
Abstract
Women who have experienced adverse childhood events (ACEs) around puberty are at the greatest risk for neuropsychiatric disorders across the lifespan. This population is exceptionally vulnerable to neuropsychiatric disease presentation during the hormonally dynamic state of pregnancy. We previously established that chronic adversity around puberty in female mice significantly altered their HPA axis function specifically during pregnancy, modeling the effects of pubertal ACEs we also reported in women. We hypothesized that the pregnancy hormone, allopregnanolone, was involved in presentation of the blunted stress response phenotype by its interaction with the molecular programming that had occurred during pubertal adversity experience. Here, in adult mice previously stressed during puberty, allopregnanolone administration was sufficient to reproduce the decreased corticosterone response after acute stress. Examination of neuronal activation and the electrophysiological properties of CRF neurons in the paraventricular nucleus of the hypothalamus (PVN) found no significant changes in synaptic function that corresponded with the blunted HPA axis reactivity. However, at the chromatin level, utilization of ATAC-Seq profiling demonstrated a dramatic remodeling of DNA accessibility in the PVN following pubertal adversity. Altogether, these data establish a potential molecular mechanism whereby adversity during puberty can enact lasting transcriptional control that manifests only during a unique period of the lifespan where dynamic hormonal changes occur. These results highlight a biological process that may impart an increased risk for a highly vulnerable population, whereby pubertal programming of the PVN results in aberrant HPA axis responsiveness when exposed to the hormonal changes unique to pregnancy.
Similar content being viewed by others

Introduction
Exposure to adverse childhood experiences (ACEs) is a significant risk factor for adult neuropsychiatric disorders [1, 2]. Women who have experienced ACEs around the onset of puberty are at the greatest risk for neuropsychiatric disorders across the lifespan [3,4,5]. This population is particularly vulnerable to presentation of neuropsychiatric symptoms during the hormonally dynamic state of pregnancy and postpartum [6,7,8,9]. We previously established that chronic adversity during puberty in female mice mimicked pubertal ACEs in women, significantly reducing their stress reactivity, only during pregnancy and postpartum [5]. Recent studies from a large cohort of women have confirmed that ACE exposure during childhood predicts maternal psychosocial outcomes, including perception of current stress [10]. Risk of peripartum depression and anxiety are also elevated in women with high ACEs and are associated with long-term-negative outcomes for both mother and baby, including increased incidence of undesirable maternal behaviors and delayed achievement of developmental milestones in infants [11,12,13,14]. Yet, few studies have examined the biological programming that contributes to this risk for such a vulnerable population.
To determine the mechanism underlying this unique dysregulation, we examined the interaction of the pregnancy hormone allopregnanolone with the molecular changes that occurred during puberty. Allopregnanolone, a metabolite of progesterone, is found in high concentration in the brain during pregnancy and early postpartum, is implicated in normal pregnancy-related shifts in stress function, and has been recently approved to treat symptoms of postpartum depression [15,16,17,18]. Allopregnanolone potentiates the effect of GABA, and as our previous study implicated a role for increased sensitivity of the GABA system in the pubertal adversity phenotype, we hypothesized that pubertal adversity and pregnancy produced a synergistic effect to enhance inhibition of the stress response [5]. In examination of the lasting effects of pubertal adversity, we focused on the paraventricular nucleus of the hypothalamus (PVN), both a key brain region that regulates the hypothalamic-pituitary-adrenal (HPA) stress axis response and a region that we have previously identified to have disruption at the level of the transcriptome following pubertal adversity. We hypothesized that pubertal stress would lead to programming of the PVN at multiple molecular levels and thus probed neuronal activation, electrophysiological properties, and the chromatin landscape.
Methods
Full details of experimental procedures and analyses are provided in the Supplemental Materials and Methods.
Animals
All mice bred were virgin, in-house mixed C57BL/6:129 mice (N = 139) [5, 19,20,21], except those used for electrophysiological experiments, which were heterozygous Crh-IRES-Cre;Ai14 (N = 26), expressing tdTomato in CRF neurons [22, 23]. All procedures were approved by the University of Pennsylvania Institutional Animal Care and Use Committee and by the University of Maryland Baltimore Institutional Animal Care and Use Committee. All procedures were conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Pubertal stress
Administration of pubertal stress was performed as described previously [5, 19]. Mice underwent 14 days of chronic variable stress (CVS) starting on postnatal day (PN) 21, during which one stressor was administered per day (60 min fox odor exposure, 15 min restraint, 3x cage change in one day, 36 h of constant light, exposure to a novel object (marbles) overnight, novel 100 dB white noise overnight, and saturated bedding overnight). Animals in the CVS group were weaned into singly housed cages at the beginning of stress, and were pair-housed with a same sex, same-stress cage mate at the end of stress. Control mice remained with the dam until they were weaned at PN28 into pair-housed cages.
Breeding scheme
At 10–12-weeks of age, female mice were bred with naive male mice for 1–3 nights. Upon confirmation of a copulation plug, female mice were established as pregnant and were immediately removed to their own cages. Female mice were left undisturbed until testing.
Glucocorticoid response to acute stress
Nulliparous adult female mice, adult female mice in the late stage of pregnancy (17.5 days post conception [dpc]) and adult male mice were tested for HPA axis responsiveness to a 15 min restraint stress, as previously [5, 19]. Corticosterone levels were determined by 125I-corticosterone radioimmunoassay (MP Biomedicals). Each set of experiments represents a different set of subjects, such that mice were not tested more than once.
Pharmacological manipulation of allopregnanolone
To address the necessity of allopregnanolone in the blunted HPA axis response of pubertally stressed females, pregnant females at 17.5 dpc were given an intraperitoneal injection of indomethacin (5 mg/kg, Tocris 1708) or vehicle (25% w/v HPβCD) 2 h prior to a 15-min restraint stress. This dose and delivery decreases allopregnanolone levels in the brain [24]. To address the sufficiency of allopregnanolone to uncover a blunted HPA response in pubertally stressed males, allopregnanolone (3 mg/kg 20 h before, and 1 mg/kg 2 h before, Tocris 3653) or vehicle (25% w/v HPβCD) was administered twice via intraperitoneal injections prior to a 15-min restraint stress. This approach has been used previously to manipulate allopregnanolone effects in the brain [25, 26].
