shows a seasonal cycle of extreme CH4 depletion, from ambient ~1,775 ppb to near zero during summer and to ~800 ppb in winter. Methanotrophic bacteria, some newly-discovered, rapidly consume methane on cave surfaces and in external karst soils with lifetimes in the cave of a few hours. Extreme bacterial selection due to the absence of alternate carbon sources for growth in the cave environment has resulted in an extremely high proportion 2–12% of methanotrophs in the total bacteria present. Unexpected seasonal bias in our cave CH4 depletion record is explained by a three-step process involving methanotrophy in aerobic karst soil above the cave, summer transport of soil-gas into the cave through epikarst, followed by further cave CH4 depletion. Disentangling cause and effect of cave gas variations by tracing sources and sinks has identified seasonal speleothem growth bias, with implied palaeo-climate record bias.
Similar content being viewed by others


Introduction
Cave methane depletion is near universal for 44 caves reported1,2,3,4,5,6. A methanotrophic bacterial oxidation mechanism was first proposed to account for rapid methane loss over a few hours1,2,5 in opposition to a recently proposed radiolytic CH4 destruction mechanism4.
A steady-state model of in-situ depletion of methane by cave dwelling methanotrophs or radiolytic destruction by ions derived from radon decay might be expected from the year round constant cave air temperature ± 2 °C. However, an in-situ steady-state model is inconsistent with our observations of strong diel and seasonal bias in CO2, CH4 and other trace gases.
Cave CO2 variations are strongly anti-correlated with those of CH4. CO2 is enhanced relative to the external ambient atmosphere, to as high as 10,000 ppm in summer but remains near external ambient in winter. CO2 has several potential sources, including breath from cave visitors, ground air7, speleothem growth8,9 and karst soil gas10. CO2 sources that would be expected to show antithetical behaviour relative to CH4 are ground air7 and the overlying soil gas, where CO2 is increased by plant root and microbial respiration and CH4 is depleted by microbial oxidation.
Dynamic cave ventilation
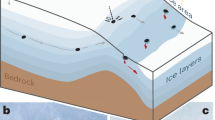
Convective cave ventilation11 driven by temperature contrast between the cave and the external environment also plays an important role in determining cave trace gas concentrations. Jenolan Caves, located approximately 100 km west of Sydney, Australia are representative of many mid-latitude cave locations exhibiting diurnal, synoptic and seasonal temperature cycles of the ambient atmosphere. These cycles create a contrast between the external temperature and the relatively constant cave temperature (11 °C ± 2 °C) to drive cyclical bi-directional convective ventilation through upper and lower cave openings (Fig. 1, Supplementary Fig. 1).

Top panel is a schematic cross-section through Chifley Cave showing the dominant convective air-flow directions for summer (red) and winter (blue). Air-flow along major passages may reverse direction daily in response to a reversal of the cave to external temperature difference, without a time lag (<15 mins). The slower passage of air from soil to cave along small cracks and fissures in summer has a longer time lag of 1–6 hrs between cave - external temperature reversal and the soil air peak in Lower Katies Bower (low CH4, high CO2, high N2O, high Rn). Lower panel is a scale diagram of continuous flow air sampling locations relative to the CRDS & FTIR instruments.
Our continuous data for CH4, CO2, δ13C-in-CO2, Rn, CO, and N2O trace gases from Chifley Cave (Fig. 1), represents a well-ventilated tourist cave hosting several tours per day. We also measured cave air-temperature, air-flow, air-pressure, drip-rate, and drip pH continuously for 3 years from September 2012 to July 2015. High precision gas analysers for CO2, CH4, CO, N2O and δ13C-in-CO2 (CRDS and FTIR, see methods) were installed approximately 20 m from the lower (Grand Arch) entrance and sequentially analysed air sampled once per hour drawn along tubing from four locations in the cave (Fig. 1) and 2 external reference locations. The karst soil overlying Chifley Cave was continuously monitored for soil temperature, soil water fraction and CO2, supplemented by hourly CH4 and δ13C-in-CO2 measured during a 10 day experiment to define soil function as a response to weather and gas composition as a potential source for cave air (Supplementary Fig. 2a,b).
The differential air density at different air temperatures inside the cave compared to external temperature drives bi-directional convective air-flow through the major passages, measured at the Flitch of Bacon (Fig. 1). The open passages provide the path for the bulk of cave air-flow. Air flows more slowly through minor passages & narrow fissures and ultimately through a multitude of cracks and through the soil. Summer air-flow is dominantly from the ridge overlying Chifley Cave including soil gas seepage with winter air-flow in the opposite direction (Fig. 2). On any one day throughout the year air-flow may change direction abruptly in response to changing external weather (temperature). In spring and autumn diurnal external temperature may dip below cave temperature and also rise above cave temperature (Fig. 2) causing a rapid (~15 min) change in air-flow direction along open passages measured at Flitch of Bacon (Fig. 1). The time lag between air-flow direction change and emerging CO2 peak in Lower Katies Bower is 1–6 hrs, giving an approximate soil to cave gas transit time. Soil to cave minimum distance is ~50–150 m compared to 268 m through major passages. The spring and autumn pattern of diurnal change in air flow direction and consequently large changes in CO2 Lower Katies Bower (1,000–5,000 ppm) is masked in summer and winter due to dominant flow directions. Normal soil gas flow into Chifley Cave is by wholesale mass transfer rather than gas diffusion. Rarely does external temperature equate to internal temperature for an extended period which may induce diffusion.

CH4 and CO2 CRDS continuous measurements (8 minute averages, recorded hourly) from LKB in Chifley Cave over 3 years from September 2012 to August 2015. Methane measurement error ± 20 ppb and carbon dioxide ± 5 ppm. Winter air-flow (blue infill) shows external ambient air drawn into Chifley Cave from the “Grand Arch” cave entrance has a greater velocity and longer duration than Summer air-flow (red infill) in the opposite direction. External weather has a direct immediate impact on cave CO2 & CH4 (eg. week 1 February 2015) linking cave trace gas composition to external weather and climate.
