
Abstract
Living organisms, ecosystems, and social systems are examples of complex systems in which robustness against inclusion of new elements is an essential feature. A recently proposed simple model has revealed a general mechanism by which such systems can become robust against inclusion of elements with totally random interactions when the elements have a moderate number of links. The interaction is, however, in many systems often intrinsically bidirectional like for mutual symbiosis and competition in ecology. This study reports the strong reinforcement effect of the bidirectionality of the interactions on the robustness of evolving systems. We show that the system with purely bidirectional interactions can grow with twofold average degree, in comparison with the purely unidirectional system. This drastic shift of the transition point comes from the reinforcement of each node, not from a change in structure of the emergent system. For systems with partially bidirectional interactions we find that the regime of the growing phase gets expanded. In the dense interaction regime, there exists an optimum proportion of bidirectional interactions for the growth rate at around 1/3. In the sparsely connected systems, small but finite fraction of bidirectional links can change the system’s behaviour from non-growing to growing.
Similar content being viewed by others


Introduction
An important and universal feature of many social, economical, ecological, and biological systems is that they are open. In these complex systems the constituting elements are not fixed and the complexity emerges or at least persists under successive appearances or introductions of new elements and disappearances or eliminations of old elements. Those systems sometimes grow or are stationary, but also some other times they collapse or go extinct. Hence one can ask a fundamental question why and when, in general, can such open and complex systems exist1,2,3,4,5,6,7,8,9,10,11,12,13.
In the previous studies, we have revisited this classical problem using a very simple graph dynamics model14. In this model, the system is composed of a collection of nodes connected by unidirectional links with weights. The nodes may represent various kinds of constitutive elements of the system i.e. chemicals, genes, animals, individuals, or species of some sort as we will term them from now on. Here the links are assumed to describe different types of directed influences between pairs of species. The strength of the influence of species j on species i is denoted by the weight of the unidirectional link from node j to node i, a ij . The weights can be either positive or negative. Each species has only one property that determines the dynamics, namely “fitness”, which is simply given by the sum of its incoming interactions from other species in the system, i.e., \({f}_{i}={\sum }_{j}^{in}{a}_{ij}\). A species can survive as long as its fitness is greater than zero; otherwise, it goes extinct. We calculate the fitness for each species and identify the species with minimum fitness and if it turns out to be non-positive, it will be deleted. Because each extinction event will also modify the fitness of the other species, the fitness is re-calculated and the least-fit species is re-identified. The deletion of species is continued until the minimum fitness value of a species becomes positive, meaning that nothing more will happen in terms of the above process. After finding such a state, we proceed to the next time step by adding a new species into the system. The m interacting species are chosen randomly from the resident species with equal probability and the directions are also determined randomly. The link weights are again assigned randomly using the standard normal distribution. Then, we re-calculate the fitness of each species to find the species that will become extinct.
The results of the model have shown that the repetition of this process gives rise either to continuous growth or stagnation for the system size, depending on the model’s unique parameter, namely the number of weighted unidirectional links per a new node, m. Furthermore it turns out that the system can grow only if the connections are moderately sparse, i.e. the average degree is within a rather narrow interval for 5 ≤ m ≤ 18. If the network becomes denser, there is a transition from growth to decay. It was shown that this transition originates from a balance between two effects, namely while on one hand the inclusion of species with more interactions makes each node more robust, on the other hand it also increases the impact of the loss of a node14, 15. This relation might be the origin of moderately sparse network structures ubiquitously found in real world cases, with the average degree of up to few tens16,17,18,19.
We should note that the approach of simple models we adopt here, for obtaining a new way to determine the system’s robustness, is quite different from the more standard and often used models of population dynamics. In our simple model, all the intrinsic dynamics of the systems is indirectly represented by the survival or fitness condition, i.e. f i > 0, which does not depend on the population of the species, as we have not introduced the population size as the degree of freedom. With the focus on robustness of the system while keeping the model general and “as simple as possible but not simpler”, we have ignored such details as the intensity of the flux of energy or nutrition (mass) in ecosystems, rates of chemical reactions, and trade volume in a company network, etc. Our aim with this general and simplified modelling approach is to study in addition to ecological systems even social and economic systems, in which it is hard to find precise equation of motion or evolution rule.
As a further support for this kind of universality seeking approach we mention that in several cases such modelling has been very successful. For example, one can find good agreement between a simple model and a real ecological system in the statistics such as the distribution of the lifetime of species20. Further examples of the power of a simplified modelling approach comes from looking neuronal and gene regulatory networks that have quite different types of elements considered as nodes, yet the model dynamics is the same for both of them with elements getting a shunt or off state going extinct and never getting excited. As a last justification worth mentioning is to regard the survival condition in our model as the condition for persistence in an original complex dynamical model. An interesting finding is that we can show a relatively natural example that the extinction condition in a population dynamics model with ratio-dependent predation term, proposed and studied in ecology to improve the model’s correspondence with reality21, reduces to the same population-independent form as is the case in our simple model22.