Immunohistochemistry for c-Fos
Pregnant females at 17.5 dpc and age-matched nonpregnant adults were exposed to a 15-min restraint stress. Ninety minutes following the start of restraint, females were anesthetized and subject to cardiac perfusion. For c-Fos immunohistochemistry, sections were stained for c-Fos (1:800, Synaptic Systems 226–004, guinea pig polyclonal) and Hoechst (Thermo Fisher Hoechst 33342). Hoechst-immunoreactive and c-Fos-immunoreactive cells were quantified from 4 to 6 sections per animal from each set of tissue stained. Cell density of Hoechst and c-Fos was quantified with the FIJI distribution of ImageJ [27], using the Otsu threshold method [28] and the Analyze Particles function. For analysis, density of c-Fos was normalized to density of Hoechst in each section.
PVN CRF electrophysiology
Pregnant females at 17.5 dpc and age-matched nonpregnant adults were deeply anesthetized with isoflurane prior to transcardial perfusion. After preparation, slices containing the PVN were identified for recording using anatomical landmarks, as well as expression of red fluorescent CRF-tdTomato cells. Spontaneous inhibitory postsynaptic currents (sIPSCs) were recorded using standard whole-cell voltage-clamp techniques.
Mouse tissue collection and analysis
Pregnant females at 17.5 dpc and age-matched nonpregnant females were anesthetized and decapitated. Brains were rapidly frozen on dry ice and were stored at −80 °C until analysis. Trunk blood was collected into EDTA-coated tubes and processed for plasma, which was stored at −80 °C until analysis. For analyses of PVN from frozen tissue, whole brains were cryosectioned at −20 °C. Using a hollow 1.0 mm needle, the PVN was removed according to the mouse brain atlas [29]. PVN micropunches were processed according to each specific analysis.
Allopregnanolone production
Allopregnanolone was quantified in blood plasma via enzyme-linked immunosorbent assay (ELISA) (Arbor Assays). Real-time quantitative PCR (qPCR) gene expression of enzymes in the allopregnanolone synthesis pathway was conducted on total RNA isolated from the PVN.
Western blot
For protein extraction, micropunches from PVNs of two littermates were pooled. Protein was separated and transferred to a nitrocellulose membrane. The membrane was probed with anti-GABAA receptor delta (GABAAR δ) antibody (1:1000; Novus Biologicals, NB300-200). As a loading control, the membrane was re-probed with anti-GAPDH antibody (1:1000; Abcam, ab128915). Quantification of sample lanes were performed in the AzureSpot program (Azure Biosystems) by an investigator blind to group. Within each sample, GABAAR δ values were normalized to GAPDH for quantitative analysis.
ATAC-sequencing
PVN micropunches were immediately dispensed into pre-chilled tubes and stored at −80 °C until processing. Each n represents the PVN from a single animal (n = 6/group). Nuclei were extracted, chromatin tagmented, and libraries prepared according to the OMNI-ATAC protocol, which was developed for frozen tissue [30]. Paired-end 75-bp sequencing of the libraries was performed in the Illumina Next 550 sequencer using the NextSeq High Output v2 kit.
RNA-sequencing
PVN micropunches were immediately dispensed into 500 µl of Trizol and stored at −80 °C until processing. Each n represents the PVN from a single animal (n = 6/group). Total RNA was extracted and prepared for RNA-Sequencing. Single-end 75-bp sequencing was performed on libraries on the Illumina NextSeq 550 sequencer using the NextSeq High Output v2.5 kit.
Statistical analysis
An investigator blind to group conducted all data collection and analysis. For hormonal, electrophysiological, and immunohistochemical findings, data were analyzed by two-way analysis of variance (ANOVA) with Fisher’s Least Significant Difference (LSD) post-hoc testing as appropriate. Western blot, gene expression, and allopregnanolone ELISA data were analyzed by t-test. Exact tests that were used for all data presented are in the Supplement (Table S1). The significance level was p < 0.05. All data for these measures are reported as mean ± SEM.
For electrophysiological data, Clampfit software (Molecular Devices) was used to identify individual sIPSCs using a variable-amplitude shape template set to detect positive-going currents with an amplitude above a threshold (typically 5 pA). Parameters of individual events collected included time of event, peak amplitude of event, and decay tau of event. Each 3-min recording was separated into 1-min bins, and the frequency, average amplitude, and average decay tau for each bin were calculated. Events with decay taus over 100 ms were rarely identified during analysis and were discarded from analysis. The average frequency, amplitude, and decay tau for ten bins for each cell were calculated, and the averages of each cell from a given animal were then averaged to produce a single n.
ATAC-Seq data were analyzed in the R environment for Mac and in the terminal using the following pipeline: RSubread [31] to align sequence reads to the genome, Rsamtools [32] to sort BAM files, MACS2 [33] to call nucleosome free regions, soGGi [34] to merge open regions within a group. To identify differentially expressed peaks, the DESeq2 package was used [35]. Database of Annotation, Visualization, and Integrated Discovery functional annotation clustering was used for determination of gene clusters that were significantly enriched within a gene set based on gene ontology terms, with an enrichment score more than 1.3 equivalent to a p < 0.05 [36, 37]. RRHO2 was used for identification of concordant and discordant peak expression in a threshold free manner [38]. HOMER was used to perform de novo and known transcription factor-binding motif analysis [39]. RNA-Seq data were analyzed in the R environment for Mac with the packages kallisto and DESeq2 [40]. To identify differentially expressed genes, the Benjamini–Hochbert false-discovery rate correction was applied and an adjusted p < 0.05 was used.
Results
Allopregnanolone is necessary and sufficient to unmask the blunted HPA axis programmed by pubertal stress
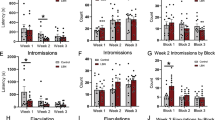
We have previously reported that pubertal stress (CVS) resulted in a blunted corticosterone response to acute restraint stress compared to mice not stressed during puberty (Control) later in adulthood when females are pregnant (Fig. 1a). We hypothesized that allopregnanolone would be necessary and sufficient to produce a blunted corticosterone response in adult, pubertally stressed mice. To determine if pregnancy-related levels of allopregnanolone were necessary to produce the blunted corticosterone response in pubertally stressed females, we blocked allopregnanolone synthesis with indomethacin that inhibits the enzyme 3α-hydroxysteroid oxidoreductase, which converts 5α-dihydroprogesterone to allopregnanolone, prior to a 15-min restraint stress (Fig. 1b). In Control females, there was no effect of indomethacin on the corticosterone response to stress (Fig. 1c, FTime(3,24) = 10.30, p = 0.0002; FDrug(1,8) = 0.5396, p = 0.4836; FTime*Drug(3,24) = 0.01778, p = 0.9967). However, in CVS females (Fig. 1d), indomethacin treatment resulted in a significantly higher rise in corticosterone in response to restraint stress (FDrug(1,7) = 0.6049, p = 0.4622; FTime(3,21) = 38.03, p < 0.0001; FTime*Drug(3,21) = 4.687, p = 0.0117). Indomethacin-treated females had more corticosterone than vehicle-treated females at 30 min following the start of restraint (p = 0.0417), but not at other time points. When compared by two-way ANOVA for treatment and stress, total corticosterone showed the same pattern as what we have previously reported, but failed to reach significance (FStress(1,15) = 1.994, p = 0.1784; FDrug(1,15) = 0.4096, p = 0.5318; FStress*Drug(1,15) = 2.033, p = 0.1744, with the Tukey post-hoc test between Control Vehicle and CVS Vehicle at p = 0.0836).

