
Abstract
Sexual dimorphism arises from genetic differences between male and female cells, and from systemic hormonal differences1,2,3. How sex hormones affect non-reproductive organs is poorly understood, yet highly relevant to health given the sex-biased incidence of many diseases4. Here we report that steroid signalling in Drosophila from the ovaries to the gut promotes growth of the intestine specifically in mated females, and enhances their reproductive output. The active ovaries of the fly produce the steroid hormone ecdysone, which stimulates the division and expansion of intestinal stem cells in two distinct proliferative phases via the steroid receptors EcR and Usp and their downstream targets Broad, Eip75B and Hr3. Although ecdysone-dependent growth of the female gut augments fecundity, the more active and more numerous intestinal stem cells also increase female susceptibility to age-dependent gut dysplasia and tumorigenesis, thus potentially reducing lifespan. This work highlights the trade-offs in fitness traits that occur when inter-organ signalling alters stem-cell behaviour to optimize organ size.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others


Data availability
Source data are provided with this paper.
Code availability
Code for all FIJI macros used in this study is available for download via the Supplementary Information. These macros are available as Supplementary Data 1–6.
References
Capel, B. Vertebrate sex determination: evolutionary plasticity of a fundamental switch. Nat. Rev. Genet. 18, 675–689 (2017).
Hudry, B., Khadayate, S. & Miguel-Aliaga, I. The sexual identity of adult intestinal stem cells controls organ size and plasticity. Nature 530, 344–348 (2016).
Sieber, M. H. & Spradling, A. C. Steroid signaling establishes a female metabolic state and regulates SREBP to control oocyte lipid accumulation. Curr. Biol. 25, 993–1004 (2015).
Ober, C., Loisel, D. A. & Gilad, Y. Sex-specific genetic architecture of human disease. Nat. Rev. Genet. 9, 911–922 (2008).
Schwedes, C. C. & Carney, G. E. Ecdysone signaling in adult Drosophila melanogaster. J. Insect Physiol. 58, 293–302 (2012).
Lavrynenko, O. et al. The ecdysteroidome of Drosophila: influence of diet and development. Development 142, 3758–3768 (2015).
Ameku, T. & Niwa, R. Mating-induced increase in germline stem cells via the neuroendocrine system in female Drosophila. PLoS Genet. 12, e1006123 (2016).
Meiselman, M. et al. Endocrine network essential for reproductive success in Drosophila melanogaster. Proc. Natl Acad. Sci. USA 114, E3849–E3858 (2017).
Simon, A. F., Shih, C., Mack, A. & Benzer, S. Steroid control of longevity in Drosophila melanogaster. Science 299, 1407–1410 (2003).
Tricoire, H. et al. The steroid hormone receptor EcR finely modulates Drosophila lifespan during adulthood in a sex-specific manner. Mech. Ageing Dev. 130, 547–552 (2009).
Jiang, H. et al. Cytokine/Jak/Stat signaling mediates regeneration and homeostasis in the Drosophila midgut. Cell 137, 1343–1355 (2009).
Regan, J. C. et al. Sex difference in pathology of the ageing gut mediates the greater response of female lifespan to dietary restriction. eLife 5, e10956 (2016).
Jiang, H., Grenley, M. O., Bravo, M. J., Blumhagen, R. Z. & Edgar, B. A. EGFR/Ras/MAPK signaling mediates adult midgut epithelial homeostasis and regeneration in Drosophila. Cell Stem Cell 8, 84–95 (2011).
Zhang, P. et al. An SH3PX1-dependent endocytosis-autophagy network restrains intestinal stem cell proliferation by counteracting EGFR-ERK signaling. Dev. Cell 49, 574–589 (2019).
Reiff, T. et al. Endocrine remodelling of the adult intestine sustains reproduction in Drosophila. eLife 4, e06930 (2015).
Ono, H. et al. Spook and Spookier code for stage-specific components of the ecdysone biosynthetic pathway in Diptera. Dev. Biol. 298, 555–570 (2006).
Marvin, K. A. et al. Nuclear receptors Homo sapiens Rev-erbβ and Drosophila melanogaster E75 are thiolate-ligated heme proteins which undergo redox-mediated ligand switching and bind CO and NO. Biochemistry 48, 7056–7071 (2009).
White, K. P., Hurban, P., Watanabe, T. & Hogness, D. S. Coordination of Drosophila metamorphosis by two ecdysone-induced nuclear receptors. Science 276, 114–117 (1997).
Biteau, B., Hochmuth, C. E. & Jasper, H. JNK activity in somatic stem cells causes loss of tissue homeostasis in the aging Drosophila gut. Cell Stem Cell 3, 442–455 (2008).
Biteau, B. et al. Lifespan extension by preserving proliferative homeostasis in Drosophila. PLoS Genet. 6, e1001159 (2010).
Patel, P. H., Dutta, D. & Edgar, B. A. Niche appropriation by Drosophila intestinal stem cell tumours. Nat. Cell Biol. 17, 1182–1192 (2015).
O’Brien, D. M., Min, K.-J., Larsen, T. & Tatar, M. Use of stable isotopes to examine how dietary restriction extends Drosophila lifespan. Curr. Biol. 18, R155–R156 (2008).
Speakman, J. R. The physiological costs of reproduction in small mammals. Phil. Trans. R. Soc. Lond. B 363, 375–398 (2008).
Hammond, K. A. Adaptation of the maternal intestine during lactation. J. Mammary Gland Biol. Neoplasia 2, 243–252 (1997).
Amos-Landgraf, J. M. et al. Sex disparity in colonic adenomagenesis involves promotion by male hormones, not protection by female hormones. Proc. Natl Acad. Sci. USA 111, 16514–16519 (2014).
Manson, J. E. et al. Menopausal hormone therapy and long-term all-cause and cause-specific mortality: the women’s health initiative randomized trials. J. Am. Med. Assoc. 318, 927–938 (2017).
Gunter, M. J. et al. Insulin, insulin-like growth factor-I, endogenous estradiol, and risk of colorectal cancer in postmenopausal women. Cancer Res. 68, 329–337 (2008).
Jiang, H. & Edgar, B. A. EGFR signaling regulates the proliferation of Drosophila adult midgut progenitors. Development 136, 483–493 (2009).
Lee, T. & Luo, L. Mosaic analysis with a repressible cell marker for studies of gene function in neuronal morphogenesis. Neuron 22, 451–461 (1999).
Palanker, L. et al. Dynamic regulation of Drosophila nuclear receptor activity in vivo. Development 133, 3549–3562 (2006).
Kozlova, T. & Thummel, C. S. Spatial patterns of ecdysteroid receptor activation during the onset of Drosophila metamorphosis. Development 129, 1739–1750 (2002).
Han, S. K. et al. OASIS 2: online application for survival analysis 2 with features for the analysis of maximal lifespan and healthspan in aging research. Oncotarget 7, 56147–56152 (2016).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
Liang, J., Balachandra, S., Ngo, S. & O’Brien, L. E. Feedback regulation of steady-state epithelial turnover and organ size. Nature 548, 588–591 (2017).
