
Abstract
We have used aminoterminal pro C-type natriuretic peptide (NTproCNP)—a stable marker of CNP secretion—to study the effect of cortisol on CNP secretion and fetal growth. In ovine pregnancy, maternal plasma NTproCNP (largely sourced from the placenta) increases at the end of the first trimester and then decreases abruptly preterm during the phase of fetal surge in cortisol secretion. Postulating that increases in cortisol, as occurs in the fetal or maternal circulation in late pregnancy, will reduce CNP secretion, we studied the fetal and maternal responses in NTproCNP to sustained low-dose infusions of cortisol (1.2 mg/d/kg for 11 d) delivered to the fetus from d 117 gestation. Fetal plasma NTproCNP was progressively reduced during fetal cortisol infusions, whereas fetal girth growth was unchanged. In contrast, maternal NTproCNP was unaffected by cortisol. We conclude that fetal but not placental tissue production of CNP is reduced by small increments in fetal cortisol. Failure to reduce maternal NTproCNP may relate to the continuing presence of the placental barrier to cortisol at this stage of pregnancy.
Similar content being viewed by others
Main
C-type natriuretic peptide (CNP) is an important growth factor expressed in a wide range of tissues (1) during development (2) and postnatal life (3). Although shown to be essential to postnatal linear growth in both rodents (4) and humans (5), CNP production has also been found to correlate strongly with linear growth velocity in lambs and throughout all phases of linear growth in humans (6–8). In rapidly growing lambs, catabolic interventions such as caloric restriction (9) and high doses of glucocorticoids (7), well-recognized inhibitors of linear growth, rapidly and reversibly reduce circulating forms of CNP and markers of bone formation. Similar reductions in CNP forms occur in the fetus during maternal caloric restriction (10) or glucocorticoid administration (6). However, the effect of physiological increments in fetal cortisol on CNP synthesis is unknown.
Recent studies of ovine pregnancy show that both CNP and the bio inactive aminoterminal fragment of pro CNP 1–103 (NTproCNP) (11) are secreted by the placenta and circulate at high concentrations in maternal plasma from the end of the first trimester (12). In contrast, the ovine placenta does not seem to contribute to circulating fetal CNP concentrations, which are much lower than maternal concentrations (12). Nonetheless, as in the human fetus (13), fetal CNP synthesis is likely to be high, as shown by the very high concentrations of NTproCNP in fetal plasma. Fetal CNP forms are clearly regulated independently of maternal concentrations (10), but the sites of synthesis and the source of NTproCNP in the fetal circulation remain to be determined.
A striking feature of the pattern of maternal plasma CNP forms in healthy pregnant ewes is the sudden decline in both NTproCNP and CNP within the last week of pregnancy (12)—a phase of rapidly increasing fetal cortisol production (14). Mindful of the inhibitory effects of glucocorticoids on CNP synthesis and linear growth, we hypothesized that contrived increases in fetal cortisol within the physiological range in late gestation would inhibit not only fetal but also maternal concentrations of NTproCNP. Accordingly, we have studied the fetal response of NTproCNP to sustained low-dose infusions of cortisol delivered to the fetus during a 11-d period before the normal late gestation rise in fetal cortisol concentrations. The results show that fetal NTproCNP is reduced by small increments in fetal cortisol, whereas maternal NTproCNP is unaffected.
MATERIALS AND METHODS
Animals.
Romney ewes were synchronized with progesterone slow release devices (Eazi-Breed Controlled Intravaginal Drug Releasing device; Pharmacia and Upjohn, Kalamazoo, MI) before mating with Dorset rams. The pregnant ewes were scanned and only those carrying single fetuses were included. Ewes were housed in individual metabolic cages in visual contact with other ewes and acclimatized to laboratory conditions and concentrate feeds (Country Harvest Stock Feeds, Cambridge, NZ) for 7 d before surgery under halothane anesthesia at 111 ± 1 d of pregnancy (term 145 d). Strepcin (4-mL intramuscular injection; Stockguard Laboratories NZ Ltd, Hamilton, NZ) was administered to the ewe before surgery. Polyvinyl catheters were inserted into a maternal femoral artery and jugular vein and into fetal femoral arteries and veins via tarsal vessels. Growth catheters were inserted s.c. around each half of the fetal chest from the sternum to spine (15). Gentamicin (80 mg; Pharmacia and Upjohn) was administered to the amniotic cavity before closure of the uterus and maternal abdomen.