a We previously established that chronic variable stress during puberty leads to a blunted HPA axis only during pregnancy and postpartum in female mice. When HPA axis tissues were examined for transcriptomic changes, pubertal stress altered the transcriptome of the paraventricular nucleus of the hypothalamus (PVN), but not the pituitary gland, adrenal gland, or placenta. In the PVN, six immediate early genes (IEGs) were significantly increased in the PVN of CVS females during late pregnancy, suggesting that the chromatin is in a more open state, thus allowing the permissive transcription of these genes at baseline state [5]. The permissive transcription of these genes in CVS females during late pregnancy suggests that pubertal stress imparted a lasting effect on the chromatin in the PVN. b As allopregnanolone levels are high during pregnancy, we hypothesize that it is the exposure to high allopregnanolone levels in pregnancy that uncovers the blunted HPA axis phenotype in CVS females. To reduce allopregnanolone synthesis in control and pubertally stressed females, indomethacin was administered 2-h prior to a 15-min restraint stress. c In Control females, indomethacin had no effect on corticosterone production (n = 4–6/group). However, as predicted, indomethacin treatment elevated the HPA axis stress response in d CVS females compared to vehicle-treated CVS females (n = 4–5/group). In particular, Indomethacin-treated CVS females had significantly more corticosterone than vehicle-treated CVS females at 30 min following the start of restraint. e To demonstrate programming specificity of pubertal stress, adult male mice treated with allopregnanolone prior to a 15-min restraint stress showed no effect in f Control males (n = 6–7/group), but significantly reduced the corticosterone response in g pubertally stress males (n = 7/group). There was significantly lower corticosterone in CVS males treated with allopregnanolone than in vehicle-treated CVS males at both 15 and 30 min following the start of restraint. Gray bars (c, d, f, g) represent 15-min restraint time. Mice were freely moving in their home cage following restraint. *p < 0.05.
Pregnancy is a dynamic stage where changes occur at receptor, endocrine, and genetic levels. To determine whether allopregnanolone alone was sufficient to blunt the HPA axis, we utilized adult males that had been stressed during puberty. By testing males, we were able to examine whether the impact of pubertal stress on the response to allopregnanolone in adulthood resulted from sex-specific programming or if the presentation of the phenotype was sex-specific because males do not get pregnant. Adult control and pubertally stressed males were given allopregnanolone or vehicle prior to a 15-min restraint stress (Fig. 1e). Raising allopregnanolone in Control males had no effect on the corticosterone response (Fig. 1f, FTime(3,33) = 60.42, p < 0.0001; FDrug(1,11) = 0.3129, p = 0.5871; FTime*Drug(3,33) = 0.9845, p = 0.4120). In adult CVS males, allopregnanolone treatment significantly blunted the total corticosterone response to an acute restraint stress (Fig. 1g, FTime(3,36) = 59.42, p < 0.0001; FDrug(1,12) = 7.244, p = 0.0196; FTime*Drug(3,36) = 2.252, p = 0.0989). This effect was driven by significantly lower corticosterone in CVS males treated with allopregnanolone than in vehicle-treated CVS males at both 15 min (p = 0.0311) and 30 min (p = 0.0039) following the start of restraint. Altogether, these findings demonstrate that allopregnanolone is necessary and sufficient to produce a blunted HPA axis response only in pubertally stressed animals. Analysis of circulating levels of allopregnanolone at 17.5 dpc and relevant allopregnanolone synthesis pathway gene expression in the PVN suggests that pubertal stress did not change the total amount of allopregnanolone present during pregnancy (Supplementary Fig. S1).
Pubertal stress alters PVN responsiveness, but not by changing GABA synaptic function
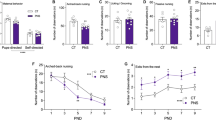
As the corticotropin-releasing factor (CRF) cells within the paraventricular nucleus of the hypothalamus (PVN) are responsible for initiation of an HPA response, we examined several cellular properties in the PVN. We first measured c-Fos immunoreactivity following a 15-min restraint stress in the PVN and other brain regions involved in stress responsiveness or known to be programmed by stressful life experiences (Fig. 2a). We hypothesized that the c-Fos response in the PVN would be blunted in pregnant females previously subjected to pubertal stress (CVS) compared with nonstressed controls (Control). In the PVN (Fig. 2b), there was a significant interaction between pubertal stress exposure and pregnancy on c-Fos immunoreactivity (FStress*Pregnancy(1,21) = 7.573, p = 0.0119; FPregnancy(1,21) = 8.627, p = 0.0079; FStress(1,21) = 12.01, p = 0.0023). Nonpregnant CVS females had significantly less c-Fos immunoreactivity than did Controls (p = 0.0009), while there was no difference between pregnant Control and CVS females (p = 0.5567). There was a main effect of pregnancy on c-Fos immunoreactivity (p < 0.05), which was to decrease c-Fos immunoreactivity relative to nonpregnant females. This decrease is consistent with several studies demonstrating a general suppression of c-Fos immunoreactivity in the PVN of rodents during late pregnancy [41,42,43]. This may represent a floor effect that could have obscured our ability to detect a further decrease by CVS. Of the other brain regions examined (Supplementary Fig. S2), only in the anteroventral bed nucleus of the stria terminalis did pubertal stress alter c-Fos immunoreactivity in response to an acute stressor.