Zheng, W. et al. Dehydration triggers ecdysone-mediated recognition-protein priming and elevated anti-bacterial immune responses in Drosophila Malpighian tubule renal cells. BMC Biol. 16, 60 (2018).
Su, Y.-H. et al. Diet regulates membrane extension and survival of niche escort cells for germline homeostasis via insulin signaling. Development 145, dev159186 (2018).
Cáceres, L. et al. Nitric oxide coordinates metabolism, growth, and development via the nuclear receptor E75. Genes Dev. 25, 1476–1485 (2011).
Acknowledgements
We thank J. Zhou, I. Miguel-Aliaga, C. Thummel, P. Patel and L. O’Brien for stocks and discussions. We thank O. Salem of McMaster Immunology Research Centre for Fig. 3i and M. AbdelMoety of American University of the Middle East for the chemical structure in Fig. 3i. This work was supported by ERC AdG 268515, DFG SFB873 and NIH GM124434 to B.A.E., and by the Helmholtz Zukunftsthema ‘Aging and Metabolic Programming’ (AMPro) to A.A.T.
Author information
Authors and Affiliations
Contributions
S.M.H.A. performed and analysed all experiments except Extended Data Figs. 8m, 9a–d (G.O.P.-S.) and Extended Data Fig. 6a, e–q (C.P.). J.A.M. contributed to Extended Data Fig. 8a, n and Fig 2o. The conception and design of experiments was by S.M.H.A., B.A.E., J.A.M. and A.A.T. Image-processing methods were designed by D.K. S.M.H.A., A.A.T. and B.A.E. wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature thanks Henri Jasper, Marc Tatar and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 20HE feeding promotes sexually dimorphic ISC mitotic activity.
a, Male ISCs do not divide strongly in response to infection elicited by pathogenic bacteria, but divide to a similar extent as mated female ISCs in response to 20HE feeding, quantified by counting the number of dividing ISCs per midgut using pH3 staining (also termed the mitotic index) in males and mated females after 16–18 h treatment with 5 mM 20HE or pathogenic P.e. infection. Males are fully and equally competent to respond to 20HE treatment as mated females. b, Mating boosts the mitotic divisions of ISCs. Feeding 0.1% SDS for 16 h to virgin females induces ISCs mitoses and this is inhibited by masculinizing ISC clones using sxl or tra RNAi. Mating increases the ISC mitotic responses to SDS feeding and restores the ability to masculinized ISCs to divide to stress. c, Mating induces basal ISC mitoses in both female (control) ISCs and in masculinized ISC clones with tra or sxl depletion. d, 20HE feeding leads to the proliferation and expansion of both control ISCs and ISCs of traRNAi masculinized progenitors. Representative images are shown 16 h after 5 mM 20HE feeding. This experiment was repeated three times with similar results. Quantification is shown in Fig. 1a. e, Quantification of ISC division at different time points (6, 9 and 12 h) after feeding 0.1% SDS to mated females. f–j, Males or mated females of the genotypes Gal4.DBD-Usp.LBD>GFP (Gal4-Usp>GFP) (f) or Gal4.DBD-EcR.LBD>GFP (Gal4-EcR>GFP) (g–j) were heat-shocked for 30 min to induce expression of the ligand sensor system, and then either infected with P.e. or fed with 5 mM 20HE or vehicle and dissected 18–20 h later. These GFP ligand traps express GFP under the control of heat-inducible promoter and mark cells with active 20HE signalling. When fed with vehicle, both Gal4-EcR>GFP and Gal4-Usp>GFP flies were expressed in a few cells in the R4 region posterior midgut (image shown) and in many more in the anterior midgut (image not shown). White arrows indicate cells that are doubly positive for delta or Su(H) lacZ markers. Feeding of 5 mM 20HE caused a strong increase in GFP expression in the posterior midgut, indicating an upregulation in the activity of both reporters. GFP was expressed in many delta+ cells (g, h) and much fewer Su(H)+ cells (i, j) of both males and females after 5 mM 20HE feeding. Most of the remaining positive cells are enterocytes. After 20 h of P.e. infection, the GFP signal disappears from males and females guts, indicating that EcR is not involved in infection-induced stress response (g, h). However, the Usp reporter was still active in many gut cells as a consequence of P.e. infection (f). The Usp reporter was also positive in many cell doublets and bigger cells of the midgut. These reporter data suggest that EcR and Usp are both activated by exogenous 20HE feeding, but they act differently in response to infection. Representative images are shown. This experiment was repeated five times with similar results. For all panels, control flies express UAS-GFP instead of the transgene. The period of RNAi induction is indicated. Results in dot plots are from at least three independent biological replicates. Data are mean and s.d. n ≥ 10 are plotted for each genotype in each scatter plot. **P ≤ 0.01, ***P ≤ 0.001, ****P < 0.0001, Mann–Whitney test with two-tailed distribution. Exact n numbers and P values are in the Source Data. Scale bars, 50 μm (f) or 100 μm (d, g–j). The overnight standard period of feeding the flies was 16–20 h.
Extended Data Fig. 2 The second mitotic wave of 20HE requires EcR–Usp in progenitors whereas EcR is dispensable to ISCs in their response to P.e. infection.