Cave and karst soil CH4, CO2 and Rn links to air-flow & external weather
Methane in Lower Katies Bower (LKB) chamber within Chifley Cave was totally depleted from the normal atmospheric mole fraction of ~1,775 ppb to 11 ± 20 ppb in summer (Fig. 2, Table 1), correlating inversely with carbon dioxide, which sometimes exceeded 10,000 ppm in LKB in summer. In winter methane depletion in LKB was approximately half compared to external air, dropping to 550-1,150 ppb, while CO2 approached ambient air, ~570 ppm in LKB. Both CO2 and CH4 also displayed daily antithetic cycles of approximately one third of the full concentration range evident during autumn, winter, and spring. Diurnal CO2 and CH4 cycles were largely absent during summer. A synoptic cycle is clearly seen linking the passing of external weather systems to cave CO2 and CH4 (Figs 2 and 3). In addition to CH4, CO was also totally depleted in the cave relative to ambient air (Figs 4 and 5). Cave CO was always near zero and did not display seasonality.

Chifley Cave, Lower Katies Bower trace gases CH4, CO2, and Rn, relative to cave air flow, cave and external temperature. The summer to winter period January 2015 – June 2015 illustrates the direct link between external temperature and cave CH4, CO2, and Rn through diurnal, synoptic & seasonal cycles.

Late summer time series of CO, CO2, Rn, N2O, and δ13C – CO2 from Lower Katies Bower corresponding with correlation plots (5/3/15–18/4/15). CO is totally depleted (measurement error ± 5 ppb) within Chifley Cave relative to a variable background ambient concentration of 75–110 ppb. CO depletion is not seasonally variable. Rn, N2O and δ13C – CO2 (−24.8‰ VPDB) gas tracers each correlate strongly with CO2 and identify karst soil overlying Chifley Cave as the only likely source of high CO2 in the cave in summer (Figs 1 and 2). Late summer bi-directional air-flow shows large synchronous fluctuations in CO2 and gas tracers.

Winter time series of CO, CO2, Rn, N2O, and δ13C – CO2 from Lower Katies Bower corresponding with correlation plots (30/6/15–17/7/15). CO2 & Rn is 10x lower in winter. Rn and N2O soil gas tracers do not correlate with CO2. Air-flow is dominantly from the Grand Arch (Figs 1 and 2) diluting cave air with ambient air. Winter uni-directional air-flow with low CO2 induces speleothem growth (δ13C – CO2 = −19.5‰ VPDB, Fig. 6) and shows large synchronous fluctuations in CO2 not Rn & N2O soil gas tracers indicating a small cave source of CO2 from speleothem growth.
Radon may emanate directly from limestone and cave sediments or be transported from overlying karst soil to the cave environment. Low uranium concentration in limestone and a short effective Rn diffusion path length favour a cave sediment (clay) or karst soil Rn source by a factor of ~100 over a limestone source12. This Rn source ratio was tested in Chifley Cave by experiments in which chambers were placed over cave surfaces and the chamber monitored for in-situ accumulation or depletion of Rn and CH4 (Supplementary Fig. 3). Synchronous measurement of Rn in sealed chambers covering a 0.23 m2 area of karst soil and 0.22 m2 of cave floor showed a karst soil to Chifley Cave Rn source ratio of 133 (Supplementary Table 1). Thus, Rn is a selective tracer for external karst soil air entering Chifley Cave, rather than being produced in-situ then diluted by winter ventilation4,13 with external air. Chifley Cave (Fig. 3), Ojo Guarena Cave4, Spain (ref.4, Supplementary Fig. 2) and Hollow Ridge Cave, Florida (ref.13, Fig. 4) also show highly correlated, relatively high, yet variable Rn and CO2 in summer and low stable Rn and CO2 in winter.
Rn geochemically discriminates between soil air and ground air7. Residual karst soils are composed of minerals that have not dissolved during limestone weathering, typically clays and Fe-oxides with organic carbon added by surface plants and soil bacteria. If ground air is distinct from soil air it is not expected to be in contact with the clays and Fe-oxides in the soil to inherit characteristically high Rn from a soil source.
Aerobic karst soil above Chifley Cave was depleted in methane (500–800 ppb) (Supplementary Fig. 2b) relative to atmospheric methane 1,750–1,800 ppb, typical of many aerobic karst and forest soils6,14,15. During a 10 day summer monitoring period, soil CH4 was observed to decrease rapidly, within 24hrs, in response to a small 4 °C soil temperature increase approximating the inverse soil diurnal CO2 response to temperature (Supplementary Fig. 2b).
Karst soil above Chifley Cave showed a variable seasonal CO2 pattern reflecting temperature and moisture-dependent soil primary productivity, sourced from plant root respiration and microbial organic matter degradation. Karst soil CO2 measured in-situ had a stable isotope label δ13C of −24‰ VPDB (Supplementary Fig. 2b) and did not vary significantly over the summer measurement period in response to large external temperature or soil moisture variations. Convective wholesale mass transfer of CO2 from soil to cave does not isotopically fractionate CO2. Potential fractionation by slow diffusion through porous media along a chemical gradient is not consistent with the measured air-flow regime and is not supported by matching soil and cave δ13C – CO2 measurements. In Chifley Cave with a summer ventilation pattern drawing air from above the cave with a soil source, the same (δ13C −24‰ VPDB) isotopic signature was observed in Lower Katies Bower (−24.8‰ Keeling intercept Fig. 6). This isotopic CO2 signature provides a second stable soil air tracer (after Rn) distinct from the well-mixed external atmosphere at Jenolan (CO2 δ13C −9.1 to −10.1‰ VPDB). A winter air-flow pattern in the opposite direction drew cool ambient air with low CO2, ~425 ppm into Chifley Cave from the Grand Arch cave entrance. Low CO2 in the cave atmosphere induces outgassing16 of CO2 from drip-waters during speleothem growth (Fig. 6, CO2 δ13C −19.5‰ VPDB) raising CO2 to ~560 ppm.