Cellular responsiveness to an acute restraint stress was examined in adult females that had been exposed to pubertal stress or not, and that were virgins or pregnant 17.5 dpc. a c-Fos staining was performed in several brain regions known to regulate the stress axis. A representative photomicrograph is shown for the PVN. b In the PVN, there was an interaction between pubertal stress and pregnancy, such that virgin CVS females had a blunted c-Fos response (n = 4–8/group). CVS nonpregnant females had significantly less c-Fos immunoreactivity than did Controls, while there was no difference between pregnant Control and CVS females. c While the GABAAR δ subunit has been previously implicated in pregnancy-related changes to HPA axis responsiveness in the hippocampus, we find no difference in d. Total GABAAR δ in the PVN between Control and CVS females either in not pregnant (n = 4–5/group) or in 17.5 dpc pregnant females (n = 3–5/group). e The PVN of adult control and stressed females, either pregnant or not, was subject to slice electrophysiological recording. The inhibitory input to corticotropin-releasing factor (CRF) cells was recorded in the form of spontaneous inhibitory postsynaptic currents (sIPSCs). Representative traces are shown from pregnant Control (black line) and CVS (red line) females. f Pubertal stress had no effect on the frequency of sIPSCs in CRF neurons of adult females, whether pregnant or not (n = 5–8/group). g Similarly, the amplitude of the sIPSCs was not altered by pubertal stress exposure. *p < 0.05.
Altogether with our findings that the effect of CVS on the HPA was to blunt reactivity and that CVS animals were more sensitive to the GABA receptor allosteric modulator allopregnanolone, we hypothesized that CVS altered GABAergic inhibitory synaptic transmission in the PVN. Pregnancy-related changes to the GABAA receptor delta subunit (GABAAR δ) in the hippocampus have been implicated in affective dysfunction during and after pregnancy [44]. Further, allopregnanolone is known to strongly modulate αβδ-containing GABAARs [45]. Therefore, we examined the effect of pubertal stress and pregnancy on levels of GABAAR δ in the PVN using Western blotting (Fig. 2c). In nonpregnant females, there was no effect of pubertal stress on the level of GABAAR δ protein in the PVN (Fig. 2d, t(7) = 1.443, p = 0.1923). Similarly, there was no effect of pubertal stress on the expression of GABAAR δ in the PVN of pregnant females (Fig. 2d, t(6) = 0.8411, p = 0.4325).
An alternate hypothesis was that pubertal stress increased GABA function within the PVN, so that when allopregnanolone was present, a synergistic effect occurred, thus lowering the output of the PVN. As CRF cells in the PVN are a critical output cell in initiation of the HPA axis, we examined the inhibitory input to these cells and whether pubertal stress or pregnancy-altered baseline GABA function. We recorded spontaneous inhibitory postsynaptic currents (sIPSCs) from tdTomato-expressing neurons in the PVN of CVS and Control female CRF-reporter mice [22, 23] using whole-cell voltage-clamp (Fig. 2e). Neither the frequency (Fig. 2f, FStress*Pregnancy(1, 22) = 0.002969, p = 0.9570; FPregnancy(1, 22) = 0.1204, p = 0.7319; FStress(1, 22) = 1.048, p = 0.3171) nor amplitude (Fig. 2g, FStress*Pregnancy(1, 22) = 0.9619, p = 0.3374; FPregnancy(1, 22) = 0.5251, p = 0.4763; FStress(1, 22) = 1.488, p = 0.2354) of sIPSCs was altered by pubertal stress or pregnancy. Altogether, these findings suggest that pubertal stress did not significantly alter GABAergic inhibitory system within the PVN.
Allopregnanolone may interact with pubertal stress programming of the chromatin landscape
We have previously shown that pubertal stress resulted in increased expression of immediate early genes (IEGs) in the adult PVN in the absence of a stimulus, which is contrary to their canonical activity-dependent expression [5]. This implicated an openness of the chromatin landscape, which would allow IEGs to be transcribed permissively. Pubertal stress-induced programming of the chromatin state represents a lasting change that could impact multiple systems within the PVN.
Given our previous findings of increased IEGs in the PVN of pubertally stressed pregnant females, we hypothesized that the chromatin in the PVN would be more permissive, thus making these IEGs and other genes available for transcription [5]. To examine this hypothesis, we used ATAC-Seq to profile the open chromatin in the PVN of adult pubertally stressed (CVS) and nonstressed (Control) females (Fig. 3a and Supplementary Fig. S3A). For each replicate, we obtained an average of 184 million paired-end reads, which is sufficient for the detection of accessible regions and downstream transcription factor analysis [46]. Control regions Gapdh and Actb demonstrated consistent signal between replicates and groups (Fig. 3b). Open regions were defined as 100 bp or less in length, as previously described [47]. This selection criterion for open regions was confirmed by both the strength of the signal at transcription start sites (TSSs) that was consistent across groups (Fig. 3c) and by the stable annotation location of peaks to the genome (Supplementary Fig. S3B). To determine the number of open sites for each group, a cutoff was applied to all detected regions (present in at least two samples within each group). Adult, nonpregnant females had minimal differences in the number of open sites in PVN chromatin, regardless of pubertal stress experience. Control, nonpregnant females had 60,404 total open sites and nonpregnant CVS females had 59,175 open sites (Fig. 3d). Exposure to pregnancy had opposing effects on the number of open sites in the PVN, where pregnant Control females responded with decreased permissiveness of the chromatin (53,103 open sites) and pregnant CVS females responded with increased permissiveness of the chromatin (65,243 open sites) compared to their nonpregnant state. These findings suggested that Control females should respond to pregnancy with a downregulation of genes at the transcriptome level, while CVS females should respond with an upregulation. RNA-Seq analysis of the PVN confirmed this prediction (Supplementary Fig. S4). The effect of CVS on the accessible chromatin peaks was not consistent between the nonpregnant and pregnant state. RRHO analysis showed that the peaks that were the most/least accessible in CVS females were discordant when comparing this effect in nonpregnant and pregnant mice (Fig. 3e). This confirms the prediction from our previous transcriptomic findings that the chromatin in the PVN of pregnant CVS females was more open than chromatin of pregnant Control females, and that exposure to pregnancy exacerbated the differences between the treatment groups.