In the previous yet even simpler model, the interactions between species of the system were generated randomly with certain probability as unidirectional links between a pair of species i and j. This means that a random unidirectional link from j to i with a unidirectional link from i to j existing already, could be generated as a second order effect proportionally to its link density, as ~ρ 2. Hence the existence of influence from species i to j is independent of the existence of an influence in the opposite direction, i.e. from j to i. However, the simulation results of this model of self-emergence have shown that the accumulation of bidirectional influence is negligible, especially for large systems their density turned out to be vanishingly small.
However, in real systems, the interactions can on one hand be considered unidirectional as in case of reaction dynamics or signaling, and on the other hand bidirectional reflecting various types of bidirectional influence, like symbiosis or competition between species, predation-prey ecology, and even action-reaction law of Newtonian mechanics, etc. Therefore, in real systems we could distinguish five main types of interactions, of which two are unidirectional in nature, and three reflect bidirectional influence in terms of being cooperative, predatory, and competitive, as shown in Table 1 with the characterization and some real life examples.
Our aim in this study is to gain better understanding of the robustness and growth of real systems, with computational modeling. In this study we will focus on studying systems in which the interactions around the species newly introduced into a system can be unidirectional, bidirectional, or a mixture of both in certain proportions. In the Model section we revisit the previous model and extend it to include bidirectional influence between species. In the following Results section we describe the simulation results for the cases purely and partially bidirectional interactions. Next in the Discussion section we summarize our findings. This is then followed with the extensive Methods section.
Model
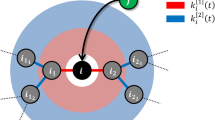
As in this study we are interested in the effects of bidirectional influences on the robustness of the evolving open system, we take the previous model as the base describing it first in detail and then extend it by introducing bi-directionality in a controlled way. In the previously model the evolution of an open system of species was set up to take place in a graph or network of randomly connected species or links describing the influences between species. This dynamical system evolves through interaction dependent survival of each species and a successive addition of new species to the system, as depicted in Fig. 1. This network consists of nodes (species) that are sparsely connected to each other with directed and (positively or negatively) weighted links, each describing the strength of interaction between a pair of species. In this system the survival of each species is determined by its fitness, which is calculated by the sum of the weights of its incoming links such that if the fitness of a given species is positive, it will survive. On the other hand if the fitness of a species is non-positive that species and all the interactions from and to it are deleted. If two or more species have non-positive fitness values, the species with the minimum fitness are deleted and the fitnesses of other species are recalculate. This so called extinction process is repeated until all the remaining species have positive fitness and once such a network structure is reached, nothing further will happen until a new node is born or introduced into the system. According to the terminology of ecology this is called a persistent state rather than a stable state.

The temporal evolution of a partially bidirectional model. (a) All the species have positive fitness and hence the system is in a persistent state. (b) The introduction of a new species (red). In this case the inclusion leads to the extinctions of a resident species (yellow) and that extinction triggers another extinction of a species, which is not directly connected to the newly introduced species (magenta). (c) Finally, the system reaches to a new persistent state. The numbers in the square brackets demonstrate the control parameters; M: the number of interacting species of the newly introduced species, P: the ratio of the bidirectional relation in the initially assigned interactions, and m = (1 + P)M: unidirectional link degree of the newly introduced species, and observables of the emerging network; N: the number of species, K: average number of interacting pairs per species in the system, Q: the ratio of the bidirectionally interacting pairs to the all interacting pairs in the system, and k = (1 + Q)K: average unidirectional link degree of the system.
When the system has relaxed to such a persistent state, a new species (node) is added to the system, and the above described process continues. Next M unidirectional links between the new node and the nodes randomly selected from the resident nodes are introduced.
Till this point our extended model is the same as the previous model, but instead of introducing unidirectional links a mixture in total of M unidirectional and bidirectional links is introduced by generating the bidirectional links with probability P and unidirectional links with probability 1 − P. The weight for each link is chosen again from a normal distribution. Thus our extended model has only two control parameters, M and P, instead of one in the previous model. In order to run the dynamics of both these models the above described addition and survival check process is repeated. A pseudo-code like description of the entire procedure of the present model is shown in the Methods section.
Counting each bidirectional link as two unidirectional links, one can say that m = (1 + P)M unidirectional links to the new species is introduced, which on average constitute PM bidirectional links. The extreme cases of P = 0 and P = 1 correspond to the original unidirectional model (with the number of unidirectional links per new species, m = M) and the pure bidirectional model (with m = 2M), respectively. We denote the density and the amount of bidirectionality of interactions in the emergent system in the same way, by the average number of interactions per species, i.e. K = ∑ i M i /N, where M i and N represent the number of links of i th resident and total number of resident species, respectively. We denote the ratio of bidirectional links by Q. Then the resident species has k = (1 + Q)K unidirectional links such that they form QK bidirectional links. The above definitions are summarized in Table 2.