Animals were allowed at least 4 d to recover from surgery before the start of infusion studies. Growth catheters were measured twice daily, and the mean was used in the analyses. Approval for all phases of animal experimentation was obtained previously from the Animal Ethics Committee of the University of Auckland.
Experimental details.
Ewes (live weight range, 48–70 kg) were randomly assigned to one of two groups: controls (C, n = 12), which received i.v. saline infusion (0.5 mL/h), or fetal cortisol infusion group (F, n = 12), which received fetal i.v. infusion of hydrocortisone sodium succinate (1.2 mg hydrocortisone/d/kg fetus, 0.5 mL/h; Solu-Cortef, Pharmacia and Upjohn). Infusions started at 117 d of gestation with an initial 3 mL bolus of infusate and then a continuous infusion for the duration of the experiment (11 d). Doses were calculated according to fetal weight (3 kg) estimated from extensive historical data (16). Paired maternal and fetal arterial blood samples (3.5 mL) were collected between 0700 and 0900 h immediately before the start of infusions and on d 118, 121, 124, and 127. Animals were killed with an overdose of pentobarbitone at the end of the experiment, and the fetus and placenta were dissected, weighed, and measured.
Sample handling.
Blood samples were collected into sterile heparinized syringes, and aliquots were either frozen at −80°C or transferred to test tubes on ice and centrifuged at 4°C, and separated plasma was stored at −20°C for later analysis.
Assays.
Cortisol was measured by RIA with inhouse produced anticortisol rabbit sera (F1M1), after extraction of plasma with diethyl ether (17). The antibody was validated for sheep plasma and exhibited low cross reactivity with derivatives of cortisol (3.3% for 11-deoxycortisol, 0.18% for cortisone, 0.5% for corticosterone, 0.015% for progesterone, 0.002% for 11α-hydroxyprogesterone at 50% binding, no detectable cross reactivity for 17α-hydroxypregnenolone, pregnanediol, 21-deoxycortisone, aldosterone, cholesterol, or dexamethasone) (18). The assay had a minimal detectable dose of 0.44 ng/mL and intra- and interassay coefficients of variation of 9.8 and 10.8%, respectively.
NTproCNP was assayed as previously described (11), except that a more sensitive primary rabbit antiserum (J39) raised against NTproCNP (1–15) was used (50 μL 1:6000 diluted antiserum/assay tube). Peptide standards were made from synthetic human proCNP (1–19), taking into account the purity data supplied (Chiron Technologies, Victoria, Australia).Within- and between-assay coefficients of variation were 4.9 and 6.4%, respectively, at 22 pmol/L.
Statistical analyses.
All values are presented as mean ± SEM. Basal hormonal and metabolic data were analyzed by factorial ANOVA. Data obtained during the period of cortisol or saline infusion were analyzed by repeated measures ANOVA using baseline values (d 117) as a covariate and Bonferroni post hoc adjustment for multiple comparisons. Fetal girth increments were averaged for the two sides of the fetal chest and analyzed by multiple linear regression, with animal number nested within treatment group to account for repeated measurements in the same animal, and Tukey-Kramer adjustment for multiple comparisons (15). All statistical analyses were performed using SPSS (version 11.0; SPSS, Chicago, IL) and JMP (SAS Institute, Carey, NC).
RESULTS
Fetal plasma cortisol concentrations at the beginning of the experiment were similar in both groups and close to the lower limits of detection (Table 1). In saline-infused controls, fetal plasma cortisol concentrations increased slightly over the experimental period to a peak of 4.4 ± 1.0 ng/mL at the completion of the infusion. Fetal cortisol infusion increased fetal plasma cortisol concentrations 9-fold to a peak of 38 ± 14 ng/mL (p < 0.001, Fig. 1A).

Effects of infusions of cortisol (•, fetal delivery, n = 12) or saline (○, maternal delivery, n = 12) on changes in plasma cortisol from baseline ([Delta]) in fetal (A) and maternal (B) plasma. Infusions were delivered from d 117–127. Values are presented as mean ± SEM. Significant differences from control time-matched data are indicated by *p < 0.05 and **p < 0.001.
Maternal plasma cortisol concentrations at the beginning of the experiment were also similar in both groups (Table 1). In controls, maternal plasma cortisol concentrations tended to increase over the experimental period to a peak of 11.3 ± 2.7 ng/mL on d 127 (Fig. 1B). Fetal cortisol infusion did not significantly change maternal plasma cortisol concentrations.
Fetal plasma NTproCNP concentrations at the beginning of the experiment were similar in both groups (Table 1). Fetal cortisol infusion decreased fetal plasma NTproCNP concentrations (F = 2.9, p < 0.03), so that concentrations were 27% lower at the end of the infusion than at baseline, whereas concentrations were unchanged during saline infusions (Fig. 2A).