a, Representative images of samples from Fig. 1c, h. Both EcR and Usp are required in progenitors for the mitoses induced 16 h after 20HE feeding, whereas only Usp is cell-autonomously required by the ISCs for P.e.-induced mitoses. Shown are images of progenitor accumulation in mated females after 20HE feeding or P.e. infection. b, ISCs depleted of EcR or its downstream target Eip75B are unable to form clones in response to 20HE feeding. Eip75B-null mutant clones also fail to regenerate the epithelium after P.e. infection. EcR-depleted or Eip75B-null mutant clones were generated by MARCM and analysed 12 days after clonal induction followed by 5 mM 20HE feeding or P.e. infection for 16–18 h. Vehicle-fed control clones were multicellular and spread throughout the epithelium, whereas EcR-depleted clones were considerably smaller, mostly between two and four cells, and rarely up to ten small cells per clone. Eip75B-null mutant clones remained mostly single ISC clones. After 16 h of 20HE feeding, the epithelium is populated with newly formed cells within the control clones; however, both EcR- and Eip75B-depleted clones remained unable to divide, indicating the ISC cell-autonomous requirement of EcR and Eip75B for ISC mitoses both basally and in response to exogenously fed 20HE. Similarly, after P.e. infection, GFP+ cells expanded in control clones, whereas Eip75B-null mutant clones were considerably smaller. c, Quantification of data in b by a macro designed to assess clonal sizes/maximum Z projection (Methods, Supplementary Data 2). d, Both EcR and Usp are required in gut progenitor cells for the 20HE-induced-mitotic response as shown by the reduced ISC mitotic activity 16 h after feeding 5 mM 20HE to flies with progenitor-specific depletion of EcR or Usp in males and mated females. Results shown are for a second RNAi line to complement the results in Fig. 1c. e, EcR or Usp depletion in ISCs abolishes ISC mitoses 16 h after feeding 5 mM 20HE to males and mated females. Results shown are for two different RNAi lines. f, EcR is required in EBs for the second wave of ISC mitoses induced 16 h after feeding 5 mM 20HE to males and mated females. Results shown are for two different RNAi lines. This experiment indicates that in contrast to the first wave (Fig. 1e), EcR is required non-cell autonomously in EBs for 20HE induced ISC divisions. g, EcR is non-autonomously required in ECs for maximal induction of ISC mitoses in response to 20HE. The Myo1A-Gal4ts driver (Myo1A-Gal4 tub-Gal80ts) activates UAS target gene expression specifically in ECs. Results shown are for two different RNAi lines for both males and females, and for a dominant-negative isoform of EcR (EcR-ADN#2) in females. h, EcR in the nervous system is not required for intestinal 20HE-stem-cell induced mitoses. EcR depletion was induced using elav-Gal4 tub-Gal80ts, a pan-neuronal driver for the adult central nervous system. Sixteen hours after 5 mM 20HE feeding, ISCs mitoses were scored and midguts with EcR depletion in the CNS did not exhibit a change in their division rates in comparison to control females. i, EcR in enteroendocrine cells has a minimal role in 20HE-induced ISC mitoses of the midgut. Slightly compromised mitotic indexes in 20HE-fed mated females after enteroendocrine cell-specific depletion of EcR in EEs using the enteroendocrine cell-specific prosV1-Gal4 tub-Gal80ts driver indicate that EcR in enteroendocrine cells is dispensable to the 20HE induced ISC mitoses. Results shown are for two different RNAi lines. j, 20HE only transiently induces ISC mitoses, quantified by mitotic indices of male and female wild-type flies subjected to two-day of the indicated treatment regimes. ISC proliferation is restored to basal levels after 5 mM 20HE was withdrawn, which suggests that the actions of 20HE are not detrimental. Male and female flies were fed vehicle or 20HE in different successions such that flies were exposed for 20 h to the first treatment, then for another 24 h to the second treatment. ISC mitoses returned to basal levels after 16–20 h treatment with 20HE then vehicle. k, Expression of an EcR-A dominant-negative isoform inhibits the ISC proliferative response to 5 mM 20HE but not to enteric infection. Left, images of progenitors marked with esg-Gal4 after P.e. infection or 5 mM 20HE feeding, indicative of ISC proliferation in control mated females. Right, mitotic counts. l, 20HE signals mostly through isoform EcR-A to mediate ISC proliferation. Progenitor-specific expression of EcR-ARNAi and EcR-BRNAi shows that EcR-A, more than EcR-B, is required in ISCs for their mitotic response 16–20 h after feeding of 20HE. Knockdown of neither EcR-A nor EcR-B had an effect on the P.e.-induced ISC mitoses. m, EcR isoform A is much more important than isoform B for driving the intestinal hyperplasia, as shown in images of posterior midguts of mated females expressing different EcR dominant-negative isoforms. Left, images of clonal expansion under basal conditions at 5 days after induction of expression of different EcR dominant-negative isoforms in mated female midguts. Right, ISC mitotic counts. n–q, EcR in ISCs or other differentiated cells is not required for the P.e.-induced mitotic response of ISCs, whereas Usp is cell-autonomously required by ISCs to proliferate in response to P.e. infection. Quantification of the mitotic indexes of ISCs after P.e. infection in mated females in which EcR or Usp was depleted: constitutively in all cells using the tub-gal4ts driver (n), in EBs (o), in ISCs (p) or in ECs (q). Collectively, these experiments indicate a functional bifurcation of EcR and Usp, in which Usp is essential in ISCs for the P.e.-induced ISC response. RNAi was induced in progenitors of mated females for 8 days before 16–20 h of P.e. infection or 20HE feeding. For all panels, control flies express UAS-GFP instead of the transgene. The period of RNAi induction is indicated. Results in dot plots are from three independent biological replicates. Data are mean ± s.d. n ≥ 10 are plotted for each genotype in each scatter plot. *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P < 0.0001, Mann–Whitney test with two-tailed distribution. Exact n numbers and P values are in the Source Data. Representative images are shown from experiments that were repeated three times. Scale bars, 100 μm. The overnight standard period of feeding the flies was 16–20 h.
Extended Data Fig. 3 The second mitotic wave of 20HE regulates Jak–Stat signalling and requires Egfr signalling in the midgut progenitors.
a, Components of Egf signalling but not the Jak–Stat pathway are transcriptionally induced 6 h after 20HE feeding. mRNA levels of Egf ligands such as keren, spitz and their cleaving protease rho are transcriptionally induced whereas unpaired cytokines upd2, upd3, and Jak-Stat target Socs36E are not induced 6 h after 20HE (light green bars) relative to vehicle-fed control females (dark pink bars). By contrast, P.e. infection causes a strong induction of Jak-Stat signalling components upd2, upd3, Socs36E as well as a milder upregulation of Egf signalling components keren, vein and rho (light pink bars). Mated female midguts of wildtype flies were fed with vehicle, P.e. or 5 mM 20HE for 6 h then expression levels in guts were determined by RT–qPCR. Expression is indicated as mean fold change relative to vehicle-treated midguts ± s.d. (n = 3). b, Left, representative images of three categories of activity for the phenotypes of STAT92E-GFP reporters on chromosome II or III. The frequency of phenotype was quantified (right; and in g) in reference to phenotypes observed in the R4 region. Dark green text/bars denote no activation of the reporter. Bright green text/bars denote a mild activation pattern. Purple text/bars denote the strongest activation pattern. 5–7-day-old mated females were used for the experiment. Right, under homeostatic conditions, the reporter expresses GFP only in ISCs (dark green bar). At 6 h after 20HE feeding, GFP is localized in midgut progenitors all over the gut (bright green bar). 