A Keeling plot of δ13C – CO2 vs 1/CO2 from Flitch of Bacon. for two time periods; autumn 5/3/15–18/4/15, least squares line fit (red), n = 935 and winter 30/6/15–17/7/15, least squares line fit (blue), n = 358. Data points are colour coded to represent air-flow direction and velocity. The Keeling plot intercept provides the δ13C – CO2 of the CO2 source component mixing with air. The source of high CO2 in LKB during autumnal (April) bi-directional airflow has δ13C – CO2 = −24.8‰ VPDB. The source of cave CO2 during winter with near uni-directional airflow has δ13C – CO2 = −19.5‰ VPDB, representing speleothem growth CO2 degassing within the Cave (δ13C drip water −HCO3 (aq) −10.0‰ + fractionation16 −HCO3 (aq) - CO2 (g) at 11 °C −9.48‰ = −19.5‰ VPDB).
N2O provided a third selective tracer for soil air in the cave. N2O is produced in soils by nitrifying and de-nitrifying bacteria17,18 and is not produced or consumed inside caves from limestone or cave sediments (Fig. 7). N2O thus acts as a conservative tracer of soil gas. N2O is also closely correlated with CO2 and Rn (Figs 4 and 5) with an N2O:CO2 ratio of around 0.1 ppb ppm−1, typical of other soils19. The unchanging N2O:CO2 ratio of around 0.1 ppb ppm−1 between literature values for soils and cave measurements suggests no other source of N2O or CO2, such as ground air7, on the path between soil and cave is necessary to explain Chifley Cave data. Figure 7 shows the close summer N2O - CO2 correlation sourced from soil with air-flow magnitude & direction shown in shades of red. When N2O is measured inside Chifley Cave in winter (Fig. 7, blue) typically with air-flows in the opposite direction and an ambient atmospheric composition there is no soil gas contribution. N2O is fixed at atmospheric composition (Table 1, N2O = 329 ppb range ± 2 ppb error ± 5 ppb).

Correlation plot of N2O vs CO2 from Flitch of Bacon (0.5 × 2.0 m passage) for two time periods; autumn 5/3/15–18/4/15, least squares line fit (red), n = 935 and winter 30/6/15–17/7/15, least squares line fit (blue), n = 358. Data points are colour coded to represent air-flow direction and velocity. Only air from above Chifley Cave (ambient + soil air, red) contains N2O greater than ambient, indicative of soil air. Air-flow in the opposite direction (blue) is tightly clustered around ambient composition with no addition or depletion of N2O within Chifley Cave.
Collectively, Rn, N2O and high concentration ( > 5,000 ppm) CO2 with an isotopic label of δ13C −24‰ VPDB unequivocally identified karst soil as the source of these trace gases in air, emerging in LKB by convective suction of air in summer. However, a karst soil source of air with methane levels of 400–800 ppb does not fully explain total CH4 depletion in LKB in summer. Further in-situ cave CH4 depletion is inferred.
Methanotrophic bacteria
Methanotrophic bacteria living on limestone and sediment cave surfaces, or in the overlying karst soil, may cause the observed depletion of methane in cave air. Methane oxidation by bacteria in soils and sediments is usually carried out by Gammaproteobacteria and Alphaproteobacteria20, though there is also recent evidence for methanotrophy by members of the Verrucomicrobia21. In natural soil environments that are not exposed to elevated levels of methane, the upland soil “USC-cluster” methanotrophs dominate. These grow well at atmospheric-methane concentrations but respond slowly to changes in methane availability, and have proved difficult to cultivate in the laboratory22. External karst soils overlying Gaden cave6, New South Wales showed that the methanotroph population in the upper 10 cm was dominated by Alphaproteobacteria, while Gammaproteobacteria were prevalent at 30 cm depth, and methanotrophic Verrucomicrobia were present at both depths. However, internal cave formations & sediments bacteria were not reported6.
This is the first study to investigate the presence, spatial and temporal distribution of cave methanotrophs. The presence of the bacterial methane monooxygenase gene pmoA gene was measured in internal cave soils and on cave wall and floor surfaces within Chifley Cave. The pmoA gene was detected in all soils and sediments tested, but not in samples swabbed from wet speleothem surfaces, indicating the presence of methanotrophic bacteria in the sediments, but not commonly on the cave walls. Similar results were obtained from other caves in the Jenolan complex, and from similar karst caves at Wombeyan, 100 km south of Jenolan. Sequencing of the pmoA gene diversity from four sites within Chifley Cave revealed a total of 88 new methanotroph OTUs (operational taxonomic units), using 86% amino acid identity as a cut-off for OTU delineation. These sequences fell into four known pmoA clades and two previously undetected clades (Supplementary Fig. 4, Supplementary Table 2). Sequences falling within the four known clades were related to sequences from methanotrophs that had previously been found in samples from upland soils, and which are predicted to comprise organisms adapted to methane metabolism at atmospheric concentrations23. There was no evidence for verrucomicrobial methanotrophs in Chifley cave, but it should be noted that the pmoA genes of this group are very poorly characterized to date.
The total bacterial population in the soils was 107−109 16S rRNA gene copies per gram soil, and did not change significantly during the year. Bacteria were highest in the dampest sediments (Pool and Flitch of Bacon), with tenfold lower populations in the deeper cave regions (Katie’s Bower). The proportion of methanotrophs in the total bacterial population was highest in the deeper cave regions, Lower Katies Bower. By contrast, the proportion of methanotrophs in these sediments (including those belonging to both Gammaproteobacteria, and Alphaproteobacteria, but largely excluding those of the so-called “upland soil cluster”) showed considerable variation in population during the year, making up around 4% of the total bacterial population in the summer months, and 0–2% in the winter months. (Supplementary Fig. 5). The population of “upland soil cluster” methanotrophs was relatively stable throughout the year, as previously reported for type II methanotrophs in other environments20.
Initial rates of methane depletion from laboratory microcosms containing Chifley Cave sediments were 0.8–3.9 nmol g−1 h−1, (Supplementary Table 3) confirming cave methanotrophic organisms actively oxidize methane. Complete depletion of 1,000 ppm methane was observed within 20 days, with no depletion seen for sterilized samples. Similar results were obtained at atmospheric methane concentrations (Supplementary Fig. 6a). Methane is depleted from laboratory air, 2 ppm, in the experiment reaction chamber at a rate of 0.51 mg CH4 m−2 h−1, five times faster than methane oxidation in aerobic upland soils15 which is rarely higher than 0.1 mg CH4 m−2 h−1. Comparative cave chamber experiments24 utilising a small 0.22 m2 sealed area of Chifley Cave rock & sediment surface had a CH4 depletion rate of 0.2 nmol m−2 s−1 (Supplementary Fig. 6b).