Effect of infusions of cortisol (•, fetal delivery, n = 12) or saline (○, maternal delivery, n = 12) on change in plasma NTproCNP from baseline ([Delta]) in fetal (A) and maternal (B) plasma. Infusions were delivered from d 117–127. Values are presented as mean ± SEM. Significant differences from control time-matched data are indicated by *p < 0.05.
Maternal plasma NTproCNP concentrations at the beginning of the experiment were similar in both groups and higher than fetal concentrations (Table 1). Maternal plasma NTproCNP concentrations did not change significantly during the experimental period in either group (Fig. 2B). Fetal growth, as measured by mean increment in fetal half-girth, was similar in both groups, and fetal weights were not significantly different at postmortem (Table 2).
DISCUSSION
This is the first study to show that plasma NTproCNP, a stable product of CNP gene expression, is progressively reduced by small increments in plasma cortisol in the fetal lamb. In contrast, maternal concentration of NTproCNP (largely of placental origin) is unaffected by cortisol in ewes for about 3–4 wk before term. These findings suggest that the inhibitory effect of glucocorticoids on CNP production is greater in fetal than in placental tissues at this stage of pregnancy.
Previous studies in 4-wk-old postnatal lambs indicate that CNP synthesis is rapidly and reversibly reduced by high doses of dexamethasone (7). Reduced CNP concentrations are also observed in umbilical blood of human newborns whose mothers were treated with glucocorticoids within 72 h of delivery (6). These findings confirm and extend these observations in showing significant inhibition of fetal CNP secretion by small increments in fetal plasma cortisol concentrations well within the range encountered in normal pregnancy.
Sites of CNP synthesis in the fetus remain to be determined, but clearly growing cartilage within growth plates of long bones is likely to be one important source (4,7). In postnatal life, there is increasing evidence that CNP synthesis within the growth plate or closely related tissue is strongly correlated with linear growth velocity (7). CNP is inhibited by catabolic interventions such as glucocorticoid administration (7) and caloric restriction (9), which also reduces circulating CNP forms in the fetal lamb (10). In this study, the increase in fetal plasma cortisol concentrations was much smaller than that normally associated with slowed fetal growth before birth (19) or demonstrated slow fetal growth earlier in gestation (20). Consistent with this, we found no changes in fetal growth, even when measured using the sensitive technique of daily changes in fetal girth (15,16). This was despite the fact that the percentage fall in plasma NTproCNP (27%) was similar to reductions (22–30%) that we observed in dexamethasone-treated postnatal lambs (7). These findings suggest that changes in growth are less tightly coupled with changes in CNP secretion in the fetus and are consistent with the predominant effect of the peptide on postnatal compared with skeletal growth before birth (2,4,21–24). Clearly, further studies are required to determine whether fall in CNP secretion is necessary rather than simply an associated feature of glucocorticoid-induced growth impairment in the fetus.
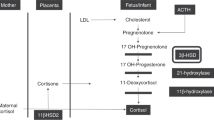
Our second hypothesis—that increasing fetal cortisol concentration would reduce maternal NTproCNP values (and therefore placental CNP secretion)—was not supported by this study. The hypothesis was based on our previous work showing that the dynamic fluctuations in maternal plasma CNP forms—including the preterm sudden decline—are strongly and positively associated with similar changes in the placental (cotyledon) CNP content (McNeill et al., The placenta is a major site of CNP synthesis during ovine pregnancy: differential contributions to maternal and fetal circulations, 2009 Endocrine Society's Annual Meeting June 10–13, 2009, Washington, DC, Abstract pp 3–251). Furthermore, we have recently localized CNP to the fetal placental binucleate cell (BNC), the population of which is known to be depleted rapidly in late pregnancy as fetal cortisol rises (25). These changes can be replicated by contrived increases in exogenous fetal cortisol concentrations (25). Although the placental concentration of CNP was not measured in this study, we conclude from these findings that in the face of unchanging maternal NTproCNP values during fetal cortisol infusion, the placental contribution to maternal levels was not significantly reduced in this study. Although it is possible that the sudden preterm decline in maternal plasma CNP forms is quite unrelated to the fetal cortisol surge, it is also possible that the lack of effect of fetal cortisol infusion on maternal plasma NTproCNP concentrations may be related to the timing of the cortisol infusions and the peak concentrations achieved, which were lower than those commonly reached close to term (14,20). A number of studies show that the site-specific expression of placental 11 beta hydroxysteroid dehydrogenase type 2, which is important in maintaining the placental barrier to cortisol (26), declines toward term, beginning from around d 125 in sheep (27). Thus, fetal cortisol infusion across d 117–127 may be too early to allow significant impact on the placental sites of CNP formation and release, despite being sufficient to reduce the production of CNP in other fetal tissues not expressing the enzyme. Clearly, studies using higher doses of cortisol extending later in gestation should resolve these issues.
