
Abstract
We investigated the concentrations of serum inhibin B and the pro-αC-containing inhibins (pro-αC inhibin) and their relations to serum FSH levels in 38 healthy boys during their progression through puberty. Furthermore, we studied the effect of recombinant human FSH (rhFSH) on pro-αC inhibin production in three prepubertal gonadotropin-deficient boys. The serum inhibin B level increased between Tanner stages G1 and G2 (p < 0.02), simultaneously with the serum LH and testosterone concentrations. In advanced puberty, inhibin B changed less, and at stage G4, correlated negatively with serum FSH level (r = -0.57, p < 0.001, n = 37). The serum pro-αC inhibin level changed differently and increased also in advanced puberty. In prepubertal healthy subjects, the serum pro-αC inhibin and FSH levels correlated positively (r = 0.50, p = 0.051, n = 16), and during the rhFSH treatment of gonadotropin-deficient boys, serum pro-αC inhibin levels increased. These findings suggest that an increased production of inhibin B is an early event in puberty and that inhibin B can inhibin pituitary FSH secretion in the course of sexual maturation. Furthermore, they suggest that during puberty free inhibin (pro)-α-subunits, detected by the pro-αC inhibin assay, are present in large amounts and that FSH stimulates their production by prepubertal testes.
Similar content being viewed by others

Main
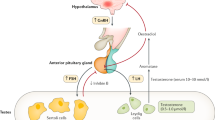
Secretion of FSH is regulated at the hypothalamic level by sex steroids and at the pituitary level by inhibin, a glycoprotein with two dissimilar subunits (1). Several recent findings suggest that inhibin B, consisting of α- and βB-subunits, is the endocrinologically most important form of inhibin in men. The serum inhibin B level increases in response to rhFSH (2,3), whereas the level is low in men with primary or secondary hypogonadism (2,4,5), suggesting that the amount of circulating inhibin B reflects the function of the Sertoli cells. Unequivocal evidence that inhibin B can suppress pituitary FSH secretion is lacking. However, a negative feedback appears likely, because in normal and infertile men, in men with primary or secondary hypogonadism, and in boys during puberty, serum inhibin B and FSH levels correlate negatively (4–9).
In addition to inhibin B, men have circulating inhibin precursor forms and free inhibin α-subunits, but the physiologic relevance of these proteins is unclear. Even in men without testes, the serum level of inhibin precursors that contain propeptide in the α-subunit (pro-αC inhibin) is measurable (2). The pro-αC inhibin assay could also detect dimeric inhibin precursor forms that contain the propeptide (10), but in men, it has been suggested to be unique in measuring circulating free inhibin (pro)-α-subunits (4,7,10).
We describe here the patterns of change in serum inhibin B and pro-αC inhibin levels during normal male development, and relate these findings to previously described parameters of puberty. Moreover, to further evaluate the role of FSH in the production of pro-αC inhibin in very early puberty, we studied serum pro-αC inhibin levels in boys with hypogonadotropic hypogonadism before and during their treatment with rhFSH.
METHODS
Subjects. The study material consisted of a longitudinal series of data gathered from an earlier follow-up study of healthy Finnish schoolboys (11). At the beginning of the study, the age (mean ± SEM) of the 38 boys studied was 11.7 ± 0.1 y. The subjects were healthy, apart from one boy in whom epilepsy was diagnosed and treated with oxcarbazepine medication. In short, the study covered 3 y and consisted of nine visits at 3-mo intervals during the first 2 y, the final check-up taking place 1 y later. At every visit, puberty was staged according to Tanner (12), and the length and width of the testes were measured with a ruler to the nearest millimeter. Testis volume was calculated from the formula 0.52 × length × width2 and converted to milliliters (13).
The study protocol was approved by the parents, the school authorities, and the ethics committee of the Hospital for Children and Adolescents, University of Helsinki.
Standard laboratory analyses. At every visit, venous blood samples were drawn from all the boys between 0830 and 1400 h. After clotting, the serum was separated by centrifugation and stored at -20°C until required. The serum testosterone concentration was measured by RIA after separation of the steroid fractions on a Lipidex-5000 microcolumn (Packard-Becker, B.V. Chemical Operations, Groningen, The Netherlands) as described previously (14). Serum LH and FSH concentrations were measured by time-resolved immunofluorometric assays, using reagents from Wallac OY (Turku, Finland), as described previously (15). Serum sex hormone-binding globulin was determined by immunofluorometric assays (DELFIA, Wallac, Turku, Finland). The free androgen index was calculated as serum testosterone/sex hormone-binding globulin × 100 (16).