18% of the guts that express the reporter on chromosome II show a slight accumulation of GFP in other cells after 20HE feeding, but the GFP signal was not as strong as in the category ‘GFP in many cells’. c–e, EcR is required in midgut progenitors (c) and EBs (d) but not ECs (e) for transcriptional induction of rho, upd2 and upd3 during the second mitotic wave in response to 20HE feeding. By contrast, induction of spitz and keren are unchanged relative to 20HE fed controls. qRT–PCR was performed on midguts from mated females 8 days after RNAi induction at 29 °C followed by feeding with vehicle or 5 mM 20HE for 16 h. Expression is indicated as mean fold change relative to vehicle-treated midguts ± s.d. (n ≥ 3). f, ISCs need to proliferate in order for rho, upd2 and upd3 to be induced during the second mitotic wave after 20HE feeding. Egf and Jak–Stat signalling are transcriptionally induced 16 h after 20HE feeding. Control midguts have a transcriptional induction of rho, upd2 and Socs36E and to a lesser extent upd3 mRNA levels (vehicle denoted as purple versus control 20HE-fed denoted as pink bars). Cell cycle arrest via string depletion or reduced Egfr signalling in midgut progenitors halts the upregulation of 20HE-induced rho, upd2, Socs36E and upd3. These data suggest that ISC division is cell autonomously controlled and this event is an initial requirement for the non-cell autonomous induction of promitotic factors to promote later ISC divisions. mRNA induction of spitz and keren is slightly decreased in string-depleted progenitors but are slightly higher in Egfr-depleted progenitors relative to 20HE fed controls. Mated female midguts of wild-type flies, string or Egfr-depleted progenitors for 8 days at 29 °C were fed with vehicle or 5 mM 20HE for 16 h then expression levels were determined by RT–qPCR. Expression is indicated as mean fold change relative to vehicle-treated midguts ± s.d. (n ≥ 3). g, 20HE feeding induces activity of a Jak–Stat reporter more mildly than P.e. infection. Frequency of phenotype occurrence is analysed based on the categories of activity in b. Under homeostatic conditions, the reporter expresses GFP only in ISCs (dark green bar). Sixteen hours after 20HE feeding, most midguts of the reporter on chromosome II have GFP localized in many midgut cells including polyploid ECs (purple bar). However, most midguts of the reporter on chromosome III have GFP localized in the midgut progenitors (bright green bar). By contrast, P.e.-infected midguts of the reporters on either chromosome showed a strong uniform activation pattern in all midgut cells of the R4 region. 5–7-day-old mated females were used for the experiment. h, The upd3-lacZ reporter is not activated by 20HE feeding. Images of the R4 region of the midgut showing basal expression of the upd3 reporter in vehicle-fed flies relative to strong activation of the reporter after P.e. infection. By contrast, 16 h of 20HE feeding did not appreciably activate the upd3 reporter. These data indicate that 20HE does not primarily activate upd3 to promote ISC mitoses in the midgut. 5–7-day-old mated females were used for the experiment. All images were acquired at the same settings and the intensities of activation are accurately represented. i, Left, representative images of Erk activity, assayed as dpErk showing the most prevalent phenotype for each condition. Right, quantifications of the prevalence of each phenotype are shown. Under non-stressed conditions, dpErk is present either in very few ECs per gut, or in progenitor cells and very few ECs. After enteric infection, there is a strong upregulation of dpErk mainly in ECs. Although 20HE feeding also induces dpErk in midguts, the pattern is distinct from the one caused by enteric infection. After 20HE feeding, dpErk in mainly visible in progenitors and young ECs, and the signal is often localized to small patches of cells. By contrast, P.e. infection induces strong dpErk broadly throughout the gut. dpErk is absent in non-stressed upd2 or upd2,3 mutants. Enteric infection induces dpErk also in upd2 or upd2,3 mutants, albeit to a lower level than wild-type flies. By contrast, upd2 or upd2,3 mutants show very little or no dpErk after 20HE feeding. 5–8-day-old mated females were used for the experiment. j, Upd2, Egfr and rho are required in gut progenitors for the second wave of mitoses induced by 20HE as shown by the diminished ISC mitoses 16 h after feeding 5 mM 20HE to mated females with progenitor-specific depletion of Upd2, Upd2+Upd3, Egfr or rho. k, Upd2 and rho are required in EBs for the second wave of mitoses induced by 20HE as shown by the diminished ISC mitoses 16 h after feeding 5 mM 20HE to mated females with EB-specific depletion of Upd2, Upd2 and Upd3 or rho. Results shown are for two different RNAi lines for Upd2. l, Upd2 but not Upd3 or rho is required in ECs for the second wave of mitoses induced by 20HE as shown by the diminished ISC mitoses 16 h after feeding 5 mM 20HE to mated females with enterocyte-specific depletion of Upd2, Upd2 and Upd3 or rho. m, Rho is partly required in EBs for the optimal ISC mitoses during the first mitotic wave in response to 6 h of 20HE feeding. ISCs were still able to divide at 6 h of 20HE feeding after rho depletion in EBs albeit at lower but non-significant levels relative to control flies. This result indicates that ISCs, with their intrinsic EGF signalling retain the ability to divide in response to 20HE in a cell-autonomous fashion. For all panels, control flies express UAS-GFP instead of the transgene. The period of RNAi induction is indicated. Results in dot plots are from three independent biological replicates except for the qPCRs in which the n numbers are indicated. N ≥ 10 are plotted for each genotype in the remaining scatter plots. Data are mean ± s.d. *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P < 0.0001, Mann–Whitney test with two-tailed distribution. Exact n numbers and P values are in the Source Data. Representative images are shown from experiments that were repeated at least three. Scale bars, 100 μm. The overnight standard period of feeding the flies was 16–20 h.
Extended Data Fig. 4 Long-term 20HE feeding promotes sexually dimorphic ISC division and gut growth.
a, 1 mM 20HE feeding does not obviously increase epithelial turnover in females. Representative images are shown and are relevant to Fig. 1i. b, 20HE feeding causes male-specific midgut growth also on a low-protein diet, quantified by counting mitotic indexes of males and females raised on 20HE-laced low-yeast sucrose solution or sucrose-yeast solution as vehicle. 20HE- or vehicle-fed female ISCs did not differ in their mitotic counts. However, 20HE-fed males had a strong increase in their mitotic index compared to vehicle-fed males. c, 20HE feeding enhances ISC mitotic activity in P.e.-infected males, altering their behaviour to resemble P.e.-induced ISC division in females, assayed by mitotic counts of males and females. Flies were raised on 20HE or vehicle-supplemented food for 12 days then the treatment was withdrawn overnight followed by P.e. infection for 20 h. Male ISCs that were 20HE-fed were able to respond to P.e. infection at similar rates to the age-controlled females fed on 20HE or vehicle. d, 20HE-fed virgins undergo epithelial turnover much faster than age-controlled virgins, which have infrequently dividing ISCs. Representative images (left) and quantification (right) of mitotic counts from control virgin flies 14 days after 20HE feeding. Both the frequency of dividing ISCs and progenitor cells of 20HE fed virgins resemble the behaviour of mated females. e, Eip75 and EcR are required in midgut progenitors to maintain proper midgut size, quantified as midgut areas in images of guts from mated females with progenitor-specific depletion of EcR or Eip75B aged for 42 days. f, Quantification of midgut lengths of control males, 20HE-fed males, control virgin females, or virgin females depleted of ecdysone via ovary-specific knockdown of dibRNAi, shows the plasticity of male and female midgut growth to 20HE levels. 20HE-fed males have increased midgut length in contrast to dibRNAi female virgins, with decreased 20HE levels and notably shorter guts. In both cases, there was a one-third gain or loss in midgut length in comparison to a control male or virgin female, respectively. g, Ecdysone signalling via EcR and Eip75B is required in ISC clones of mated females for maximal proliferation in response to SDS. ISC mitotic counts of virgin females are minimal under basal conditions. After SDS feeding, control ISC clones divide to regenerate the epithelium but EcR- or Eip75B-depleted ISC clones are significantly impaired in their ability to divide. RNAi was induced in ISC clones for 8 days before 16–18 h of 0.1% SDS feeding. For all panels, control flies express UAS-GFP instead of the transgene. The period of RNAi induction is indicated. Results in dot plots are from three independent biological replicates. N ≥ 10 are plotted for each genotype in the remaining scatter plots. Data are mean ± s.d. **P ≤ 0.01, ***P ≤ 0.001, ****P < 0.0001, Mann–Whitney test with two-tailed distribution (all panels except f) or ordinary ANOVA test followed by Bonferroni’s multiple comparisons test (f). Exact n numbers and P values are in the online Source Data. Long-term 20HE feeding indicates that 1 mM 20HE was fed to the flies for 12 (c) or 14 days (a, b, d). Representative images are shown from experiments that were repeated three independent times. Scale bars, 100 μm.