Results
Purely bidirectional interaction (P = 1)
Transition in growth behavior at twofold critical point
We first examine the system with purely bidirectional interaction, i.e. we choose P = 1. Similarly with what was obtained for the model with unidirectional interactions, the purely bidirectional model yields two distinct phases characterized by its growth behavior. This is dependent on the unique parameter that specifies the number of interactions, M, we introduce to each new species (Fig. 2(a)). While the system keeps growing without any limitation to its size if M is smaller than a certain value M C (\({lim}_{t\to \infty }N(t)/t > 0\)), but the system shows finite size fluctuations if M > M C (\({lim}_{t\to \infty }N(t)/t=0\)). We call these phases diverging phase and finite phase, respectively.

The critical behavior of emergent systems with P = 0 or P = 1. (a) Temporal evolution of total number of species N(t) for P = 1. It diverges in time if the sum of in-degree and out-degree of each new species is small (m = 2M = 20 and 30), and it keeps fluctuating in a finite size for larger M (2M = 40 and 50). (b) Phase diagram of the growth behavior for P = 0 (red) and P = 1 (green). (top) Speed of divergence. (bottom) Average number of species. Filled symbols represent the systems with positive speed N(t)/t. The bidirectional model (circles) involves a transition from the diverging phase to the finite phase between m = 38 and 40. The critical point \({m}_{C}^{P=1}=2{M}_{C}^{P=1}=39\) is almost twofold of that of the unidirectional model (triangles) \({m}_{C}^{P\mathrm{=0}}=18.5\).
From the systematic simulations, we have estimated the critical number of bidirectional interactions to be at between M = 19 and 20. As in this case all the interactions are bidirectional, the critical point, measured by the number of unidirectional links, is \({m}_{C}^{P\mathrm{=1}}=2{M}_{C}^{P\mathrm{=1}}=39\) (Fig. 2(b)). This value is almost two times that of the pure unidirectional model (i.e. \({m}_{C}^{P\mathrm{=0}}=18.5\)). Another clear difference from the unidirectional model is the absence of a finite phase in the very sparse regime. In the unidirectional case, the systems with m ≤ 4 are in finite phase, due to the fragility of the emerging network. In contrast, in the bidirectional case, the system even with M = 1 turns out to exhibit diverging behavior. This is basically caused by the easiness of forming mutually supporting “dimers”, which is enough to let the nodes survive even when they are not globally connected. This regime will be discussed later in more detail. Our primary aim in what follows is to understand the twofold transition point in the well-connected regime.
To illustrate why we measure the interaction density by the number of unidirectional links and call the transition point “twofold”, we will briefly review the mean field argument on the basis of which the understanding and the rough prediction of the transition in the unidirectional model was obtained14, 23. In a system the introduction of a new species can lead to the extinctions of the resident species. We denote the extinction probabilities during this event as E i . It should be noted that the extinction of resident species can also cause a cascading extinction through the deletion of the links from it, with a certain probability E e . Assuming a structure-less random network topology, which is supported by the direct observation of the emergent network, the average number of extinctions N E per a newly introduced species can be calculated as a simple infinite geometric series as follows
for the unidirectional model, and,
for the bidirectional model. Here the critical point corresponds to N E = 1. Substituting E i and E e by an average extinction probability E and k by m, we obtain a simple guide of the transition point, namely mE = 1. Therefore we expect \({m}_{C}^{P\mathrm{=1}}E=2{M}_{C}^{P\mathrm{=1}}E=1\) for the bidirectional model, which means that the critical point of the bidirectional model should have the same number of unidirectional links \({m}_{C}^{P\mathrm{=1}}=2{M}_{C}^{P\mathrm{=1}}\) with that of the unidirectional model \({m}_{C}^{P\mathrm{=0}}=18.5\) 14. The details and an enhanced version of this mean-field argument are shown in the Methods section.
Reinforcement of elements
In order to clarify the crucial factor for the shift of the transition point, we evaluate the transition point using Eq (1) and Eq (2) with empirically obtained extinction probabilities (see the Method section). The average number of total extinctions per a newly introduced species increases monotonically as a function of the degree of a new species both for the unidirectional model and for the bidirectional model. The quantities \({N}_{E}^{P\mathrm{=0}}\) and \({N}_{E}^{P\mathrm{=1}}\) cross the critical value 1 at m * ~ 19.5 for the unidirectional model and 2M * ~ 39 for the bidirectional model, respectively, as depicted in Fig. 3(a). These critical points m * and 2M * agree well with the real transition points, \({m}_{C}^{P\mathrm{=0}}=18.5\) and \(2{M}_{C}^{P\mathrm{=1}}=39\).