Inhibin B and pro-αC inhibin assays. Serum inhibin B concentration was determined as described in detail previously (7,17,18). The sensitivity of the assay was 10 pg/mL, there being a 0.5% cross-reaction with inhibin A and 0.1% cross-reactions with human pro-αC precursor, activin A, activin B, and follistatin. The within- and between-plate coefficients of variation were less than 10%. Serum pro-αC inhibin concentrations were determined as previously described (19). The sensitivity of this assay was 3 pg/mL. Recombinant forms of activin A, activin B, and follistatin all showed cross-reactions of less than 0.02%. The within- and between-plate variations were less than 5 and 7%, respectively. All measurements were carried out in duplicate.
Statistics. For analyses of serum inhibin B and pro-αC inhibin concentrations at each Tanner's stage, the median value within each stage was calculated for each subject. In cases with an even number of observations for a stage, one of the two central observations was randomly selected. Spearman's rank correlation coefficients between serum inhibin B, pro-αC inhibin, and FSH levels were calculated for the boys at different Tanner's genital stages, to avoid the effect of nonlinear relationships between these variables. To examine the significance of the differences between mean serum hormone and pro-αC inhibin levels in boys at different Tanner stages, we used ANOVA, followed by Fisher's protected least significant difference test. Before ANOVA, a log transformation was applied to the serum hormone concentrations, as the variances and means increased simultaneously. Thereafter, the variances of the transformed variables at the different stages did not differ. All values are expressed as geometric means (±SD), unless stated otherwise. Statistical significance was accepted for p < 0.05.
RESULTS
The associations between serum FSH and inhibin B levels in boys at the different Tanner genital stages are presented in Fig. 1. In boys at stage G4, serum FSH and inhibin B levels correlated significantly (r = -0.57, p < 0.001, n = 37; Fig. 1). Individual patterns of serum inhibin B and FSH levels in six boys with similar rates of pubertal progression are presented in Fig. 2. The development of the inverse relationship between serum inhibin B and FSH levels is less obvious in boy 5, recognizable in boy 1, but can be particularly well seen in boys 2, 3, 4, and 6 (Fig. 2). The distributions of the serum inhibin levels and other parameters investigated in boys at different Tanner stages are given in Table 1. In boys at each stage, serum inhibin B and pro-αC inhibin levels did not correlate (r = -0.10-0.10).

Association of FSH with inhibin B in healthy boys at different Tanner genital stages (from G1 to G4) during the 3-y follow-up. There were observations made on 16 subjects in stage G1 and on 37 subjects in stages G2, G3, and G4.

Individual serum (S) inhibin B and FSH concentrations in six boys during the 3-y follow-up. These boys were selected on the basis of similar tempo of puberty; they were followed up from Tanner stage G1 to stage G4, and during that time, had comparable testicular growth rates (range, 4.2-5.4 mL/y).
Because the numbers of boys in each Tanner stage differed, the longitudinal changes in the parameters examined were studied in the 16 boys who entered the study at stage G1, and were followed up until stage G4. Their mean serum inhibin B concentration increased from 109 (90-131) pg/mL at stage G1 to 134 (99-181) pg/mL at stage G4 (Fig. 3), the only significant change occurring between stages G1 and G2 (p < 0.02), after which there was a plateau. From G1 to G2, the mean serum FSH concentration did not change significantly (p = 0.18), but the mean testis volume (p < 0.001), and mean serum LH and testosterone concentrations were elevated (p < 0.001 and p < 0.01, respectively, Fig. 3). In this same subgroup of boys, the mean serum pro-αC inhibin level increased, but did not show the plateau in mid-puberty (Fig. 3). In contrast, the only statistically significant change between successive stages was from G3 to G4 (p < 0.05).

Distributions of serum (S) inhibin B (upper left panel), pro-αC inhibin (upper right panel), gonadotropin (middle panels), and testosterone (lower left panel) levels, and testis volumes (lower right panel) in 16 subjects followed up from Tanner stage G1 to stage G4. The 10th, 25th, 50th, 75th, and 90th percentiles are shown.
Serum pro-αC inhibin levels in boys at the earliest stages of puberty (G1 and G2) were not associated with age, but in boys at stage G1, the levels correlated positively with serum FSH concentrations (r = 0.50, p = 0.051, n = 16, Fig. 4, left panel). To clarify the causality behind this finding, we examined the serum pro-αC inhibin levels in three prepubertal (G1P1) gonadotropin-deficient boys from an earlier study (3). These boys (A, B, and C), were treated with rhFSH (1.5 IU/kg s.c. three times/wk) for 1 y from ages 12.8, 13.2 and 13.2 y, respectively. During the 12-mo treatment period, serum pro-αC inhibin levels, measured at 3-mo intervals, increased (p < 0.02, ANOVA). The values before and during the treatment are presented in Fig. 4 (right panel).