Causal mechanism
Initial reporting of methane depletion in Jenolan Caves1 proposed bacterial consumption to explain the observed rapid loss of methane over a few hours. At St Michaels Cave, Gibraltar2, isotopic measurement of cave air depleted in CH4 shows13C enrichment in remaining CH4 to give δ13C up to −15‰ VPDB from an initial −47‰ in ambient air. This demonstrates an inverse correlation of CH4 abundance and isotopic12C depletion2,6 characteristic of Rayleigh distillation25 by biological kinetic fractionation26,27 consistent with methanotrophy.
An alternate radiolytic mechanism of radon generating atmospheric ions, causing methane oxidation has recently been proposed to explain a spatial correlation between low methane and relatively high ion concentrations in Spanish caves4. Methanotrophy was dismissed for these Spanish caves based on the absence of detected methanotrophs in remote subterranean locations and an interpreted inability to sustain methanotrophs’ metabolic requirements; this is contrary to ecophysiological features of microorganisms in natural ecosystems28,29.
To test the competing hypotheses for cave CH4 depletion we conducted laboratory experiments with and without methanotrophs and Rn (Supplementary Fig. 6a, Exp. 1 & 2). We presume Rn is the source of elevated ions observed in Spanish caves and measured an approximate proportional relationship; 1 Bq m−3 ≈ 10 ions cm−3. In these experiments no CH4 loss was observed at 1,000 times the Rn concentration in Spanish caves4 (ref.26 and Supplementary Fig. 6a, Exp. 1). Further increasing the radiolytic dose equivalent to an extreme 17 kGy, 2.7 × 106 times cave Rn4, reduces CH4 in air by 19%. The same small reaction chamber, populated with 250 g of Chifley Cave mud, sediment & methanotrophs (methane monooxygenase pmoA gene 1.5 × 106 copies g−1 sediment) consumed methane from 2,040 ppb in laboratory air to 293 ppb over 5.75 days (Supplementary Fig. 6a Exp. 2a). A control experiment under the same conditions in which the reaction chamber was sterilised to kill methanotrophs showed no methane depletion after 6 days (Supplementary Fig. 6a Exp. 2b). These experiments together demonstrate both a direct link to methanotroph depletion of CH4 and no link to Rn-initiated depletion.
Chemical kinetic arguments further confirm that the maximum possible reaction rates for destruction of CH4 by atmospheric ions, sourced from Rn at the observed concentrations are several orders of magnitude too slow to account for the observed destruction lifetime of CH4 of a few hours (see Supplementary information).
We conclude that methanotrophs are solely responsible for cave methane depletion and ions derived from Rn play no significant part in the cave CH4 sink.
Cave methane depletion process
From our data for Chifley Cave we resolve that a three-step process is responsible for seasonal total CH4 depletion.
The seasonal ventilation difference between summer suction of karst soil gas into Lower Katies Bower compared to winter expulsion of cave air from LKB provides a mechanism for seasonally different sources of air in LKB (Fig. 1). External air temperature has a rapid, compounding effect on cave CO2 and CH4 by influencing primary productivity of the source soil gas composition and modulating the strength of convective air-flow for transport from soil to LKB. Diurnal, synoptic and seasonal cyclic variations in observed trace gas concentrations are due to temperature-driven convective ventilation patterns.
Total methane depletion in summer is a consequence of initial partial depletion in the overlying karst soil by aerobic methanotrophs14,15 followed by slow interaction with cracks and fissure cave surfaces along the transport pathway to emerge in LKB. Further cave CH4 depletion occurs in-situ by methanotrophy on cave surfaces. Path length bias (Supplementary information text, Supplementary Fig. 7, Supplementary Table 4) and greater air residence time (Figs 1 and 8a,b) due to localised micro-temperature stratification in LKB may also contribute to the extreme summer methane depletion.

(a) Excerpt of high-resolution (3 cm) 3D mapping data30,44 for Jenolan Caves. This 2D image shows the details of passage connection from Grand Arch (left corner) through Chifley and Imperial Caves. Cut-away shows internal detail of 3D digital model, illustrating the complex volumetric and surface area relationships used to calculate air – cave surface interaction for bi-directional convective flow paths. Blue circle positions Lower Katies Bower. (b) Physical 3D printed model of Lower Katies Bower produced from high-resolution 3D mapping data30,44. Figure 1 schematically illustrates the thermal stratification in Lower Katies Bower under different convective air-flow regimes causing changes in air residence time.
A simple measure of the temperature contrast between cave air and the external atmosphere coupled with the vertical difference in elevation between upper and lower cave openings gives an estimate of the strength of bi-directional convective cave ventilation (Table 1). Caves may also vary in convective ventilation response due to differing air path length and tortuosity30. Seasonal bias in convective cave ventilation strength and direction is likely to follow latitude and climatic zones11 with an increasing proportion of soil gas evident in low-mid latitude caves. Soil source gas composition, particularly CO2 and CH4 will vary with soil microbial primary productivity. Quantification of the processes described above give a guide to likely estimates of trace gases in caves.
The Chifley Cave methanotrophs reported here, which grow in cave soils at atmospheric methane concentrations (Type II methanotrophs), may be typical of many caves but are not unique to this cave or cave system. We have identified similar levels of methanotrophs in the Orient, Lucas, and Temple of Baal caves at Jenolan, and in the Wolondilly and Junction caves at Wombeyan Caves. The 88 new methanotrophic phylotypes identified in Chifley Cave are therefore likely to be representative of a larger group of limestone cave methanotrophs that have not yet been studied in detail, and are enriched in these environments. Extreme bacterial selection due to the absence of alternative carbon sources for growth in the cave environment has resulted in methanotrophs constituting 2–12% of the total bacteria present.
Caves and karst soils constitute an unrecognised31 aerobic soil methane sink14,15 and may contribute to uncertainties in aerobic soil sink climate feedback dynamics.