The mean-field approximation predictions of the transition points. (a) Estimation of the transition point from the diverging phase to the finite phase. The average number of total extinctions, \({N}_{E}^{P\mathrm{=0}}\) (circles) and \({N}_{E}^{P\mathrm{=1}}\) (boxes), increases monotonically as a function of m. They cross the critical value 1 at m * = 19.5 for the unidirectional model and at 2M * = 39 for the bidirectional model. Both of m * and 2M * agree well with the real transition points \({m}_{C}^{P\mathrm{=0}}=18.5\) and \(2{M}_{C}^{P\mathrm{=1}}=39\). (b) Extinction probability E i . (c) Extinction probability E e . For the unidirectional model (circles) and the bidirectional model (boxes) with the same unidirectional link degree of a newly incoming species m, the extinction probabilities E i and E e of the bidirectional model are smaller than the corresponding probabilities of the unidirectional model.
This result suggests that the shift of the transition point stems from the decrease in the extinction probability of the emergent system. In Fig. 3(b) we see that the quantities E i and E e decrease monotonically as a function of the degree of a new species both for the unidirectional and bidirectional models. Comparing the bidirectional model with the unidirectional model, however, the bidirectional model gives a smaller E i and E e if the new species of the unidirectional model and the bidirectional model are assigned same number of unidirectional links, m P=0 = 2M P=1.
Further investigation of the emerging system reveals that the larger decrease in the extinction probability of the bidirectional model is caused by the slight increase in the average degree, together with the small change in the link weight correlation (see Method section for more details).
Model with partially bidirectional interaction (P ∈ [0, 1])
Bidirectionality dependence of the transition of the growth behavior
Next, we examine the systems with partially bidirectional interactions to investigate the influence of the proportion of bidirectionality P on the growth behavior. We find that P ∈ (0, 1) expands the area of the diverging phase, as seen in Fig. 4(a). Interestingly or even surprisingly, the system with a certain level of bidirectionality steadily grows even if the system with purely unidirectional interactions (P = 0) or purely bidirectional interactions (P = 1) is in the finite phase. There are two phase boundaries, one found at around 18 < M < 22 and another at around 1 ≤ M < 5, as can be seen in Fig. 4(a). Here the finite phase remains not only in the dense interaction regime and but also in the sparse interaction regime.

The phase diagram of the growth behavior for (a) 1 ≤ M ≤ 22, (b) the dense interaction regime 18 ≤ M ≤ 22, and (c) the sparse interaction regime 1 ≤ M ≤ 5. The partial bidirectionality expands the area of the diverging phase (filled circles). There exists an upper phase boundary (dotted line) at around 18 < M < 22 and a lower phase boundary (solid line) at around 1 ≤ M < 5. The upper boundary is asymmetric for the bidirectionality and there is an optimum bidirectionality for the growth rate at around P ~ 0.3. The finite phase (open circles) in the sparse connecting regime M < 5 remains within a range of small but finite bidirectionality. The area of the finite phase gradually decreases as a function of the increasing bidirectionality.
One may expect a strong effect of birectionality, because strong accumulation and cooperative interaction is possible. The most prominent example of this is the case of M = 1, in which each cooperative dimer can survive independently letting the system to consist of many independent clusters with broad size distribution. However, in the dense regime, one cannot find such a strong structuring effect. For example, in the dense regime, the proportion of bidirectional links in the emerging system Q remains almost the same as the initial proportion P (see Methods).
Growth behavior in the dense interaction regime (19 ≤ M ≤ 22)
Figure 5(a) shows the speed of divergence in the dense interaction regime. Here the speed of divergence is asymmetrically distributed as a function of a proportion of the bidirectional links. It is seen that there is an optimum bidirectionality for the growth rate P ~ 1/3. Thus, the upper phase boundary in the denser interaction regime (18 < M < 22) is asymmetric for the proportion of the bidirectional links (Fig. 4(b)). We also find that there exists an optimum bidirectionality for the growth rate at around P ~ 1/3.

The growth behavior of the partially bidirectional systems in (a) the dense interaction regime and (b) the sparse interaction regime. (a) Moderate proportion of bidirectional links brings the system from the finite phase to the diverging phase. The speed of divergence is asymmetric as a function of the bidirectionality and there exists an optimum bidirectionality for the growth rate at around P ~ 1/3. (b) The speed of divergence increases monotonically from zero as a function of proportion of the bidirectional links. A small but finite bidirectionality brings the sparsely connected system from the finite phase to the diverging phase.
The system with a moderate amount of bidirectionality grows for M = 19, 20, and 21, even if the system with purely unidirectional interactions or purely bidirectional interactions (P = 0 or P = 1) is in the finite phase. In addition, a small amount of bidirectionality makes the partially bidirectional system grow. On the other hand, the system with M = 22 remains in the finite phase only for all P ∈ [0, 1].