(Left panel) Association of pro-αC inhibin with FSH in healthy boys (n = 16) at Tanner stage G1. (Right panel) Serum (S) pro-αC inhibin concentrations in three prepubertal gonadotropin-deficient boys from an earlier study (3), treated with recombinant human FSH (1.5 IU/kg three times/wk s.c.) for 1 y. Serum levels before (first data point for each subject), and measured with 3-mo intervals during the treatment are shown for boys A (○), B (▪), and C (▵).
DISCUSSION
The recent development of specific immunoassays for measuring the different inhibin forms in human plasma and serum has significantly improved the understanding of the signaling between the pituitary gland and the gonads. The present study was undertaken to investigate the changes occurring in serum gonadotropin and inhibin levels during pubertal development of healthy boys. We found that, in prepubertal boys, FSH is likely to regulate the production of pro-αC inhibin, but in the course of puberty, the most dramatic findings were observed between serum FSH and inhibin B levels.
In early puberty, simultaneously with elevations in serum LH and testosterone levels, the mean serum inhibin B concentration increased. The relative roles of LH and FSH in this process are unclear. Although a significant production of inhibin B by the Leydig cells is unlikely (4), the LH-induced androgen production may indirectly stimulate inhibin B secretion from the Sertoli cells by, for example, enhancing the interaction between the Sertoli cells and germ cells (7). On the other hand, rhFSH appears to stimulate inhibin B production by immature testes (3), but in the present study, serum inhibin B and FSH levels in healthy prepubertal boys were not positively correlated. Nevertheless, the temporal relationship between FSH and inhibin B secretion is not known, and therefore, this lack of positive association in healthy prepubertal subjects does not rule out FSH-induced secretion of inhibin B.
After the early increase in serum inhibin B level, it remained relatively unchanged and correlated negatively with serum FSH level, suggesting, in agreement with the results of Crofton et al. (9), an establishment of a feedback loop between testicular inhibin B and pituitary FSH secretion. A significant correlation was observed relatively late, in boys at Tanner stage G4. However, an earlier feedback regulation has been proposed (9) and is further suggested by our previous finding that pre- and early pubertal boys with gonadal failure have higher serum FSH levels than do normal subjects (20).
In men, as evidenced by immunologic characterization of adult male serum (10), and suggested by the lack of correlation between serum inhibin B and pro-αC inhibin levels (4,7), the pro-αC assay mainly measures the amount of circulating free inhibin (pro)-α-subunits and not the inhibin B precursor forms (containing the propeptide in the α-subunit). In the present study, serum inhibin B and pro-αC inhibin levels were not correlated and displayed different patterns of change during puberty. These findings suggest that also in boys the pro-αC inhibin assay measures the amount of free inhibin (pro)-α-subunits. These proteins have different potential sources (2,4,21), which can explain the wide distributions of the serum pro-αC inhibin levels observed in boys at different Tanner stages. Nevertheless, in healthy prepubertal boys, serum pro-αC inhibin and FSH levels correlated, and during the rhFSH treatment of prepubertal gonadotropin-deficient boys, the serum pro-αC inhibin level increased. These findings suggest that FSH is an important factor stimulating pro-αC inhibin secretion by immature testes. Parenthetically, the rhFSH treatment related increase could also have been a normal age-related phenomenon. This was not, however, supported by the finding that in healthy pre- and early pubertal boys, serum pro-αC inhibin levels did not correlate with age.
The different precursor forms of free inhibin α-subunits are proposed to inhibit the binding of FSH to its receptor (22). However, there is no unequivocal evidence for any distinct endocrine function of these proteins in men (7,23), which is in agreement with our results obtained by studying boys during puberty. Nevertheless, our results suggest that if the serum inhibin level in boys is measured by an assay not specific for dimeric inhibin B, the presence of free inhibin (pro)-α-subunits in large amounts can potentially explain the lack of negative association between serum FSH and immunoactive inhibin levels (24).
In conclusion, the present work demonstrates that serum inhibin B level increases in early puberty, plateaus thereafter, and correlates negatively with the serum FSH level, supporting the view that, in healthy boys during puberty, pituitary FSH secretion is inhibited by inhibin B. During puberty, serum pro-αC inhibin level increases differently, displays wide variation, and is not negatively correlated with serum gonadotropin levels, suggesting that, in boys, the pro-αC inhibin assay measures mainly the serum level of free circulating inhibin (pro)-α-subunits.