a The PVN of adult mice, either not pregnant or pregnant (17.5 dpc), that had been exposed to pubertal stress (CVS) or not was subject to ATAC-Seq (n = 6/group). Analysis of both the signal in b housekeeping genes and c the location of reads relative to transcription start sites (TSSs) shows that the ATAC-Seq results are of high quality and that group does not alter the basic representation of peak signal in control regions or location of peaks within the genome. d Examination of the number of open peaks within the PVN by group shows that Control and CVS females have a similar number of open peaks in the nonpregnant PVN that is dramatically altered in opposite directions by exposure to pregnancy. Control females respond to pregnancy with fewer open peaks in the PVN while CVS females respond to pregnancy with more open peaks in the PVN relative to the not pregnant state. e The effect of CVS on the accessible chromatin peaks is not consistent between the not pregnant and pregnant state. RRHO analysis shows increased signal in the upper left and lower right quadrant of the heatmap, which indicates a discordant effect of CVS on the chromatin landscape in not pregnant and pregnant females. f Heatmap displaying the differentially accessible peaks (162 peaks) when nonpregnant Control peaks were compared with nonpregnant CVS peaks. This confirms that there are latent differences in PVN chromatin in nonpregnant stressed females. Heatmap displaying the differentially accessible peaks (483 peaks) when pregnant Control peaks were compared to pregnant CVS peaks. This shows that there is an enhancement of differences in chromatin accessibility between Control and CVS females when they are exposed to pregnancy. Heatmaps are ranked from most open (red) to least open (blue) in Controls. g Transcription factor (TF)-binding motif analysis was performed on the differentially accessible peaks in Control females (not pregnant versus pregnant) or CVS females (not pregnant versus pregnant) for the effect of pregnancy. Pregnancy resulted in a dramatic remodeling of CVS chromatin that resulted in significantly more types of binding motifs becoming available throughout the chromatin. h These enriched motifs cluster into relevant TF families for controlling how the PVN responds to input. Example binding motifs are provided for several critical TF families.
We performed differential accessibility analysis to identify peaks with significantly different openness within the PVN based on pubertal stress and pregnancy experience (p < 0.05; log2 fold change >1 or <−1). There were 162 differentially accessible peaks when nonpregnant Control and nonpregnant CVS females were compared (Fig. 3f). Of these peaks, the majority (150 peaks) were more open in CVS females than in Controls. These findings confirm a latent programming of the PVN at the level of chromatin accessibility, where we detect subtle differences in outcomes when females are not pregnant. When pregnant Control and CVS chromatin was analyzed, there were 483 differentially accessible peaks in the PVN, recapitulating the enhancement of differences between the two groups during pregnancy, similar to the phenotype we have observed in the HPA axis response [5] (Fig. 3f). Functional annotation clustering of the differentially accessible peaks shows that these peaks converge on pathways that provide the potential to alter the way that the PVN responded to general remodeling associated with pregnancy, including categories: postsynaptic membrane, synapse, membrane integrity, cell differentiation, axon guidance, and calmodulin binding (Supplementary Fig. S5). This suggests that Control and CVS females may have a divergent response in how pregnancy remodels the structure or responsiveness of synapses and cells in the PVN.
To determine the transcription factors that would be associated with regions of chromatin accessibility during pregnancy in CVS females, we performed binding motif analysis using HOMER (Fig. 3g and Supplementary Table S2). The effect of pregnancy produced some overlap in significantly enriched binding motifs, including for that of the glucocorticoid receptor, progesterone receptor, and androgen receptor. Outside of this expected overlap in hormone-related-binding motifs, pregnancy induced a wider remodeling of transcription factor-binding sites in CVS females (83 enriched binding sites) than in the PVN of Control females (18 enriched binding sites, Fig. 3h). These enriched sites in CVS females clustered into several important families, including ETS-domain, forkhead, high-mobility group (HMG), and basic leucine zipper (bZIP). The most enriched family was the ETS-domain, which functions to recruit histone acetyl transferases or deacetylases to broadly regulate chromatin dynamics [48]. Altogether, these results suggest that there were lasting effects of pubertal stress on chromatin dynamics.
Discussion
Adverse childhood experiences (ACEs) that occurred during the plastic period of brain maturation, puberty, are one of the strongest predictors of adult neuropsychiatric disorders in women [49, 50]. For this population, ACEs may result in a more susceptible background upon which the dynamic hormonal changes that occur in pregnancy and postpartum represent a “second hit” that further increases risk for disease presentation [51]. We previously established that chronic adversity during puberty in female mice recapitulated pubertal ACEs in women, significantly altering their HPA stress axis function during pregnancy and postpartum [5]. To determine the mechanisms underlying this unique dysregulation, we examined the pubertal molecular programming and its interaction with the pregnancy hormone allopregnanolone.
We first utilized pharmacological manipulations in adult mice exposed to pubertal adversity to examine whether allopregnanolone, a progesterone metabolite implicated in pregnancy neuropsychiatric symptoms, was involved in the production of our maternal stress axis phenotype. Blocking allopregnanolone synthesis in pregnant females exposed to pubertal adversity disrupted phenotype presentation, while elevating circulating allopregnanolone levels in adult males exposed to pubertal adversity recapitulated it. These results suggest that allopregnanolone is both necessary and sufficient to cause the dysregulation of the HPA stress axis found during pregnancy in mice that had been exposed to adversity during puberty.
Next, we examined the molecular level at which pubertal adversity programmed the HPA axis. We previously reported that the transcriptome of the paraventricular nucleus of the hypothalamus (PVN) was disrupted by pubertal adversity, while the peripheral nodes of the HPA axis (pituitary and adrenal gland) remained unaffected [5]. Using c-Fos immunoreactivity as a readout of PVN activation, we found that fewer cells were activated in response to an acute stressor in nonpregnant females exposed to pubertal adversity, compared to mice not subjected to pubertal adversity. In pregnant mice, in contrast, pubertal adversity had no effect. These results are not consistent with our HPA axis function findings, where we instead observed a blunted HPA response only during pregnancy and not in the nonpregnant state. These results also do not follow the prediction from our previous transcriptome findings, where c-Fos gene expression was increased in pregnant females exposed to pubertal adversity. However, this discrepancy may be due to differences in measurement timing in pregnancy (18.5 dpc versus 17.5 dpc), c-Fos expression at baseline versus after acute stress, or a floor effect of pregnancy on c-Fos immunoreactivity. The PVN is populated by a number of different cell types, so it remains possible that cell-type-specific examination of c-Fos immunoreactivity would yield further insight that is masked by a limited approach. While there is clearly something different about how the PVN of adults exposed to pubertal adversity responds to acute stressors, it is not obvious how this manifests at the cellular level.