The direct links between dynamic methanotrophy and soil primary biological productivity dependent upon temperature and precipitation are observed and amplified in the cave system. When extrapolated to aerobic soils these insights may help to explain the observed anomalies in global atmospheric CH4 growth32. Aerobic soil response to climate change remains uncertain with CH4 feedback contributions from temperature and particularly precipitation strongly effecting aerobic soil CH4 sink dynamics. An example of this sensitivity is the reversal of seasonal aerobic karst soil CO2 and CH4 maxima and minima between Jenolan Caves and Wellington Caves6 175 km to the North West, where primary biological productivity is limited in summer by soil water availability.
An exception to universal limestone cave methane depletion is noted33 for Movile Cave, Romania where Type I methanotrophs have also been described associated with a cave atmosphere at 1–2% methane33,34. Movile Cave is fed by hydrothermal groundwater rich in H2S, and this distinguishes it from other karst caves since the sulphide supports an unusual in-situ chemoautotrophic ecosystem. Microbial primary productivity acts as the food source for 48 species of cave-adapted terrestrial and aquatic invertebrates in Movile Cave33. By contrast, microbial life in most limestone caves is limited by the oligotrophic nature of the environment and organisms rely on reduced compounds in air, such as methane or carbon monoxide, or on chemoautrophy, oxidizing reduced metal ions present in the rock35.
Speleothem growth is favoured by low CO2 in the cave atmosphere. Definition of the soil source (Rn, N2O, δ13C CO2 tracers) and high concentration of CO2 with its isotopic label drawn into Chifley Cave in summer without change (soil & cave, δ13C −24‰ PDB) suggests speleothem growth inhibition. Winter speleothem growth is evident (Fig. 6) leading to the inference of strong seasonal bias in speleothem growth for convection-ventilated caves. Projection of this result in Chifley Cave to other caves and impact on palaeo-climate records is conditional upon the specific speleothem location in the cave ventilation pattern driven by local external weather and climate.
Methods
Cave gas measurement method
We designed our study to take advantage of technology advances in high precision field-deployable CRDS and FTIR spectrometers to continuously measure trace gases (CH4, CO, N2O, CO2 and δ13C-CO2) in a cave for the first time. This approach overcomes problems of 1. data paucity for spot sampling (ie monthly 12 samples per year) which cannot resolve diurnal and synoptic variations compared to hourly continuous 8,760 measurements per year per location × 6 2. the problem of individual samples taken sequentially during a sampling field trip not being an accurate snapshot in time due to changing cave ventilation (<1 hour) 3. Spot sampling of cave environments is also inherently difficult because of the human sampler breathing and potentially effecting results. Our trace gas data come from four cave locations, Lower Katies Bower (LKB) 120 m from the Grand Arch and Upper Katie’s Bower (UKB), Flitch of Bacon (FOB), Entrance path) (Fig. 1) together with two external reference locations in the ambient atmosphere outside the cave. A large cave chamber, Katies Bower, has two measurement points subject to different air-flow patterns, Lower Katies Bower at the base and Upper Katies Bower at the entrance – exit point in the main flow path. Flitch of Bacon is a 0.5 × 2.0 m narrow passage in the direct flow path.
Cavity ring down spectrometer (CRDS)
CO2, CH4, H2O and δ13C-in-CO2 were measured with a CRDS spectrometer (Picarro Inc., Santa Clara, CA, USA, model G1101i with CH4 cell upgrade, CFFDS-35) installed with a programmable multi port valve (MPV) sequencer for switching between 16 possible inlet ports. Sample air was drawn continuously through all sample lines from the 4 locations in Chifley Cave and 2 external locations Ports 1–6 by a Vacuubrand ME2 vacuum pump at 1–5 L/min, then passed to an exhaust line and vented outside the cave. The CRDS instrument draws sample at 25 ml/min via the MPV, which is connected by 1/8” Tee-union to the side of all 6 sample lines and also 7th reference gas line. Non-return valves were fitted to all sample lines to prevent possible back-flow from the lowest resistance path or shortest sample line. The MPV was programmed to analyse gas from each sample line sequentially after a dwell time of 10 minutes for cave air and 5 mins for external air. A reference gas instrument air cylinder was also measured in the hourly sequence. CRDS instrument raw data recorded at 8 sec intervals was averaged to 1 minute intervals and the first 2 minutes after port switching was discarded for cell flushing. Further data averaging reduced the data to one representative, 8 minute average, sample per hour. A manufacturer installed CRDS instrument data correction procedure for spectral CO2 and CH4 interferences was augmented by an additional empirical H2O correction after extensive testing at low CH4 concentrations and high H2O (details available on request). After averaging and post-processing H2O correction, data were calibrated. 5 calibration gases were analysed at the National Measurement Institute, by gas chromatograph with methanizer-FID detector. The composition was determined by comparison with primary gas standards prepared gravimetrically using a Sartorius CC10000S mass comparator, with buoyancy correction based on the cylinder volume and air density at the time of each measurement. 3 of the 5 calibration gases measured by NMI were commercially available zero air cylinders certified to contain less than 100 nmol/mol trace gas other than N2 and O2.
These three zero air gases measured by NMI reported 23, 39, 32 nmol/mol CH4, compared to a sample of summer air from Chifley Cave, Lower Katies Bower which contained 10 nmol/mol. Measurement error for low CH4 calibration gases measured NMI by gas chromatography was 5 nmol/mol. Repeated measurement (8 minute average) of the instrument air gas cylinder in the hourly cycle had a 1σ precision of 15 ppb. Summation of calibration gas error and CRDS instrument precision gives a CH4 measurement error of 20 ppb.
FTIR trace gas analyser
From March 2015 until the end of the experiment in July 2016 an FTIR trace gas analyser was deployed in parallel to the CRDS, sampling air from Upper and Lower Katies Bower, Flitch of Bacon and one external location through the same inlet lines in the same 1 hour cycle. The FTIR analyser was built at the University of Wollongong and is functionally similar to the Spectronus trace gas analyser (Ecotech Pty Ptd, Knoxfield, Vic Australia) and described in Griffith et al.36. The FTIR measured CO2 δ13C in CO2, CH4, CO and N2O in air with precision of the order of 0.1% for all species (1% for CO) and 1‰ for δ13C in CO2. The analyser was calibrated in the laboratory against four WMO-scale-traceable standards in the ambient air mole fraction ranges, and in the cave system against a single ambient level standard and against the high level standards described above for the CRDS.