Growth behavior in the sparse interaction regime (1 ≤ M ≤ 4)
In Fig. 5(b) it is seen that the speed of divergence increases monotonically from zero with the increasing proportion of the bidirectional links. Thus, a small but finite amount of bidirectionality brings the system from the original finite phase to the diverging phase, as is evident in Fig. 4(c). The area of the finite phase gradually decreases as a function of the amount of bidirectionality. Then if the proportion P of the bidirectionality is set larger than a certain value, the sparsely connected system involves only a diverging phase. We can also observe that, in this sparse regime, the emergent systems tend to have larger degree and larger proportion of bidirectional links than the input values (〈K〉 > M and 〈Q〉 > P, see Table 3 in the Method section). This indicates relatively strong self-organization that is regarded as the main reason for the change in the system’s growth behavior such as the disappearance of the finite phase.
Discussion
We have studied the effects of introducing new species with variable degree of bidirectionality in the interactions between species, focusing especially on the dynamics of the robustness of the network of species.
First we found that the system with purely bidirectional interactions is more robust against the inclusion of new species, which is characterized by the twofold transition point from the growing phase to the finite phase. The twofold transition point also means that one can roughly identify or even mix the unidirectional and bidirectional interactions to apply our framework to real systems, in which the detailed information of interactions and their nature is often not available. The upper limit of the average number of interacting species per species for letting the system to grow its diversity is around 20 irrespective of the bidirectionality of interaction. This is again confirmed to be significantly looser than the classical diversity-stability relation known for random dynamical systems1 and for generalized Lotka-Volterra system with random inclusion of new species24, 25. This limiting condition seems to be more consistent with the persistence condition of ecological models with realistic nonlinear interactions8 and of a model with spatial structure26, but most importantly with empirical data19.
In the model with partial bidirectionality, we find that there exists an optimum ratio of bidirectional interactions at around 1/3, for the growth of the system to take place. Because the bidirectionality of the emergent system remains near to its input proportion, this optimum in the middle seems to be consistent with the enhancement of the system by the diversity of interaction types in the system, found in a more complex models of ecosystems27. We also find that only a small, though finite, portion of bidirectional interactions let the very sparse (M ≤ 4) system to grow to a large and connected structure.
Except for the very sparse regime, all those drastic changes in the entire system’s robustness are caused by a little change in the degree and also in the emergent correlations between the link weights. The network characteristics such as clustering and assortativity are also found to be not much deviated from that of Erdös-Rényi random graph (see Methods). This is quite noteworthy because the system in our model self-emerges without any apparent constraints and hence more drastic self-organization could be possible. Indeed, stronger self-organization and its effect on the growth behavior of the system is observed in the sparse regime (M ≤ 4).
A study of an extended model in which we force the system to have stronger correlations in the link and weight topologies could be the next step6, 26, 28, 29, to further relate our framework to the real systems.
Methods
A pseudo-code representation of the model
The present model is expressed as the following pseudo-code
-
0.
(Create an initial network.)
-
1.
Calculate the fitness for each species: \({f}_{i}={\sum }_{j}^{incoming}{a}_{ij}\)
-
2.
If the fitness of the species are all positive (Fig. 1(a) and (c), persistent state), go to step 3. If not,
-
(i)
Delete the species with minimum (and non-positive) fitness value.
-
(ii)
Delete the links connecting to and from that species.
-
(iii)
Go back to the step 1 (to re-evaluate the fitnesses).
-
(i)
-
3.
A new species is added to the system (Fig. 1(b)).
-
(i)
M interacting species are chosen randomly from the resident species, with equal probability 1/N(t).
-
(ii)
The new species forms bidirectional links, with probability P.
-
(iii)
With probability 1 − P, we assign a unidirectional link. Its direction is randomly chosen with probability 1/2.
-
(iv)
The link weights are independently drawn from the standard normal distribution. The link weights between incoming and outgoing unidirectional links which consist up one bidirectional link are also independent: 〈a ij a ji 〉 = 0.
-
(v)
Go to 1.
-
(i)
The mean-field estimation of the transition point
In the original unidirectional model (P = 0), the system keeps growing only if the interaction is moderately sparse (5 ≤ m ≤ 18) and cannot grow outside of this regime (m ≤ 4 and m ≥ 19). The basic mechanism of the transition from the diverging phase to the finite phase at m c = 18.5 is known to be explained by the mean-field argument14, 15, which assumes a correlation-less random network structure for the interaction network of self-emerging system. The validity of this assumption is confirmed in the simulation, from the direct observation of the system. With such an assumption, the system’s growth behaviour against the inclusions of new species is characterized by two average extinction probabilities, E i and E e . E i is the extinction probability of the resident species after obtaining a direct incoming link from the newly introduced species. The other extinction probability, E e , is the probability of a species to get extinct after loosing one incoming link which is brought by the extinction of the neighboring species. Because of the assumption of random network topology, the total number of extinctions of the resident species per newly introduced species 〈N e 〉 is calculated from a simple geometric sum as
where mE i /2 is the average number of extinctions directly caused by the newly introduced species and the infinite geometric sum corresponds to the process of cascade extinctions. The transition point is the point at which 〈N E 〉 = 1, i.e. the average number of gain and loss of the species are equal.