Allopregnanolone produces inhibitory effects in the brain via action at GABAA receptors containing delta (δ) subunits (GABAAR δ) [52]. We tested the hypothesis that our HPA stress axis phenotype was due to pubertal adversity programming of increased GABAAR δ expression, thus increasing brain sensitivity to allopregnanolone. However, in examination of protein levels in the PVN of adults we found no difference in GABAAR δ based on pubertal adversity in either pregnant or nonpregnant females, suggesting that changes in GABAAR δ are not involved in the detected phenotype. Next, we recorded spontaneous inhibitory postsynaptic potentials (sIPSCs) to determine if pubertal adversity affected responses to acute stressors by programming GABAergic synaptic function in stress-responsive corticotropin-releasing factor (CRF) neurons in the PVN. No significant differences in sIPSC amplitude or frequency were found between treatment groups. This suggests that pubertal adversity did not alter CRF-cell activation by changing spontaneous inhibitory input from GABAergic cells. The possibilities remain that differences in inhibitory input would be detected under different conditions, such as with the addition of local stimulation, or that pubertal adversity instead altered excitability of CRF neurons.
We next examined the molecular transcriptional machinery in adult females exposed to pubertal adversity. Our previous transcriptomic analysis of the PVN revealed widespread increase in the baseline expression of immediate early genes (IEGs) as a result of pubertal adversity [5]. Increased IEG expression suggested that the chromatin in the PVN of pregnant females exposed to pubertal adversity was in a more permissive state. We used ATAC-Sequencing to profile DNA accessibility in the PVN and found significantly more open peaks associated with pubertal adversity. The impact of pubertal adversity on chromatin accessibility was further enhanced in pregnancy, where there were ~3x more differentially accessible peaks. The perturbation of the normal chromatin landscape by pubertal adversity is likely due to epigenetic mechanisms, as epigenetic modifiers are responsible for chromatin conformation and can be altered by the environment [53,54,55,56]. Further, puberty represents a time when stress may alter the trajectory of the brain by enacting programming at the epigenetic level [57,58,59,60,61].
Using transcription factor-binding motif analysis, we identified distinct signatures of pregnancy-altered binding motifs that depended upon pubertal adversity [62]. Pregnancy induced a dramatic remodeling in the PVN specifically in females subject to pubertal adversity. The accessible transcription factor-binding sites in the PVN of pubertal adversity-exposed females clustered into several important families, with the most enriched families being those with an ETS-domain that contain transcription factors that interact with a wide variety of co-regulatory partners, can produce gene-specific responses, and recruit histone acetyl transferases or deacetylases [48]. Other transcription factor family motifs that were highly enriched in PVN of females exposed to pubertal adversity belong to families that can bind to condensed chromatin, are involved in DNA binding, and initiate alterations to post-translational modifications of histones [63,64,65]. Whether the chromatin was left in a generally unstable state by pubertal adversity or there was more specific programming of chromatin regulators is yet to be examined. However, open chromatin detected in the PVN of pregnant mice exposed to prior pubertal adversity suggests a mechanism whereby permissive transcription by these neurons allows them to be more dynamic in their response to neuroendocrine-negative feedback, thereby shutting down the stress response faster. While it is yet unknown what the mechanism of allopregnanolone action is in the remodeling of DNA availability in pubertally stressed adult females, our current data suggest an interaction at the chromatin level to uncover the pubertal stress programming.
In sum, we demonstrate that puberty is a sensitive window for adversity, potentially mediated through enduring changes to the chromatin that can permit lasting changes in transcriptional control, to increase risk of HPA axis dysfunction during later times of dynamic hormonal change, such as pregnancy and postpartum. Our findings are especially interesting in light of the recent approval of Brexanolone, a proprietary formulation of allopregnanolone, for the treatment of postpartum depression [18]. It is of critical importance that drug development focus on inclusion of women’s health as a priority, as prior research has shown that women experience greater prevalence of affective disorders and often respond differently than men to therapeutic treatments [66, 67]. While studies in mice with abnormal postpartum behaviors and a dysregulated HPA stress axis were responsive to Brexanolone [68,69,70], our data in mice and humans suggest that a subpopulation of women with high-pubertal ACEs who manifest with a hyporesponsive HPA axis in pregnancy and postpartum would respond to allopregnanolone treatment with a further exacerbation of stress axis dysfunction, the consequences for which are not currently known [5].
Funding and disclosure
The studies were supported by P50-MH099910, MH108286, MH104184, HD097093, ES028202 (TLB), and K99 HD091376 (KEM). The authors have no completing financial interests to disclose in relation to the work described.
References
Merrick MT, Ports KA, Ford DC, Afifi TO, Gershoff ET, Grogan-Kaylor A. Unpacking the impact of adverse childhood experiences on adult mental health. Child Abus Negl. 2017;69:10–9.
Rosenberg SD, Lu W, Mueser KT, Jankowski MK, Cournos F. Correlates of adverse childhood events among adults with schizophrenia spectrum disorders. Psychiatr Serv. 2007;58:245–53.
Weiss EL, Longhurst JG, Mazure CM. Childhood sexual abuse as a risk factor for depression in women: psychosocial and neurobiological correlates. Am J Psychiatry. 1999;156:816–28.
Dahl RE, Gunnar MR. Heightened stress responsiveness and emotional reactivity during pubertal maturation: implications for psychopathology. Dev Psychopathol. 2009;21:1–6.
Morrison KE, Epperson CN, Sammel MD, Ewing G, Podcasy JS, Hantsoo L, et al. Preadolescent adversity programs a disrupted maternal stress reactivity in humans and mice. Biol Psychiatry. 2017;81:693–701.
Dickens MJ, Pawluski JL. The HPA axis during the perinatal period: implications for perinatal depression. Endocrinology. 2018;159:3737–46.
Jones I, Chandra PS, Dazzan P, Howard LM. Bipolar disorder, affective psychosis, and schizophrenia in pregnancy and the post-partum period. Lancet. 2014;384:1789–99.
Borri C, Mauri M, Oppo A, Banti S, Rambelli C, Ramacciotti D, et al. Axis I psychopathology and functional impairment at the third month of pregnancy: results from the Perinatal Depression-Research and Screening Unit (PND-ReScU) study. J Clin Psychiatry. 2008;69:1617–24.
Vesga-Lopez O, Blanco C, Keyes K, Olfson M, Grant BF, Hasin DS. Psychiatric disorders in pregnant and postpartum women in the United States. Arch Gen Psychiatry. 2008;65:805–15.
Racine NM, Madigan SL, Plamondon AR, McDonald SW, Tough SC. Differential associations of adverse childhood experience on maternal health. Am J Prev Med. 2018;54:368–75.