The FTIR was also used to perform the in-cave chamber experiments. Air from the surface enclosures described in the main text was circulated in a closed loop between the surface enclosure and the FTIR, typically for 10–20 minutes alternating with flushing of the enclosure with ambient cave air for 10–20 minutes. Loss rates of CH4 in the enclosure were determined from the time-series of FTIR measurements. The flux calculations followed the methods given by Phillips et al.24.
In-cave sensors
Environmental sensors were placed above the 2.0 × 0.5 m passage-way at Flitch of Bacon (FOB) drip water site near Chifley Cave exit to Grand Arch. Data was recorded at 15 minute intervals on a Datataker DT80 Series 2 data logger. Air-flow direction and velocity data were obtained using a Gill Instruments Windsonic 2D sonic anemometer Option A. Air temperature data was obtained using a Vaisala HMP50 sensor prior to 24-Jul-13 and a Vaisala HMP60 sensor installed after 24-Jul-13. Carbon dioxide concentration was collected using a Vaisala GMM220 transmitter and Vaisala GMP222 carbon dioxide 0–7000ppm probes until 4-Dec-2013, with a data gap occurring between 11-Oct-13 to 21-Feb-14 due to a GMP222 probe failure and replacement. Radon concentration data was collected at 1 hour intervals in the cave using a Saphymo Alphaguard P30 radon monitor configured in diffusion mode from 29-Aug-2014 to 17-Aug-2015 and located adjacent to the Picarro air intake for lower Katies Bower.
Soil sensors
Soil sensors were installed on the surface of the hill above Chifley cave in the area surrounding the Plughole cave entrance. Soil sensor data was recorded at 15 minute sample intervals on Datataker DT82I Series 3 data logger. Soil CO2 concentration was obtained using a Vaisala GMM220 transmitter with a Vaisala GMP221 Carbon dioxide probe 0–3%. The GMP221 probe was installed in a 20 cm deep augered hole inside a Vaisala 211921GM in-soil adapter. Soil moisture and temperature data was obtained using a Stevens digital hydraprobe II installed in a separate augered hole 20 cm deep. Air temperature at the soil monitoring site was obtained using a Vaisala HMS82 intercap humidity and temperature transmitter in mini stevenson screen mounted at 2 m elevation on pole adjacent to soil probes. Radon concentration data was collected at 1 hour intervals using a Saphymo Alphaguard PQ2000 Pro radon monitor configured in flow mode. Air was pumped though the radon monitor at 1 L/min from a soil flux chamber located within 5 metres of the soil sensors.
Weather station
Weather data was collected using a Monitor Sensors weather station installed in forest 50 north of the plughole cave entrance. Data was logged using a Monitor Sensors SL5 uSmart data logger at 30 minute intervals until to 12-Mar-13, and hourly thereafter. Air temperature was obtained using a Monitor sensors uSmart TA1 ambient temperature sensor. Rainfall data was collected using a Monitor sensors uSmart RG2 0.2mm tipping bucket rain gauge (TBRG) until 20-Mar-2014 and a Hydrological Services TB4 0.2mm TBRG with onboard ML1-FL data logger after 29-May-2014.
Cave microbial diversity and quantification
Bacterial populations in cave sediments were classified by sequencing of the methane monooxygenase pmoA gene, using sediment DNA as template. Sediment samples (0–5 cm) were sampled aseptically, while swab samples were taken using a cotton swab dampened in sterile water then trimmed to fit into a 1.5 ml Eppendorf tube. Sediment and swab samples were stored at −20 °C until required, and total DNA was extracted using commercial kits (Mobio Powersoil), following the manufacturer’s instructions.
Methane monooxygenase diversity in the soil DNA was determined after amplification with the pmoA primers A189/A68237. Tag encoded-pyrosequencing analysis was done from the A189 primer, using Roche 454 FLX TitaniumTechnology (Research and Testing Laboratories, Lubbock, Texas). Raw sequence data was trimmed of low-quality bases using the Fungene pipeline38, and reads shorter than 150 base pairs were removed. Chimeric sequences were removed using USEARCH 6.0. Amino acid translations of sequences were obtained using FrameBot39, and sequences exhibiting less than 40% identity to a pmoA database (Fungene pipeline) were removed. Amino acid sequences were aligned using HMMER3 and phylogenetic trees were constructed in MEGA 5.240.
Total bacteria and methanotrophic bacteria in cave sediments were quantified by real-time PCR, using sediment DNA as template and determining the abundance of the 16 S rRNA gene and the methane monooxygenase pmoA gene respectively. 16 S abundance was determined using the 341 F/806 R primer set41, and individual pmoA clades were quantified using specific gene probes: A189/mb661 (pmoA)42; A189/Gam634r (USC-γ clade)23; A189/CL1603R (Cluster 1 clade)23. The quantification standards used were either purified amplicons from cave DNA, or a plasmid containing the pmoA gene of Methylococcus capsulatus Bath43.
Methanotrophic consumption of methane in cave sediments
Consumption of methane by sediments was measured by incubation of soil samples (5 g) in 120 ml serum bottles, corrected to a 10% (w/w) moisture content with deionized water. Methane was injected (1000 ppm), and the samples were incubated statically at room temperature. Methane disappearance was measured by gas chromatography on an HP-PLOT Q capillary column with helium as the carrier gas, using an oven temperature of 200 °C, and a flame ionisation detector. Dead controls were autoclaved prior to the initial methane injection.
Laboratory radon exposure experiments
Experiment 1
To study radon (ion) – air (CH4) interaction a 10 L gas bag (SKC Flexfoil) was infused with radon from a sealed source (PYLON 2000A) providing an initial radon concentration of 1.25MBq m−3 decaying to 0.70MBq m−3 after 3.2 days (AlphaGuard PQ2000).