Let us perform the same calculation in the system which consists of fully bidirectional interactions (P = 1). In this case, the expected number of extinctions in the layer of direct connection to the newly introduced species is ME i . These extinctions cause the following link-deletion events, subsequently giving rise to KE e extinctions in the next-nearest neighbors. After this layer, the contribution of the cascading extinction in each deeper layer is (K − 1)E e . Therefore, the total number of extinctions per a newly introduced species in the bidirectional model is calculated as,
For the simple evaluation of the transition point, we further assume that the average degree of the emergent system is same as the initial degree, i.e. m = k and M = K. Because the correlation-less network assumption it also means that the extinction probability is a function of the unique system parameter, i.e. average degree, meaning that it can be written as a function of initial degree m = 2M. Based on the empirical findings, we also assume that the extinction probabilities are similar and much smaller than 1 near the transition point (\({E}_{c}={E}_{i}={E}_{e}\ll 1\)). Then the condition for the unidirectional case (Eq. 3) becomes the following very simple form14
and the corresponding simple form for the bidirectional system (Eq. 4) is
These results gives us the basic estimation for the transition point for the bidirectional systems, which says that the transition occurs at the same critical unidirectional degree:
This is why we regard the observed transition point, \({M}_{c}^{P\mathrm{=1}} \sim {m}_{c}^{P\mathrm{=0}}\), “tow-fold” of what is naively expected.
Calculation of the extinction probabilities in the emerging systems
The extinction probabilities E i and E e in an emerging system are calculated according to those definitions as,
where \({\#}_{{a}_{ij}\le {f}_{i}}\) and # link represent the number of species with smaller fitness than the link weight receiving from each link and the total number of links of the emergent system, respectively.
The network structure and its effect in the bidirectional model (P = 1)
Degree distribution
As shown in the Fig. 6(a), the average unidirectional link degree of the emergent system k increases almost linearly as a function of the degree of a new species m, both for the unidirectional model and for the bidirectional model. For the unidirectional model and the bidirectional model with the same degree of a newly incoming species m P=0 = 2M P=1, the average degree of the bidirectional model is slightly larger than that of the unidirectional model. This difference comes mainly from the broader tail of degree distribution in the bidirectional case. The peak position of the degree distribution at k = m and the tail on the large degree side is fatter than that on the other side (Fig. 6(b)).

(a) Average degree of the emergent system. The emergent system in the bidirectional model (boxes) has larger average degree than that of the unidirectional model (circles) with the same unidirectional link degree of the new species m P=0 = 2M P=1. (b) Degree distribution of the emergent system at m = 18. The degree distribution of the bidirectional model (solid line) has broader tails than that of the unidirectional model (dashed line), with sharing the same peak position.
One can find a scaling relation in the fitness distributions of the unidirectional model and the bidirectional model (Fig. 7(a)). The fitness distributions of the unidirectional model and the bidirectional model match well when the control parameter of those satisfies the relation: m P=0 = 2(M P=1 + 1). This scaling relation holds also for the distributions of the link weights (Fig. 7(b)). All the results above can be explained by the slight increase in the actual average degree in the emergent system in the bidirectional case: k P=1(M) = k P=0(2(M + 1)).

Scaling relation in the fitness distribution function. (a) Fitness distributions of the unidirectional model (m = 14, 18, 22, and 30) and of the bidirectional model (M = 6, 8, 10, and 14). (b) Link weight distributions of the unidirectional model with m P=0 = 18 and of the bidirectional model with M P=1 = 8. Solid blue line is a fitting function, a standard normal distribution function G(x, μ) (solid line) with its mean μ = 0.45.
Weak correlation in link weights
According to the scaling relation, the unidirectional model and the bidirectional model give the same average degree of the emergent systems k P=0 = 2K P=1 at m P=0 = 2(M P=1 + 1). Therefore, the unidirectional model and the bidirectional model are expected to have the same extinction probabilities at k P=0 = 2K P=1 (Fig. 8(a)). However, while E i is found to obey this relation, E e of the bidirectional model is slightly smaller than that of the unidirectional model (Fig. 8(b)). It suggests that the system of the bidirectional model has stronger weight-fitness correlation than the unidirectional model.

(a) Extinction probability E i . (b) Extinction probability E e . Emergent systems of the unidirectional and bidirectional models with same average unidirectional link degree show the same extinction probability against the direct disturbance event from the new species, E i . On the other hand, the extinction probability against the loss of a link, E e , is slightly smaller for the bidirectional model than that of the unidirectional model.
Here we conclude that the shift of the transition point stems from the small decrease in the extinction probability of the bidirectional model due to the weak shift in the average degree and the correlation in link weights, not from strong structuring or emergence of a certain motif in the emergent system.
The network structure of the partially bidirectional model
Dense regime
The self-emerging network of the partially bidirectional model in the dense regime is found to have only weak structuring, similar to the fully bidirectional case. The proportion of bidirectional links in the system also remains the same as the newly introduced interactions (Q = P).