In conclusion, fetal but not maternal plasma NTproCNP is reduced by small increments in fetal cortisol concentrations in late pregnancy. Reduced fetal concentrations of NTproCNP during fetal stress (10,13) or across parturition (6) are likely to be cortisol induced.
Abbreviations
- BNC:
-
binucleate cell
- CNP:
-
C-type natriuretic peptide
- NTproCNP:
-
aminoterminal pro C-type natriuretic peptide
References
Minamino N, Aburaya M, Kojima M, Miyamoto K, Kangawa K, Matsuo H 1993 Distribution of C-type natriuretic peptide and its messenger RNA in rat central nervous system and peripheral tissue. Biochem Biophys Res Commun 197: 326–335
Cameron VA, Aitken GD, Ellmers LJ, Kennedy MA, Espiner EA 1996 The sites of gene expression of atrial, brain, and C-type natriuretic peptides in mouse fetal development: temporal changes in embryos and placenta. Endocrinology 137: 817–824
Stepan H, Leitner E, Bader M, Walther T 2000 Organ-specific mRNA distribution of C-type natriuretic peptide in neonatal and adult mice. Regul Pept 95: 81–85
Chusho H, Tamura N, Ogawa Y, Yasoda A, Suda M, Miyazawa T, Nakamura K, Nakao K, Kurihara T, Komatsu Y, Itoh H, Tanaka K, Saito Y, Katsuki M 2001 Dwarfism and early death in mice lacking C-type natriuretic peptide. Proc Natl Acad Sci U S A 98: 4016–4021
Bartels CF, Bukulmez H, Padayatti P, Rhee DK, van Ravenswaaij-Arts C, Pauli RM, Mundlos S, Chitayat D, Shih LY, Al-Gazali LI, Kant S, Cole T, Morton J, Cormier-Daire V, Faivre L, Lees M, Kirk J, Mortier GR, Leroy J, Zabel B, Kim CA, Crow Y, Braverman NE, van den Akker F, Warman ML 2004 Mutations in the transmembrane natriuretic peptide receptor NPR-B impair skeletal growth and cause acromesomelic dysplasia, type Maroteaux. Am J Hum Genet 75: 27–34
Prickett TC, Dixon B, Frampton C, Yandle TG, Richards AM, Espiner EA, Darlow BA 2008 Plasma amino-terminal pro C-type natriuretic peptide in the neonate: relation to gestational age and postnatal linear growth. J Clin Endocrinol Metab 93: 225–232
Prickett TC, Lynn AM, Barrell GK, Darlow BA, Cameron VA, Espiner EA, Richards AM, Yandle TG 2005 Amino-terminal proCNP: a putative marker of cartilage activity in postnatal growth. Pediatr Res 58: 334–340
Olney RC, Prickett TC, Yandle TG, Espiner EA, Han JC, Mauras N 2007 Amino-terminal propeptide of C-type natriuretic peptide and linear growth in children: effects of puberty, testosterone and growth hormone. J Clin Endocrinol Metab 92: 4294–4298
Prickett TC, Barrell GK, Wellby M, Yandle TG, Richards AM, Espiner EA 2007 Response of plasma CNP forms to acute anabolic and catabolic interventions in growing lambs. Am J Physiol Endocrinol Metab 292: E1395–E1400
Prickett TC, Rumball CW, Buckley AJ, Bloomfield FH, Yandle TG, Harding JE, Espiner EA 2007 C-type natriuretic peptide forms in the ovine fetal and maternal circulations: evidence for independent regulation and reciprocal response to undernutrition. Endocrinology 148: 4015–4022
Prickett TC, Yandle TG, Nicholls MG, Espiner EA, Richards AM 2001 Identification of amino-terminal pro-C-type natriuretic peptide in human plasma. Biochem Biophys Res Commun 286: 513–517
McNeill BA, Barrell GK, Wellby M, Prickett TC, Yandle TG, Espiner EA 2009 C-type natriuretic peptide forms in pregnancy: maternal plasma profiles during ovine gestation correlate with placental and fetal maturation. Endocrinology 150: 4777–4783
Prickett TC, Kaaja RJ, Nicholls MG, Espiner EA, Richards AM, Yandle TG 2004 N-terminal pro-C-type natriuretic peptide, but not C-type natriuretic peptide, is greatly elevated in the fetal circulation. Clin Sci (Lond) 106: 535–540
Fowden AL, Mundy L, Silver M 1998 Developmental regulation of glucogenesis in the sheep fetus during late gestation. J Physiol 508: 937–947