Experiment 2
For methanotroph – air interaction we constructed a simple closed circuit loop; 1.6 L fixed volume reaction chamber with ~250 g Chifley Cave, Jenolan Caves, NSW sediment, and 5 L gas bag with air recirculated at 100 mL min−1. Experiment 2b includes sterilisation (60Co gamma, dose 25kGy). Methane was measured with a CRDS spectrometer (Picarro) and methanotrophs by qPCR (Supplementary methods).
Methane was measured with a CRDS spectrometer by removing approximately 200 ml air from the circuit for each measurement (Picarro Inc., Santa Clara, CA, USA, model G1101i CH4 cell upgrade, CH4 is water corrected & calibrated with 1σ precision, 20 ppb). Laboratory air was continuously measured for background methane concentration during the experiments.
An empirical proportional relationship between radon concentration and ion concentration (AlphaLab Air Ion Counter) in air was observed for a sealed 30 L container which approximates 1 Bq m−3 ≈ 10 ions cm−3. However, we noted significant variability (factor of ~5) of ion counter values with position, time (1–10 mins) and proximity of people ± static charge on clothing in laboratory measurements.
Three-dimensional cave mapping
Three-dimensional models of the caves44 were acquired using a novel handheld laser scanning system30. Path lengths between the entrances and various points in the caves were calculated by applying the A* path planning algorithm45 to a volumetric octree model derived from the laser data.
Data Availability
Additional full data for all plots in “Seasonal total methane depletion in limestone caves” is available; Waring, Chris L; Hankin, Stuart I; Griffith, David W T; Kettlewell, Graham (2017): Continuous 3-year record of cave methane, trace gases and environmental data collected in Jenolan Caves, Australia. PANGAEA, https://doi.org/10.1594/PANGAEA.878077 now available at the PANGAEA data repository.
Change history
11 April 2018
A correction to this article has been published and is linked from the HTML and PDF versions of this paper. The error has been fixed in the paper.
References
Waring, C. L., Griffith, D. W. T., Wilson, S. & Hurry, S. Cave atmosphere: A guide to calcification and a methane sink. Geochim Cosmochim Acta 73, A1419 (2009).
Mattey, D. P. et al. Methane in underground air in Gibraltar karst. Earth and Planetary Science Letters 374, 71–80 (2013).
Webster, K. D. et al. Methane dynamics in limestone caves. Geological Society of America Abstracts 45, (2013).
Fernandez-Cortes, A. et al. Subterranean atmospheres may act as daily methane sinks. Nature Communications 6, 7003 (2015).
Webster, K. D., Drobniak, A., Sauer, P. E., Mastalerz, M. & Schimmelmann, A. Cave air as a biosignature. 2nd International Planetary Caves Conference LPI Contribution No. 1883, 9009 (2015).
McDonough, L. K. et al. Spatial variability of cave-air carbon dioxide and methane concentrations and isotopic compositions in a semi-arid karst environment. Environmental Earth Sciences 75, 700 (2016).
Mattey, D. P. et al. Carbon dioxide, ground air and carbon cycling in Gibraltar karst. Geochimica et Cosmochimica Acta 184, 88–113 (2016).
Fairchild, I. J. et al. Modification and preservation of environmental signals in speleothems. Earth-Science Reviews 75, 105–153 (2006).
Spotl, C., Fairchild, I. J. & Tooth, A. F. Cave air control on dripwater geochemistry, Obir Caves (Austria): Implications for speleothem deposition in dynamically ventilated caves. Geochimica Et Cosmochimica Acta 69, 2451–2468 (2005).
Breecker, D. O. et al. The sources and sinks of CO2 in caves under mixed woodland and grassland vegetation. Geochimica et Cosmochimica Acta 96, 230–246 (2012).
James, E. W., Banner, J. L. & Hardt, B. A global model for cave ventilation and seasonal bias in speleothem paleoclimate records. Geochemistry, Geophysics, Geosystems 16, 1044–1051 (2015).
Cigna, A. A. Radon in Caves. International Journal of Speleology 34, 1–18 (2005).
Kowalczk, A. J. & Froelich, P. N. Cave air ventilation and CO2 outgassing by radon-222 modeling: How fast do caves breathe? Earth and Planetary Science Letters 289, 209–219 (2010).
Topp, E. & Pattey, E. Soils as sources and sinks for atmospheric methane. Canadian Journal of Soil Science 77, 167–177 (1997).
Le Mer, J. & Roger, P. Production, oxidation, emission and consumption of methane by soils: A review. European Journal of Soil Biology 37, 25–50 (2001).
Mook, W. G. Environmental isotopes in the hydrological cycle; Principles and applications. In: Introduction, Theory, Methods, Review. IHP-V. Technical Documents in Hydrology (ed Mook WG). IAEA-UNESCO (2000).
Hénault, C., Grossel, A., Mary, B., Roussel, M. & Léonard, J. Nitrous Oxide Emission by Agricultural Soils: A Review of Spatial and Temporal Variability for Mitigation. Pedosphere 22, 426–433 (2012).
Koehler, B. et al. An in-depth look into a tropical lowland forest soil: nitrogen-addition effects on the contents of N2O, CO2 and CH4 and N2O isotopic signatures down to 2-m depth. Biogeochemistry 111, 695–713 (2012).
Laubach, J., Barthel, M., Fraser, A., Hunt, J. E. & Griffith, D. W. T. Combining two complementary micrometeorological methods to measure CH4 and N2O fluxes over pasture. Biogeosciences 13, 1309–1327 (2016).
Ho, A. et al. Conceptualizing functional traits and ecological characteristics of methane-oxidizing bacteria as life strategies. Environmental Microbiology Reports 5, 335–345 (2013).
van Teeseling, M. C. F. et al. Expanding the Verrucomicrobial Methanotrophic World: Description of Three Novel Species of Methylacidimicrobium gen. nov. Appl Environ Microbiol 80, 6782–6791 (2014).
Knief, C. Diversity and Habitat Preferences of Cultivated and Uncultivated Aerobic Methanotrophic Bacteria Evaluated Based on pmoA as Molecular Marker. Frontiers in Microbiology 6, (2015).