Extended Data Fig. 5 Mating requires ecdysteroidogenic enzymes from the early ovarian follicles and escort cells to induce ISC divisions in the gut through EcR–Usp, which causes increased stem-cell number and subsequent gut growth.
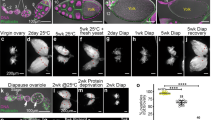
a, 20HE induces ISC mitoses in a dose-dependent manner in ISCs of virgin females. Virgin females were fed with different doses of 20HE and their mitotic indexes were assessed after 16–18 h of feeding. At 0.25–1.00 mM 20HE, ISCs divide similar to basal levels in mated females. At 2 mM 20HE, ISCs mildly divide (3–4 times higher than divisions induced by 1 mM 20HE). At 5 mM 20HE, ISCs divide at 10–11 times higher that divisions induced by 1 mM 20HE. b, The increase in width of the R4 region in response to mating in females requires EcR and Eip75B in progenitors. c, EcR is required in intestinal progenitors for their accumulation upon mating, shown by quantification of the GFP+ labelled areas of progenitors in the midgut after progenitor-specific depletion of EcR ± mating at early and later time points after mating. d, EcR-depleted ISC clones are unable to divide in response to mating, as quantified by the GFP+ clonal area in EcR-depleted ISC-derived clones and age-matched control clones. ISC-derived clones in control females have GFP+-labelled ISCs and all their subsequent progeny stably express GFP as well. e, Usp is required in progenitors for the mating-induced midgut growth as shown by quantification of midgut areas in females with Usp-depleted progenitors ± mating. f, EcR is cell-autonomously required in ISCs for mating-induced midgut growth, shown by quantification of midgut areas in females with EcR-depleted ISCs ± mating. After the first mating, control female midgut initially grows and midgut growth persists in flies that are raised repeatedly mated. This midgut growth requires EcR functions in ISCs. g, Ecdysone signalling via EcR, Usp and Eip75B are required in midgut progenitors for the mating-induced mitotic response, as shown by the reduced ISC mitoses upon 48 h mating in female midguts with progenitor-specific depletion of EcR, Usp or Eip75B. Virgins were left to mate for 48 h before dissection, then mitotic counts were assessed. Results shown are for a second RNAi line to complement the results in Fig. 2. h, EcR is cell-autonomously required in ISCs for mating-induced ISC mitoses shown by mitotic counts of midgut in females with EcR-depleted ISCs 72 h after mating. Results shown are for a second independent RNAi to complement Fig. 2f. i, Masculinized traRNAi progenitors undergo mating-induced expansion of GFP+ progenitors similar to controls, indicating that the mating effects on progenitors are independent of the sex determination pathway, quantified as GFP+ area of progenitors in the R4 region. Virgins typically have GFP-marked single cells (ISCs) or few pairs (ISC-EB). Shortly after mating, the ISC cells divide and the resulting progeny are transiently marked with GFP, but then turn off GFP expression as they differentiate. j, EcR is not required in ECs for mating-induced ISC mitoses. 48 h to 72 h after mating, ISCs of EcR depleted ECs midguts divide at similar rates to control midguts indicating that EcR in ECs is dispensable to mating-induced ISC mitoses. Results shown are for two different RNAi lines. k, Representative confocal image of GFP-expressing progenitors using esgts in females 5 days after mating. Flies were raised as virgins and were aged for 8 days (similar to conditions in Fig. 2b), and then mated for 5 days. Females were always mated to males with no genetic manipulations. Equal number of males and females were allowed to mate (a ratio of 1:1). Image is acquired in the R4 region. This suggests that the strong mitotic effect of mating is transient. Scale bars, 100 μm. l, Rho and upd2 are transcriptionally upregulated in female midguts 24 h (green) or 72 h (orange) after mating relative to virgins (pink). 5–7-day-old control virgins were mated for 24 or 72 h, then mRNA expression levels were determined by RT–qPCR. Expression is indicated as mean fold change relative to vehicle-treated midguts ± s.d. (n = 4). m, Representative images of whole-body spo mutants that are either heterozygous and hence viable with no growth or egg-laying defects (top) or sterile, homozygous spo mutants rescued to adulthood with by a pulse of 20HE given to dechorionated embryos (bottom). Images are complementary to Fig. 2i. Scale bars, 1 mm. n, RNAi-mediated depletion of spo in ovaries blunts ISC mitoses in response to mating. The traffic jam (tj-Gal4) driver that is expressed in somatic gonadal cells was used for spo depletion. Flies were raised as virgins then mated for 72 h. o, spoRNAi depletes the spo gene efficiently. Constitutive driver tubts was used to deplete spo in mated females for 8 days, and then mRNA expression levels were determined by RT–qPCR. Expression is indicated as mean fold change relative to vehicle-treated midguts ± s.d. (n = 4). p, Ovary-derived ecdysone is required for the proper size of the midgut, shown by quantification of midgut areas in mated female midguts depleted of 20HE-synthesizing enzyme Dib in the ovary. The C587ts driver, which is expressed in escort cells and immature follicle cells of the ovary, is used to induce ecdysteroidogenic enzymes depletion. Decreased midgut area in mated females with reduced 20HE levels is completely rescued by raising females on exogenous 1 mM 20HE. DibRNAi was previously validated35. q, Depletion of EcR in midgut ECs does not significantly decrease their size 8 days after mating. Cells of the midgut were stained with CellMask, a plasma membrane stain, and a custom macro (Supplementary Data 1) was used to segment the cells according to size. Shown is a frequency distribution of the different cell sizes. EcR-depleted ECs have a bigger proportion of cells sized 75–175 μm2 than control midguts. However, the differences in distribution of the cell sizes are statistically non-significant. Data are from n ≤ 5 stacks of midguts taken at the R4 region. r, Basal levels of EcR signalling are required to maintaining the optimal number of progenitors in the midgut as shown by quantification of GFP+ progenitors in mated females expressing dominant-negative EcR-A in comparison to the control. s, Basal levels of Eip75B are required for maintenance of ISCs in non-stressed flies, quantified by the number of GFP+ progenitors in mated females after progenitors-specific depletion of Eip75B. A small reduction of progenitor numbers (~25%) suggests that Eip75B is not critical for ISC survival. Note that y axis does not go to zero. t, Control midguts display an increase of delta+ cells at several time points following mating shown by quantification of delta+ (red) and Su(H)+ (green) cells. At 24 h after mating, most delta+ cells remain singlets, similar to virgins. At 40 h after mating, most delta+ cells expand to become doublets to triplets (Fig. 2k). At 7 days after the first mating most delta+ cells are again singlets; however, their numbers are irreversibly increased relative to virgins. Females were mated to males with no genetic manipulations. Equal number of males and females were allowed to mate (a ratio of 1:1) and females were allowed to mate for 18–20 h after which males were removed, except for the condition ‘raised mated for 7 days’, in which males were always in the vial with the females. Images are acquired in the R4 region. This suggests that mating induces an initial symmetric increase in the number of ISCs that is irreversible. Representative images for other conditions and quantifications are shown in Fig. 2k. Each dot represents a gut, and the percentage of delta+ or Su(H)+ cells is calculated from absolute number of positive cells relative to total DAPI+ cells. Scale bars, 100 μm. For all panels, control flies express UAS-GFP instead of the transgene. The period of RNAi induction is indicated. Results in dot plots are from three independent biological replicates. n ≥ 10 are plotted for each genotype in the remaining scatter plots. Data are mean ± s.d. *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P < 0.0001, ordinary ANOVA test followed by Bonferroni’s multiple comparisons test (gut measurements in b, e, f, p) or Mann–Whitney test with two-tailed distribution (all other panels). Exact n numbers and P values are in the online Source Data. Representative images are shown from experiments that were repeated three times.