Paulson JF, Dauber S, Leiferman JA. Individual and combined effects of postpartum depression in mothers and fathers on parenting behavior. Pediatrics. 2006;118:659–68.
Slomian J, Honvo G, Emonts P, Reginster J-Y, Bruyère O. Consequences of maternal postpartum depression: a systematic review of maternal and infant outcomes. Women’s Health Lond Engl. 2019;15:1745506519844044.
O’Leary N, Jairaj C, Molloy EJ, McAuliffe FM, Nixon E, O’Keane V. Antenatal depression and the impact on infant cognitive, language and motor development at six and twelve months postpartum. Early Hum Dev. 2019;134:41–6.
Duarte-Guterman P, Leuner B, Galea LAM. The long and short term effects of motherhood on the brain. Front Neuroendocrinol. 2019;53:100740.
Brunton PJ, Russell JA. Allopregnanolone and suppressed hypothalamo-pituitary-adrenal axis stress responses in late pregnancy in the rat. Stress Amst Neth. 2011;14:6–12.
Paul SM, Purdy RH. Neuroactive steroids. FASEB J Publ Fed Am Soc Exp Biol. 1992;6:2311–22.
Morrison KE, Cole AB, Thompson SM, Bale TL. Brexanolone for the treatment of patients with postpartum depression. Drugs Today. 2019;55:537.
Meltzer-Brody S, Colquhoun H, Riesenberg R, Epperson CN, Deligiannidis KM, Rubinow DR, et al. Brexanolone injection in post-partum depression: two multicentre, double-blind, randomised, placebo-controlled, phase 3 trials. Lancet Lond Engl. 2018;392:1058–70.
Morrison KE, Narasimhan S, Fein E, Bale TL. Peripubertal stress with social support promotes resilience in the face of aging. Endocrinology. 2016;157:2002–14.
Rodgers AB, Morgan CP, Leu NA, Bale TL. Transgenerational epigenetic programming via sperm microRNA recapitulates effects of paternal stress. Proc Natl Acad Sci USA. 2015;112:13699–704.
Jašarević E, Howard CD, Morrison K, Misic A, Weinkopff T, Scott P, et al. The maternal vaginal microbiome partially mediates the effects of prenatal stress on offspring gut and hypothalamus. Nat Neurosci. 2018;21:1061–71.
Taniguchi H, He M, Wu P, Kim S, Paik R, Sugino K, et al. A resource of Cre driver lines for genetic targeting of GABAergic neurons in cerebral cortex. Neuron. 2011;71:995–1013.
Madisen L, Zwingman TA, Sunkin SM, Oh SW, Zariwala HA, Gu H, et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat Neurosci. 2010;13:133–40.
Ahboucha S, Jiang W, Chatauret N, Mamer O, Baker GB, Butterworth RF. Indomethacin improves locomotor deficit and reduces brain concentrations of neuroinhibitory steroids in rats following portacaval anastomosis. Neurogastroenterol Motil. 2008;20:949–57.
Brunton PJ, Donadio MV, Yao ST, Greenwood M, Seckl JR, Murphy D, et al. 5α-reduced neurosteroids sex-dependently reverse central prenatal programming of neuroendocrine stress responses in rats. J Neurosci. 2015;35:666–77.
Brunton PJ, McKay AJ, Ochędalski T, Piastowska A, Rębas E, Lachowicz A, et al. Central opioid inhibition of neuroendocrine stress responses in pregnancy in the rat is induced by the neurosteroid allopregnanolone. J Neurosci. 2009;29:6449–60.
Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, et al. Fiji: an open-source platform for biological-image analysis. Nat Methods. 2012;9:676–82.
Otsu N. A threshold selection method from gray-level histograms. IEEE Trans Syst Man Cyber. 1979;9:62–6.
Paxinos G, Franklin K. Paxinos and Franklin’s the mouse brain in stereotaxic coordinates. 4th ed. Academic Press; 2012.
Corces MR, Trevino AE, Hamilton EG, Greenside PG, Sinnott-Armstrong NA, Vesuna S, et al. An improved ATAC-seq protocol reduces background and enables interrogation of frozen tissues. Nat Methods. 2017;14:959–62.
Liao Y, Smyth GK, Shi W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 2019;47:e47–7.
Morgan M, Pages H, Obenchain V, Hayden N. Rsamtools: binary alignment (BAM), FASTA, variant call (BCF), and tabix file import. 2019. https://doi.org/10.18129/B9.bioc.Rsamtools.
Zhang Y, Liu T, Meyer CA, Eeckhoute J, Johnson DS, Bernstein BE, et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 2008;9:R137.
Dharmalingam G, Carroll T. soGGi: visualise ChIP-seq, MNase-seq and motif occurrence as aggregate plots Summarised Over Grouped Genomic Intervals. 2019. https://doi.org/10.18129/B9.bioc.soGGi.
Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15:550.
Huang DW, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009;4:44–57.
Huang DW, Sherman BT, Lempicki RA. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009;37:1–13.
Cahill KM, Huo Z, Tseng GC, Logan RW, Seney ML. Improved identification of concordant and discordant gene expression signatures using an updated rank-rank hypergeometric overlap approach. Sci Rep. 2018;8:9588.
Heinz S, Benner C, Spann N, Bertolino E, Lin YC, Laslo P, et al. Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell identities. Mol Cell. 2010;38:576–89.
Bray NL, Pimentel H, Melsted P, Pachter L. Near-optimal probabilistic RNA-seq quantification. Nat Biotechnol. 2016;34:525–7.
Windle RJ, Wood SA, Kershaw YM, Lightman SL, Ingram CD. Reduced stress responsiveness in pregnancy: relationship with pattern of forebrain c-fos mRNA expression. Brain Res. 2010;1358:102–9.
da Costa AP, Wood S, Ingram CD, Lightman SL. Region-specific reduction in stress-induced c-fos mRNA expression during pregnancy and lactation. Brain Res. 1996;742:177–84.
Russell JA, Douglas AJ, Brunton PJ. Reduced hypothalamo-pituitary-adrenal axis stress responses in late pregnancy: central opioid inhibition and noradrenergic mechanisms. Ann N. Y Acad Sci. 2008;1148:428–38.
Maguire J, Ferando I, Simonsen C, Mody I. Excitability changes related to GABAA receptor plasticity during pregnancy. J Neurosci. 2009;29:9592–601.
Zheleznova NN, Sedelnikova A, Weiss DS. Function and modulation of δ-containing GABAA receptors. Psychoneuroendocrinology. 2009;34S1:S67–73.