Harding JE 1997 Prior growth rate determines the fetal growth response to acute maternal undernutrition in fetal sheep of late gestation. Prenat Neonatal Med 2: 300–309
Rumball CW, Harding JE, Oliver MH, Bloomfield FH 2008 Effects of twin pregnancy and periconceptional undernutrition on maternal metabolism, fetal growth and glucose-insulin axis function in ovine pregnancy. J Physiol 586: 1399–1411
Fraser M, Matthews SG, Braems G, Jeffray T, Challis JR 1997 Developmental regulation of preproenkephalin (PENK) gene expression in the adrenal gland of the ovine fetus and newborn lamb: effects of hypoxemia and exogenous cortisol infusion. J Endocrinol 155: 143–149
Davidson JO, Fraser M, Naylor AS, Roelfsema V, Gunn AJ, Bennet L 2008 Effect of cerebral hypothermia on cortisol and adrenocorticotropic hormone responses after umbilical cord occlusion in preterm fetal sheep. Pediatr Res 63: 51–55
Fowden AL, Li J, Forhead AJ 1998 Glucocorticoids and the preparation for life after birth: are there long-term consequences of the life insurance?. Proc Nutr Soc 57: 113–122
Fowden AL, Szemere J, Hughes P, Gilmour RS, Forhead AJ 1996 The effects of cortisol on the growth rate of the sheep fetus during late gestation. J Endocrinol 151: 97–105
Tamura N, Doolittle LK, Hammer RE, Shelton JM, Richardson JA, Garbers DL 2004 Critical roles of the guanylyl cyclase B receptor in endochondral ossification and development of female reproductive organs. Proc Natl Acad Sci U S A 101: 17300–17305
Tsuji T, Kondo E, Yasoda A, Inamoto M, Kiyosu C, Nakao K, Kunieda T 2008 Hypomorphic mutation in mouse Nppc gene causes retarded bone growth due to impaired endochondral ossification. Biochem Biophys Res Commun 376: 186–190
Sogawa C, Tsuji T, Shinkai Y, Katayama K, Kunieda T 2007 Short-limbed dwarfism: slw is a new allele of Npr2 causing chondrodysplasia. J Hered 98: 575–580
Hachiya R, Ohashi Y, Kamei Y, Suganami T, Mochizuki H, Mitsui N, Saitoh M, Sakuragi M, Nishimura G, Ohashi H, Hasegawa T, Ogawa Y 2007 Intact kinase homology domain of natriuretic peptide receptor-B is essential for skeletal development. J Clin Endocrinol Metab 92: 4009–4014
Ward JW, Wooding FB, Fowden AL 2002 The effects of cortisol on the binucleate cell population in the ovine placenta during late gestation. Placenta 23: 451–458
Waddell BJ, Benediktsson R, Brown RW, Seckl JR 1998 Tissue-specific messenger ribonucleic acid expression of 11beta-hydroxysteroid dehydrogenase types 1 and 2 and the glucocorticoid receptor within rat placenta suggests exquisite local control of glucocorticoid action. Endocrinology 139: 1517–1523
Clarke KA, Ward JW, Forhead AJ, Giussani DA, Fowden AL 2002 Regulation of 11 beta-hydroxysteroid dehydrogenase type 2 activity in ovine placenta by fetal cortisol. J Endocrinol 172: 527–534
Acknowledgements
We thank Janine Street for the assays and members of the Fetal and Neonatal Physiology group at the Liggins Institute for their expert technical assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
Supported, in part, by the Health Research Council of New Zealand and the National Research Centre for Growth and Development.
Rights and permissions
About this article
Cite this article
Prickett, T., McNeill, B., Oliver, M. et al. Effect of Cortisol on C-Type Natriuretic Peptide in Ovine Pregnancy: Differential Responses in Fetal and Placental Tissues. Pediatr Res 68, 462–465 (2010). https://doi.org/10.1203/PDR.0b013e3181f9039d
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/PDR.0b013e3181f9039d
This article is cited by
-
Acute inflammation in young children inhibits C-type natriuretic peptide
Pediatric Research (2013)