Kolb, S., Knief, C., Dunfield, P. F. & Conrad, R. Abundance and activity of uncultured methanotrophic bacteria involved in the consumption of atmospheric methane in two forest soils. Environ Microbiol 7, 1150–1161 (2005).
Phillips, R. et al. Tracking Short-Term Effects of Nitrogen-15 Addition on Nitrous Oxide Fluxes Using Fourier-Transform Infrared Spectroscopy. Journal of Environmental Quality 42, 1327–1340 (2013).
Rayleigh, L. Theoretical considerations respecting the separation of gases by diffusion and similar processes. Philosophical Magazine Series 5 42, 493–498 (1896).
Templeton, A. S., Chu, K.-H., Alvarez-Cohen, L. & Conrad, M. E. Variable carbon isotope fractionation expressed by aerobic CH4-oxidizing bacteria. Geochimica et Cosmochimica Acta 70, 1739–1752 (2006).
Whiticar, M. J. Carbon and hydrogen isotope systematics of bacterial formation and oxidation of methane. Chemical Geology 161, 291–314 (1999).
Lennon, J. T. & Jones, S. E. Microbial seed banks: the ecological and evolutionary implications of dormancy. Nat Rev Micro 9, 119–130 (2011).
Lennon, J. T. et al. Microbial contributions to subterranean methane sinks. Geobiology 15, 254–258 (2017).
Zlot, R. & Bosse, M. Three-dimensional mobile mapping of caves. Journal of Cave and Karst Studies 76, 191–206 (2014).
Kirschke, S. et al. Three decades of global methane sources and sinks. Nature Geosci 6, 813–823 (2013).
Nisbet, E. G., Dlugokencky, E. J. & Bousquet, P. Methane on the Rise—Again. Science 343, 493–495 (2014).
Hutchens, E., Radajewski, S., Dumont, M. G., McDonald, I. R. & Murrell, J. C. Analysis of methanotrophic bacteria in Movile Cave by stable isotope probing. Environmental Microbiology 6, 111–120 (2004).
Sarbu, S. M., Kane, T. C. & Kinkle, B. K. A Chemoautotrophically Based Cave Ecosystem. Science 272, 1953–1955 (1996).
Tomczyk-Zak, K. & Zielenkiewicz, U. Microbial diversity in caves. Geomicrobiol J 33, 20–38 (2016).
Griffith, D. W. T. et al. A Fourier transform infrared trace gas analyser for atmospheric applications. Atmospheric Measurement Techniques 5, 2481–2498 (2012).
Holmes, A. J., Costello, A., Lidstrom, M. E. & Murrell, J. C. Evidence that particulate methane monooxygenase and ammonia monooxygenase may be evolutionarily related. FEMS Microbiol Lett 132, 203–208 (1995).
Fish, J. A et al. FunGene: the functional gene pipeline and repository. Front Microbiol 4, (2013).
Wang, Q. et al. Ecological patterns of nifH genes in four terrestrial climatic zones explored with targeted metagenomics using FrameBot, a new informatics tool. mBio 4, 9 (2013).
Tamura, K. et al. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28, 2731–2739 (2011).
Takahashi, S., Tomita, J., Nishioka, K., Hisada, T. & Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of bacteria and archaea using next-generation sequencing. PLOS One 9, 9 (2014).
Costello, A. M. & Lidstrom, M. E. Molecular characterization of functional and phylogenetic genes from natural populations of methanotrophs in lake sediments. Appl Environ Microbiol 65, 5066–5074 (1999).
Ly, M. A., Liew, E. F., Le, N. B. & Coleman, N. V. Construction and evaluation of pMycoFos, a fosmid shuttle vector for Mycobacterium spp. with inducible gene expression and copy number control. Journal of Microbiological Methods 86, 320–326 (2011).
Zlot, R. & Bosse, M. Jenolan Caves: 3D Data Collection. v1. CSIRO. Data Collection., (2015).
Nilsson, N. J. Principles of Artificial Intelligence. Springer-Verlag Berlin Heidelberg (1982).
Acknowledgements
We would like to acknowledge support from ANSTO Environment, University of Wollongong, Sydney University, Stephan Sestak CSIRO, Julia James and Alan Warild, Jenolan Caves guides Grant Commins, Dan Cove, Michael Collins, Geoff Melbourne, Peter, Ted, Alison, Anne, Scott and Jenolan Caves Reserve Trust management. We thank the reveiwers for their constructive comments.
Author information
Authors and Affiliations
Contributions
C.L.W. conceived and designed the study. S.I.H. and C.L.W. installed and operated all monitoring instruments, except FTIR, operated by D.W.T.G. and G.K. M.A.K. designed microbiological studies with field and laboratory measurements by V.K., N.L.W., and N.V.C. R.Z., M.B., and G.B. measured path length and area in Jenolan Caves. C.L.W., D.W.T.G., S.I.H. and M.A.K. wrote the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
41598_2017_7769_MOESM1_ESM.pdf
Supplementary Information; 2 text sections, Kinetics of radiolytic CH4 destruction and Assessment of potential mechanisms for seasonal CH4 depletion pattern, 7 Figures, and 4 Tables
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Waring, C.L., Hankin, S.I., Griffith, D.W.T. et al. Seasonal total methane depletion in limestone caves. Sci Rep 7, 8314 (2017). https://doi.org/10.1038/s41598-017-07769-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-07769-6
This article is cited by
-
Microbial tapestry of the Shulgan-Tash cave (Southern Ural, Russia): influences of environmental factors on the taxonomic composition of the cave biofilms
Environmental Microbiome (2023)
-
Environment-driven control of fungi in subterranean ecosystems: the case of La Garma Cave (northern Spain)
International Microbiology (2021)
-
Diversity of Polyketide Synthases and Nonribosomal Peptide Synthetases Revealed Through Metagenomic Analysis of a Deep Oligotrophic Cave
Microbial Ecology (2021)
-
Atmospheric Methane Oxidizers Are Dominated by Upland Soil Cluster Alpha in 20 Forest Soils of China
Microbial Ecology (2020)
-
Shotgun metagenomic sequencing from Manao-Pee cave, Thailand, reveals insight into the microbial community structure and its metabolic potential
BMC Microbiology (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