Extended Data Fig. 6 Ovaries of the esg-Gal4ts, esgts Su(H)-Gal80 midgut drivers have GFP expression in their germaria in a subset of escort cells.
a, Top, most midgut drivers express GFP in ovary germaria. The frequency of germaria expressing GFP is displayed in the bar graph. Some ovaries with the esgts driver have no GFP in their germarium while almost all ovaries of the esgts Su(H)gal80 driver express GFP. Bottom, the number of GFP+ cells per germarium for both midgut drivers esgts or esgts Su(H)-Gal80, which are expressed in midgut progenitors and ISCs respectively. Further examination of esgts driver shows that it is expressed in approximately 4 escort cells, whereas the esgts Su(H)-Gal80 driver shows expression in around 14 escort cells. The number of germaria analysed is indicated. Control germaria typically have 45–70 escort cells36. b, Mated females with EcR- or Eip75B-depleted midguts have reduced reproductive output. This graph is related to Fig. 2p. Average eggs per fly per 3 days are plotted instead of the cumulative sums. Flies that died during the experiment were excluded in the analysis. c, Mated females with EcR- or Eip75B-depleted midguts have reduced reproductive output. Flies with control, EcR- or Eip75B-depleted midgut progenitors were raised as virgins for 8 days and then allowed to mate to males with no genetic manipulations at a ratio of 1:1 in populations of 5 females and 5 males. Eggs were collected from the fly vials every day for up to 11 days and the average total eggs per fly every 3 days is plotted. An independent alternative second RNAi is shown to complement data in Fig. 2p. Data are mean and s.d. P values were determined by t-test with two-tailed distribution assuming unequal variance. d, Mated females with EcR- or Eip75B-depleted ISCs have reduced reproductive output. Flies with control, EcR- or Eip75B-depleted midgut ISCs were raised at 18 °C for 2 days maximum and were then shifted to 29 °C and allowed to mate to males with no genetic manipulations at a ratio of 1:1. Flies were pooled together the first night of mating to ensure mating then on the next day, single females were housed with a control male in single vials. Eggs were collected from the fly vials every 48 h for up to 14 days. Flies that died during the experiment were excluded in the analysis. Left, cumulative eggs laid across 14 days ± s.d. Right, the average total eggs per fly every 3 days plotted across 14 days ± confidence intervals. P values were determined by t-test with two-tailed distribution assuming unequal variance. Exact n numbers are in the online Source Data. e–h, esg-Gal4ts and esgts Su(H)-Gal80 drive expression in a small number of ovary escort cells. Drosophila ovaries are composed of 16 ovarioles. At the anterior tip of every ovariole, the germarium contains the germline stem cells and the somatic stem cells that constantly produce follicles or egg chambers. As the follicles progress to the posterior end of the ovariole, they develop to lead to the formation of a mature egg. Follicle development is divided into 14 stages. In the most anterior part of the germarium (region I) the cap cells and the escort cells constitute the niche required for the maintenance of the GSCs and the proper differentiation of the early germline cyst. We detected expression of the esg-Gal4ts and the esgts Su(H)-Gal80 drivers within the germarium in a subset of escort cells (a). Confocal sections of follicles from stage 2–7 (e), stage 9 (h) and germaria (f, g) isolated from esg-Gal4ts flies and stained for GFP (green), coracle (red) and DNA (DAPI, grey). No GFP signal was detected in follicles from stage 2 to 9 (e, h) or in later stages (not shown). However, 96% of germaria showed GFP in a subset of cells in the anterior region I (f, g). The GFP-expressing cells were located in between the germline cysts and exhibited a triangular shape indicating that they were the escort cells. i–l, All germaria from esgts Su(H)-Gal80, UAS-GFP flies express GFP in escort cells (a, j, k, l) and no GFP expression was detected from stage 2 to 9 (i, j) or in later stages (not shown). m–q, We detected expression of the Switch GS5961-Gal4 driver within ovaries in the posterior follicular cells from stage 8 of oogenesis. Confocal section of follicles isolated from GS5961/UAS-GFP flies kept on yeast paste only (RU−) or yeast paste supplemented with RU486 (RU+) for 4 days and stained for GFP (green), actin (phalloidin, grey) or DNA (DAPI, grey). In the absence of RU486 induction, no GFP was detected in the ovary (m, n). After RU486 feeding, no expression was detected in germaria or follicles before stage 7 (p, q). At stage 7, a subset of the most posterior follicular cells started to express weakly the GFP, this expression was then stronger and spreading to more follicular cells in a posterior to anterior gradient during stage 8 of oogenesis (q, most posterior follicle) and maintained later on in most of the posterior follicular cells that cover the oocyte (o, stage 10). All pictures are presented with the anterior on the left and the posterior on the right.
Extended Data Fig. 7 JH receptors are required for ISC divisions, and exogenously fed JH inhibits ISC mitoses in response to other pro-mitotic stimuli.