Sparse regime
In the sparse regime, the emergent systems tend to have larger degree and larger proportion of bidirectional links than input values (〈K〉 > M and 〈Q〉 > P, see Table 3). This indicates the relatively strong self-organization effect in this regime, which is regarded as the main reason for the disappearance of the finite phase.
In order to investigate the network structure in this regime, we analyze the network size distribution. As shown in Fig. 9(a), the systems are composed of multiple clusters in the diverging phase, whereas all resident species construct a single big cluster in the finite phase. In the diverging phase, in contrast, the systems with M = 2, 3 and 4 are composed of a single big cluster with some small fragments such as dimers, trimers, etc. (Fig. 9(b)). The cluster size distribution of the system with M = 1 is continuous up to its largest cluster size, meaning that there is no dominant giant component.

(a) Average number of clusters of the emergent systems at t = 9.5 × 106. Except for the case of M = 1, The systems in the finite phase (open symbols) have single cluster, i.e., those are connected. The systems in the diverging phase (filled symbols) consist of multiple independent clusters. (b) Cluster size distributions of the systems with purely bidirectional interactions in the diverging phase, at the same time step (t = 9.5 × 106). In the case of M = 1 (open reserve triangles), the distribution is continuous and broad up to the largest cluster size. Systems with M > 1 are composed of a single big cluster and some small fragments.
The basic mechanism of the self-organization seems to be a formation of cooperative dimers. Such a dimer can survive independently and hence can be a nuclei of non-connecting cluster. We can see this effect in the cluster size distribution clearly in the case of M = 1. Although it becomes less visible in the cluster size distribution in the case of M > 1, because of the random reconnection (or, bridging) process due to the introduction of new nodes, the dimer nucleus still let the network, which consists of sparsely connected almost independent clusters, survive.
Assortativity, Nestedness, and Clustering
Some network characteristics of the emergent systems are shown in Table 3. For this we treat unidirectional and bidirectional links equally as directionless links. Therefore the assortativity coefficient is calculated from the directionless link degrees of the nodes in the both ends of link i, \({\kappa }_{i}^{\alpha }\) and \({\kappa }_{i}^{\beta }\), as follows
Moreover, we adopt a nestedness measure for uniparpite and dilectionless networks30 as
where a ij and K i represent the adjacency matrix element and the degree of species i, defined by the directionless links. The clustering coefficient is also calculated from the number of triples and triangles formed by the directionless links as,
Comparing to the Erdös-Rényi random graph, clustering coefficient is slightly smaller and nestedness is almost the same. In addition the assortativity coefficient turns out to be slightly negative.
References
Gardner, M. R. & Ashby, W. R. Connectance of large dynamic (cybernetic) systems: critical values for stability. Nature 228, 784–784, doi:10.1038/228784a0 (1970).
Pimm, S. L. Complexity and stability: another look at macarthur’s original hypothesis. OIKOS 33, 351–357, doi:10.2307/3544322 (1979).
Bak, P. & Sneppen, K. Punctuated equilibrium and criticality in a simple model of evolution. Phys. Rev. Lett. 71, 4083–4086, doi:10.1103/PhysRevLett.71.4083 (1993).
Tokita, K. & Yasutomi, A. Mass extinction in a dynamical system of evolution with variable dimension. Phys. Rev. E 60, 842–847, doi:10.1103/PhysRevE.60.842 (1999).
Albert, R., Jeong, H. & Barabási, A.-L. Error and attack tolerance of complex networks. Nature 406, 378–382, doi:10.1038/35019019 (2000).
Christensen, K., di Collobiano, S. A., Hall, M. & Jensen, H. J. Tangled nature: a model of evolutionary ecology. J. theor. Biol. 216, 73–84, doi:10.1006/jtbi.2002.2530 (2002).
Jain, S. & Krishna, S. Large extinctions in an evolutionary model: The role of innovation and keystone species of innovation and keystone species. Proc. Natl. Acad. Sci. USA 99, 2055–2060, doi:10.1073/pnas.032618499 (2002).
Kondoh, M. Foraging adaptation and the relationship between food-web complexity and stability. Science 299, 1388–1391, doi:10.1126/science.1079154 (2003).
Moreira, A. A. How to make a fragile network robust and vice versa. Phys. Rev. Lett. 102, 018701, doi:10.1103/PhysRevLett.102.018701 (2009).
Perotti, J. I. Emergent self-organized complex network topology out of stability constraints. Phys. Rev. Lett. 103, 108701, doi:10.1103/PhysRevLett.103.108701 (2009).
Buldyrev, S. V., Parshani, R., Paul, G., Stanley, H. E. & Havlin, S. Catastrophic cascade of failures in interdependent networks. Nature 464, 1025–1028, doi:10.1038/nature08932 (2010).