Buenrostro J, Wu B, Chang H, Greenleaf W. ATAC-seq: a method for assaying chromatin accessibility genome-wide. Curr Protoc Mol Biol Ed Frederick M Ausubel Al. 2015;109:21.29.1–9.
Buenrostro JD, Giresi PG, Zaba LC, Chang HY, Greenleaf WJ. Transposition of native chromatin for multimodal regulatory analysis and personal epigenomics. Nat Methods. 2013;10:1213–8.
Sharrocks AD. The ETS-domain transcription factor family. Nat Rev Mol Cell Biol. 2001;2:827–37.
Epperson CN, Sammel MD, Bale TL, Kim DR, Conlin S, Scalice S, et al. Adverse childhood experiences and risk for first-episode major depression during the menopause transition. J Clin Psychiatry. 2017;78:e298–307.
Marshall AD. Developmental timing of trauma exposure relative to puberty and the nature of psychopathology among adolescent girls. J Am Acad Child Adolesc Psychiatry. 2016;55:25–32.e1.
Li H, Bowen A, Bowen R, Balbuena L, Feng C, Bally J, et al. Mood instability during pregnancy and postpartum: a systematic review. Arch Womens Ment Health. 2019. Mar. https://doi.org/10.1007/s00737-019-00956-6.
Stell BM, Brickley SG, Tang CY, Farrant M, Mody I. Neuroactive steroids reduce neuronal excitability by selectively enhancing tonic inhibition mediated by delta subunit-containing GABAA receptors. Proc Natl Acad Sci USA. 2003;100:14439–44.
Pusalkar M, Suri D, Kelkar A, Bhattacharya A, Galande S, Vaidya VA. Early stress evokes dysregulation of histone modifiers in the medial prefrontal cortex across the life span. Dev Psychobiol. 2016;58:198–210.
Kenworthy CA, Sengupta A, Luz SM, Ver Hoeve ES, Meda K, Bhatnagar S, et al. Social defeat induces changes in histone acetylation and expression of histone modifying enzymes in the ventral hippocampus, prefrontal cortex, and dorsal raphe nucleus. Neuroscience. 2014;264:88–98.
Provençal N, Arloth J, Cattaneo A, Anacker C, Cattane N, Wiechmann T, et al. Glucocorticoid exposure during hippocampal neurogenesis primes future stress response by inducing changes in DNA methylation. Proc Natl Acad Sci USA. 2019. Aug. https://doi.org/10.1073/pnas.1820842116.
Marrocco J, Gray JD, Kogan JF, Einhorn NR, O’Cinneide EM, Rubin TG, et al. Early Life stress restricts translational reactivity in CA3 neurons associated with altered stress responses in adulthood. Front Behav Neurosci. 2019;13:157.
Asimes A, Kim CK, Rao YS, Bartelt K, Pak TR. microRNA expression profiles in the ventral hippocampus during pubertal development and the impact of peri-pubertal binge alcohol exposure. Non-Coding RNA. 2019;5:21.
Lomniczi A, Wright H, Castellano JM, Matagne V, Toro CA, Ramaswamy S, et al. Epigenetic regulation of puberty via Zinc finger protein-mediated transcriptional repression. Nat Commun. 2015;6:10195.
Yang C, Ye J, Liu Y, Ding J, Liu H, Gao X, et al. Methylation pattern variation between goats and rats during the onset of puberty. Reprod Domest Anim Zuchthyg. 2018;53:793–800.
Yang C, Ye J, Li X, Gao X, Zhang K, Luo L, et al. DNA methylation patterns in the hypothalamus of female pubertal goats. PLoS ONE. 2016;11:e0165327.
Rowson SA, Bekhbat M, Kelly SD, Binder EB, Hyer MM, Shaw G, et al. Chronic adolescent stress sex-specifically alters the hippocampal transcriptome in adulthood. Neuropsychopharmacology. 2019;44:1207–15.
Li Z, Schulz MH, Look T, Begemann M, Zenke M, Costa IG. Identification of transcription factor binding sites using ATAC-seq. Genome Biol. 2019;20:45.
Golson ML, Kaestner KH. Fox transcription factors: from development to disease. Dev Camb Engl. 2016;143:4558–70.
Malarkey CS, Churchill MEA. The high mobility group box: the ultimate utility player of a cell. Trends Biochem Sci. 2012;37:553–62.
Bahrami S, Drabløs F. Gene regulation in the immediate-early response process. Adv Biol Regul. 2016;62:37–49.
Hantsoo L, Epperson CN. Anxiety disorders among women: a female lifespan approach. Focus Am Psychiatr Publ. 2017;15:162–72.
Bolea-Alamanac B, Bailey SJ, Lovick TA, Scheele D, Valentino R. Female psychopharmacology matters! Towards a sex-specific psychopharmacology. J Psychopharmacol (Oxf). 2018;32:125–33.
Maguire J, Mody I. GABAAR plasticity during pregnancy: relevance to postpartum depression. Neuron. 2008;59:207–13.
Melón LC, Hooper A, Yang X, Moss SJ, Maguire J. Inability to suppress the stress-induced activation of the HPA axis during the peripartum period engenders deficits in postpartum behaviors in mice. Psychoneuroendocrinology. 2018;90:182–93.
Melón L, Hammond R, Lewis M, Maguire J. A novel, synthetic, neuroactive steroid is effective at decreasing depression-like behaviors and improving maternal care in preclinical models of postpartum depression. Front Endocrinol. 2018;9:703.
Acknowledgements
We thank Eldin Jasarevic, Bridget Nugent, and Jennifer Chan for helpful input on these projects.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
About this article

Cite this article
Morrison, K.E., Cole, A.B., Kane, P.J. et al. Pubertal adversity alters chromatin dynamics and stress circuitry in the pregnant brain. Neuropsychopharmacol. 45, 1263–1271 (2020). https://doi.org/10.1038/s41386-020-0634-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41386-020-0634-y
This article is cited by
-
Chemogenetic activation of CRF neurons as a model of chronic stress produces sex-specific physiological and behavioral effects
Neuropsychopharmacology (2024)
-
Extracellular vesicles are dynamic regulators of maternal glucose homeostasis during pregnancy
Scientific Reports (2023)
-
In search of sex-related mediators of affective illness
Biology of Sex Differences (2021)
-
The composition of human vaginal microbiota transferred at birth affects offspring health in a mouse model
Nature Communications (2021)