a, JH receptors Met and Gce are required for exogenously fed 20HE to induce ISC mitosis. Virgin females were fed with 1.5 mM methoprene, 5 mM 20HE, or 20HE and methoprene in combination, and their mitotic indexes were assessed after 16–18 h of feeding. Knockdown of Met or Gce in progenitors blunted the proliferative response to all three fed stimuli. Virgins were aged for 8 days at permissive temperature then fed with the different hormone regimes for 16–18 h. b, c, Met and Gce receptors are required in midgut progenitors of mated females for P.e.-induced ISC mitoses. Mated females of indicated genotypes were aged for 8 days at permissive temperature then fed with P.e. for 18–20 h. b, ISC mitotic counts. c, Images of progenitor accumulation after P.e. feeding to mated females. d, Methoprene induces ISC mitoses in ISCs of virgin females. Virgin females were fed with active JH III ligand (JH), JH agonist methoprene (M), 2 mM or 5 mM 20HE, or the two compounds in combination, and their mitotic indexes were assessed after feeding for 16–18 h (left) or 72 h (right side). After 16–18 h of feeding, the average number of ISC mitoses per midgut was as follows. Vehicle fed: 3.8, 1 mM JH: 6.6, 1.5 mM methoprene: 8, 2 mM 20HE: 14, 5 mM 20HE: 41. A combination feeding of 1.5 mM methoprene with either 2 mM 20HE or 5 mM 20HE blunts mean ISC mitoses to 3.6 or 2.3, respectively. Combination feeding of 1 mM JH with 5 mM 20HE suppresses mean ISC mitoses to 11.5. After 72 h of feeding, the average number of ISC mitoses per midgut was as follows. Vehicle control: 5.5, 1.5 mM methoprene: 9.5, 5 mM 20HE: 13.5 mitoses, 5 mM 20HE + 1 mM JH 10.9, 5 mM 20HE + 1.5 mM methoprene: 10. These results indicate that 16 h of 2–5 mM 20HE act as a strong promitotic signal to ISCs of virgin females, but after 72 h the mean 20HE-induced mitoses drop towards basal levels. 1.5 mM methoprene causes a mild but persistent increase in ISC mitoses over 72 h. Overnight combination feeding of 20HE and 1.5 mM methoprene or 1 mM JH strongly suppressed 20HE-induced mitoses. e, Methoprene does not promote ISC mitoses in mated females. Mated females were fed with 1 mM or 5 mM active JH III ligand (“JH”), JH agonist methoprene (“M”), 1 mM or 5 mM 20HE, or 20HE and JH in combination and their mitotic indexes were assessed 16-18 h after feeding. Feeding of 1 mM or 5 mM JH, 1 mM 20HE, 1.5 mM or 5 mM methoprene do not induce mitoses in mated females. 5 mM 20HE feeding induces a boost of ISC mitoses that were suppressed by combination feeding with 1 mM JH. f, Exogenous JH feeding inhibits ISC mitoses when combined with other promitotic stimuli. Mated females were heat-shocked for 30 min, infected with P.e. for 18–20 h or fed with 20HE, either alone or in combination with 1 mM JH feeding for 16–18 h, and mitotic indexes were scored. In each case, feeding 1 mM JH suppresses the mitotic response of the stimulus. g, Ovarian ecdysteroidogenic enzymes are required for methoprene-induced mitoses of the midgut. 1.5 mM methoprene causes ISC mitoses in control midguts (mean of 6.5 mitoses relative to 2 mitoses in vehicle-fed flies). In animals in which the ecdysteroidogenic enzyme Dib is depleted in ovaries, methoprene failed to significantly induce ISC proliferation (mean of 3.3 mitoses relative to mean of 1.4 basal mitoses in dibRNAi vehicle-fed flies). Virgins were aged for 8 days at permissive temperature then fed with the different hormone regimes for 3 days. For all panels, control flies express UAS-GFP instead of the transgene. The period of RNAi induction is indicated. Results in dot plots are from three independent biological replicates. n ≥ 10 are plotted for each genotype in the remaining scatter plots. Data are mean ± s.d. *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P < 0.0001, Mann–Whitney test with two-tailed distribution. Exact n numbers and P values are in the online Source Data. Representative images are shown from experiments that were repeated three independent times.
Extended Data Fig. 8 Eip75B is a downstream ecdysone-inducible effector required to stimulate ISC proliferation, through Hr3 repression.
a, 20HE feeding or P.e. infection transcriptionally upregulate the ecdysone-inducible targets Eip75B and Broad. 5–7-day-old mated females were fed with 20HE or infected with P.e. for 6 h, then mRNA levels were determined by RT–qPCR on RNA from whole midguts. Expression is indicated as mean fold change relative to vehicle-treated midguts ± s.d. (n = 4). b, Broad and Eip75B are required by adult Drosophila midgut progenitors for P.e. or 20HE-induced ISC mitoses. Increased mitoses were observed after P.e infection or 20HE feeding in control mated flies, which were significantly blunted after Broad or Eip75B depletion in midgut progenitors. c, d, Eip75B is only cell-autonomously required in ISCs (c), but not EBs (d) for P.e.-or 20HE-induced ISC mitoses. Flies were fed with 20HE or P.e. for 16–20 h. Results are shown for two independent RNAi lines. e, Eip75B-null mutant clones are strongly impaired in their ability to divide and regenerate the epithelium. Eip75B-null mutant clones were generated by MARCM and analysed 6 days after P.e. infection. This experiment was done with a different recombinant mutant stock than that used in Extended Data Fig. 2. f, Eip75BRNAi blocks renewal of the midgut epithelium; brRNAi does not. Representative images from ISC clones of ageing epithelia with reduced levels of Eip75B or Broad. Broad depletion does not affect ISC clonal growth, whereas Eip75B depletion blocks any ISC growth and most cells remain singlets. g, Eip75B overexpression in ISC-derived esgFOts clones is pro-proliferative as shown by representative images of ISC clones in the epithelium of mated females. h, i, Eip75B is required by ISCs to divide in response to 20HE, haem, paraquat and enteric infection. h, Representative images of Eip75B-depleted ISC clones in response to the different stresses. Clonal growth to any stress stimulus is impaired. i, Quantification of mitotic counts. Results for P.e.-induced mitoses are shown for two independent Eip75BRNAi lines. j, Representative images of the heatshock-inducible Hr3 reporter (hs-Gal4.DBD-Hr3.LBD>GFP). Conditions of low Eip75B activity result in high Hr3 reporter expression and high Eip75B activity is reflected by low Hr3 reporter expression. Of note, owing to its transcriptional repressive activity, the Eip75B reporter cannot be used to monitor its activity30. Under basal conditions, midguts express high levels of Hr3 reporter. Hr3 activity is repressed by 20HE or haem feeding, P.e. infection (stimuli that require Eip75B) or co-expression of Eip75B. Nitric oxide (NO) inhibits Eip75B binding to Hr337. SNAP is a nitric oxide donor compound that modulates nitric oxide availability and is used to regulate Eip75B activity. However, increased nitric oxide levels through SNAP feeding relieved the repressive actions of P.e. and Eip75B on GFP expression. This indicates that in ISCs, Eip75B inhibits Hr3 and nitric oxide blocks this suppressive effect. Right, mitotic counts are shown for vehicle-fed, haem-fed, P.e., or P.e.+SNAP-fed mated females after 30 min heatshock (to induce the Hr3-GFP reporter) and 18–20 h of feeding. k, l, Hr3 overexpression strongly impairs epithelial renewal as the flies age, depicted by quantifications of mitotic indexes in k. l, Representative images of GFP-marked Hr3-overexpressing ISC clones showing impaired clonal growth in midguts of mated females. m, Hr3 depletion permits ISCs to divide in response to P.e. infection as shown by mitotic counts of Hr3-depleted ISC clones in mated females, which respond to P.e. infection at similar rates to control midguts. n, Repression of ISC mitoses in Eip75B-depleted esgFOts clones is rescued by Hr3RNAi as shown by mitotic counts of ageing or P.e.-infected guts with Eip75B, Hr3 depletion or both. This experiment shows that Hr3 is epistatic to Eip75B. For all panels, control flies express UAS-GFP instead of the transgene. The period of RNAi induction is indicated. The overnight standard period of feeding the flies was 18–20 h. Results in dot plots are from three independent biological replicates. n ≥ 10 are plotted for each genotype in the scatter plots. Data are mean ± s.d. ****P < 0.0001, Mann–Whitney test with two-tailed distribution. Exact n numbers and P values are in the online Source Data. Representative images are shown from experiments that were repeated three times. Scale bars, 100 μm.