Stollmeier, F. Possible origin of stagnation and variability of earth’s biodiversity. Phys. Rev. Lett. 112, 228101, doi:10.1103/PhysRevLett.112.228101 (2014).
Watanabe, A., Mizutaka, S. & Yakubo, K. Fractal and small-world networks formed by self-organized critical dynamics. Journal of the Physical Society of Japan 84, 114003, doi:10.7566/JPSJ.84.114003 (2015).
Shimada, T. A universal transition in the robustness of evolving open systems. Scientific Reports 4, 4082, doi:10.1038/srep04082 (2014).
Shimada, T. A Universal Mechanism of Determining the Robustness of Evolving Systems in Mathematical Approaches to Biological Systems, (ed. Ohira T. and Uzawa T.) 95–117, doi:10.1007/978-4-431-55444-8_5 (Springer, 2015).
Winemiller, K. O. Must connectance decrease with species richness? The American Naturalist 134, 960–968, doi:10.1086/285024 (1989).
Albert, R. & Barabási, A.-L. Statistical mechanics of complex networks. Rev. Mod. Phys. 74, 47–97, doi:10.1103/RevModPhys.74.47 (2002).
Ma, S., Gong, Q. & Bohnert, H. J. An arabidopsis gene network based on the graphical gaussian model. Genome Research 17, 1614–1625, doi:10.1101/gr.6911207 (2007).
Ings, T. C. et al. Review: Ecological networks–beyond food webs. Journal of Animal Ecology 78, 253–269, doi:10.1111/j.1365-2656.2008.01460.x (2009).
Murase, Y., Shimada, T. & Ito, N. A simple model for skewed species-lifetime distributions. New J. of Physics 12, 063021, doi:10.1088/1367-2630/12/6/063021 (2010).
Arditi, R. & Ginzburg, L. R. Coupling in predator-prey dynamics: Ratio-dependence. Journal of Theoretical Biology 139, 311–326, doi:10.1016/S0022-5193(89)80211-5 (1989).
Shimada, T., Murase, Y. & Ito, N. Do Connections Make Systems Robust? A New Scenario for the Complexity-Stability Relation. Proceedings of the International Conference on Social Modeling an Simulation, plus Econophysics Colloquium 2014, 99–109, doi:10.1007/978-3-319-20591-5_9 (2015).
Shimada, T. & Ogushi, F. A mean-field analysis of the simple model of evolving open systems. Journal of Physics: Conference Series 750, 012008, doi:10.1088/1742-6596/750/1/012008 (2016).
Taylor, P. J. Consistent scaling and parameter choice for linear and generalized lotka-volterra models used in community ecology. J. theor. Biol. 135, 543–568, doi:10.1016/S0022-5193(88)80275-3 (1988).
Taylor, P. J. The construction and turnover of complex community models having generalized lotka-volterra dynamics. J. theor. Biol. 135, 569–588, doi:10.1016/S0022-5193(88)80276-5 (1988).
Mathiesen, J., Mitarai, N., Sneppen, K. & Trusina, A. Ecosystems with mutually exclusive interactions self-organize to a state of high diversity. Phys. Rev. Lett. 107, 188101, doi:10.1103/PhysRevLett.107.188101 (2011).
Mougi, A. & Kondoh, M. Diversity of interaction types and ecological community stability. Science 337, 349–351, doi:10.1126/science.1220529 (2012).
Coyte, K. Z., Schluter, J. & Foster, K. R. The ecology of the microbiome: Networks, competition, and stability. Science 350, 663–666, doi:10.1126/science.aad2602 (2015).
Yoshida, K. & Tokita, K. Properties of ecosystems that are vulnerable during eco-fusion. Scientific Reports 5, 7939, doi:10.1038/srep07939 (2015).
Lee, D.-S., Maeng, S. E. & Lee, J. W. Scaling of nestedness in complex networks. Journal of the Korean Physical Society 60, 648–656, doi:10.3938/jkps.60.648 (2012).
Acknowledgements
F.O. was partly supported by “Materials Research by Information Integration” Initiative (MI2I) project of the Support Program for Starting Up Innovation Hub from the Japan Science and Technology Agency (JST). K.K. acknowledges financial support by the Academy of Finland Research project (COSDYN) No. 276439 and EU HORIZON 2020 FET Open RIA project (IBSEN) No. 662725. JK was partially supported by H2020 FETPROACT-GSS CIMPLEX Grant No. 641191. T.S. was partly supported by JSPS KAKENHI Grant Number 15K05202 and CREST, JST.
Author information
Authors and Affiliations
Contributions
F.O. and T.S. conceived the model and conducted the simulation. All authors analysed the results and wrote the manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ogushi, F., Kertész, J., Kaski, K. et al. Enhanced robustness of evolving open systems by the bidirectionality of interactions between elements. Sci Rep 7, 6978 (2017). https://doi.org/10.1038/s41598-017-07283-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-07283-9
This article is cited by
-
Motif dynamics in signed directional complex networks
Journal of the Korean Physical Society (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