Extended Data Fig. 9 Nitric oxide modulates the interaction of Eip75B and Hr3 to regulate ISC division.
a–d, Eip75B is not required in other midgut cell types besides progenitors for P.e. infection to induce ISC proliferation. Eip75B was depleted in progenitors using esg-gal4ts (two independent RNAi lines are shown to complement results in Fig. 2) (a), visceral muscle using how-Gal4ts (b), ECs using Myo1A-gal4ts (c), or enteroendocrine cells using prosV1-gal4ts (d). e, Overexpression of Hr3 in ISC-derived clones impedes the mitotic ability of ISCs to divide in response to P.e. infection. f, g, Inhibition of nitric oxide (NO) rescues the ISC mitotic activity of Hr3-overexpressing progenitors. f, ISC mitotic counts. g, Representative images of progenitor-specific overexpression of GFP with or without Hr3 followed by P.e. infection alone or in combination with the NO inhibitor l-NAME. NO represses the ability of Eip75B to interact with Hr3 hence, allowing transcriptional regulation of Hr3 targets. Treatment with l -NAME rescued the ISC ability to divide and progenitors expanded to fill the epithelium similar to the control mated females after infection (compare to results in Extended Data Fig. 8j). h, Model summarizing the regulation of Eip75B, Hr3 and Broad. i, Model summarizing the crosstalk between the gut and the ovary. For all panels, control flies express UAS-GFP instead of the transgene. The period of RNAi induction is indicated. The overnight standard period of feeding the flies was 18–20 h. Results in dot plots are from three independent biological replicates. n ≥ 10 are plotted for each genotype in the scatter plots. Data are mean ± s.d. ****P < 0.0001, Mann–Whitney test with two-tailed distribution. Exact n numbers and P values are in the online Source Data. Representative images are shown from experiments that were repeated three times. Scale bars, 100 μm.
Extended Data Fig. 10 Ovary-derived 20HE promotes intestinal dysplasia through EcR, Usp and Eip75B, which may affect Drosophila lifespan.
a, The number of mitotic cells in midguts increases with age, and this is inhibited by RNAi-mediated knockdown of EcR or Usp in ISC clones (esgFOts). Mitotic counts are shown at 19, 23 and 27 days after eclosion in non-stressed female guts. b, Basal 20HE levels promote age-dependent intestinal dysplasia. Mitotic indexes are shown in aged mated female midguts from flies ubiquitously expressing dibRNAi at two different ages after RNAi induction. c, Ovary ecdysone is required for ISC mitoses in non-stressed animals. Young and old mated females with spo knockdown in their ovaries have reduced ISC mitoses compared to controls. This was rescued by feeding the flies 1 mM 20HE. A second independent RNAi for spo is shown to complement data in Fig. 2. d, Representative images for the three classes of tumour phenotypes used to score mated female tumours in Fig. 3. e, 20HE feeding potentiates the tumour growth in NRNAi males. Left, representative images with which males have been scored in Fig. 3. Males exhibiting big tumour clusters of at least 30 neighbouring cells along the gut were classified strong. By contrast, guts with one or two tumour clusters with less than ten neighbouring cells were classified mild. Right, quantifications are derived by calculating the ratio between the GFP+ area and DAPI+ area. Tumour induction was commenced a few days before 20HE feeding. f, 20HE feeding potentiates the tumour initiation in virgin females with NRNAi. Representative images are shown for the quantifications presented in Fig. 3. Guts with no tumour clusters and just doublets of progenitor cells were classified as mild. Guts with tumour clusters of fewer than 10 neighbouring cells were classified as moderate, and guts with tumour clusters of at least 30 neighbouring cells were classified as strong. g–i, Progeny of the GS5961-Gal4 UAS-EcRF645A genotype were mated for 48 h. The populations followed up were segregated based on their sex (males (g) and females (h)) and separated into groups of 25 flies per vial. Approximately half of the flies were fed 0.2 mg ml−1 RU486 to induce dominant-negative EcR expression in progenitors and the other half were fed with vehicle. RU486 or vehicle (ethanol) was deposited on the food vials 4–6 h before flipping the flies into the vials at 48-h intervals. Dead flies were visually identified and recorded. Lifespan assays were performed in two replicates and for each replicate the percentage survival was plotted as a function of days elapsed after the start of the assay. Statistical analysis was performed using log-rank test. χ2 represents chi-squared value and the P values were provided from pairwise comparison with Bonferroni correction. i, Experimental details and the percentage mortality of the male or female replicates. Exact n numbers and P values are in the online Source Data.
Supplementary information
Supplementary Information
This file includes a Supplementary Discussion relevant to the main text, and additional references.
Supplementary Table 1
The table lists full Drosophila genotypes for all animals used, for each figure panel.
Supplementary Table 2
This table lists all Drosophila stocks used in this study and, where applicable, their stock numbers from public stock centers.
Supplementary Table 3
This table lists the DNA sequences of forward and reverse primers used to quantify mRNAs by RT-qPCR in Figure 2l and ED Figures 3a, 3d, 3e, 3f, 5l, 5o, and 8a.
Supplementary Data 1
This file contains the Cell Size Macro.zip. For this macro, plasma membrane is stained and then cells are segmented with reference to DAPI nuclear stain.
Supplementary Data 2
This file contains the Clone Size Macro.zip. Using this macro, GFP-marked cell clones are selected then their areas are measured.
Supplementary Data 3
This file contains the GFP Area Macro.zip. This macro calculates the area of DAPI+ cells (if required), the area of GFP+ cells, or both then, and then normalizes GFP area to total gut area.
Supplementary Data 4
This file contains the Gut Analysis Macro 1 (Zeiss).zip. This macro derives length, width and areas of whole midguts prior to mounting for images acquired using a Zeiss Stereo Discovery.V8 stereo microscope.
Supplementary Data 5
This file contains the Gut Analysis Macro 2 (Leica).zip. This macro derives length, width and areas of whole midguts prior to mounting for images acquired using a Leica M205 FA stereo microscope.
Supplementary Data 6
This file contains the Cells Quantification Macro.zip. This macro defines nuclei based on DAPI stain, and delta+ cells, or su(H)+ cells based on channel type, and calculates absolute numbers of cells.
Source data
Rights and permissions
About this article

Cite this article
Ahmed, S.M.H., Maldera, J.A., Krunic, D. et al. Fitness trade-offs incurred by ovary-to-gut steroid signalling in Drosophila. Nature 584, 415–419 (2020). https://doi.org/10.1038/s41586-020-2462-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-020-2462-y
This article is cited by
-
The midgut epithelium of mosquitoes adjusts cell proliferation and endoreplication to respond to physiological challenges
BMC Biology (2024)
-
Drosophila activins adapt gut size to food intake and promote regenerative growth
Nature Communications (2024)
-
Microbes control Drosophila germline stem cell increase and egg maturation through hormonal pathways
Communications Biology (2023)
-
Functional transcriptome analyses of Drosophila suzukii midgut reveal mating-dependent reproductive plasticity in females
BMC Genomics (2022)
-
Sexual identity of enterocytes regulates autophagy to determine intestinal health, lifespan and responses to rapamycin
Nature Aging (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