Abbreviations
- rhFSH:
-
recombinant human FSH
- G1-G5:
-
Tanner genital stages 1-5
REFERENCES
Burger HG, Igarashi M 1988 Inhibin: definition and nomenclature, including related substances. J Clin Endocrinol Metab 66: 885–886
Anawalt BD, Bebb RA, Matsumoto AM, Groome NP, Illingworth PJ, McNeilly AS, Bremner, WJ 1996 Serum inhibin B levels reflect Sertoli cell function in normal men and men with testicular dysfunction. J Clin Endocrinol Metab 81: 3341–3345
Raivio T, Toppari J, Perheentupa A, McNeilly AS, Dunkel L 1997 Treatment of prepubertal gonadotrophin-deficient boys with recombinant human follicle-stimulating hormone. Lancet 350: 263–264
Wallace EM, Groome NP, Riley SC, Parker AC, Wu FCW 1997 Effects of chemotherapy-induced testicular damage on inhibin, gonadotropin, and testosterone secretion: a longitudinal prospective study. J Clin Endocrinol Metab 82: 3111–3115
Nachtigal LB, Boepple PA, Seminara SB, Khoury RH, Sluss PM, Lecain AE, Crowley WF Jr 1996 Inhibin B secretion in males with gonadotropin-releasing hormone (GnRH) deficiency before and during long-term GnRH replacement: relationship to spontaneous puberty, testicular volume, and prior treatment-a clinical research center study. J Clin Endocrinol Metab 81: 3520–3525
Andersson RA, Wallace EM, Groome NP, Bellis AJ, Wu FWC 1997 Physiological relationships between inhibin B, follicle stimulating hormone secretion and spermatogenesis in normal men, and response to gonadotrophin suppression by exogenous testosterone. Hum Reprod 12: 746–751
Illingworth PJ, Groome NP, Byrd W, Rainey WE, McNeilly AS, Mather JP, Bremmer WJ 1996 Inhibin B: a likely candidate for the physiologically important form of inhibin in men. J Clin Endocrinol Metab 81: 1321–1325
Seminara S, Boepple PA, Nachtigal LB, Pralong FP, Khoury RH, Sluss PM, Lecain AE, Crowley WF Jr 1996 Inhibin B in males with gonadotropin-releasing hormone (GnRH) deficiency: changes in serum concentration after short term physiologic GnRH replacement a clinical research center study. J Clin Endocrinol Metab 81: 3692–3696
Crofton PM, Illingworth PJ, Groome NP, Stirling HF, Swanston I, Gow S, Wu FC, McNeilly A, Kelnar CJ 1997 Changes in dimeric inhibin A and B during normal early puberty in boys and girls. Clin Endocrinol 46: 109–114
Robertson DM, Cahir N, Findlay JK, Burger HG, Groome N 1997 The biological and immunological characterization of inhibin A and B forms in human follicular fluid and plasma. J Clin Endocrinol Metab 82: 889–896
Anttila R, Koistinen R, Seppala M, Koistinen H, Siimes MA 1994 Insulin-like growth factor I and insulin-like growth factor binding protein 3 as determinants of blood hemoglobin concentration in healthy subjects. Pediatr Res 36: 745–748
Tanner JM 1962 Growth at adolescence. Blackwell, Oxford, UK, 22–29.
Hansen P, With T 1952 Clinical measurement of testes. Acta Med Scand 206( suppl): 457–465
Apter D, Jänne O, Karvonen P, Vihko R 1976 Simultaneous determination of five sex hormones in human serum by radioimmunoassay after chromatography on Lipidex-5000. Clin Chem 22: 32–38
Dunkel L, Alfthan H, Stenman U-H, Perheentupa J 1990 Gonadal control of pulsatile secretion of luteinizing hormone and follicle stimulating hormone in prepubertal boys evaluated by ultrasensitive time-resolved immunofluorometric assays. J Clin Endocrinol Metab 70: 107–114
Selby C 1990 Sex hormone binding: origin, function and clinical significance. Ann Clin Biochem 27: 532–541
Illingworth P, Groome NP, Duncan C, Grant V, Tovanabutra S, Baird DT, McNeilly AS 1996 Measurement of circulating inhibin forms during the establishment of pregnancy. J Clin Endocrinol Metab 81: 1471–1475
Groome NP, Illingworth PJ, O'Brien M, Pai R, Rodger FE, Mather JP, McNeilly AS 1996 Measurement of dimeric inhibin B throughout the human menstrual cycle. J Clin Endocrinol Metab 81: 1401–1405
Groome NP, Illingworth PJ, O'Brien M, Priddle J, Weaver K, McNeilly AS 1995 Quantification of inhibin pro-αC-containing forms in human serum by a new ultra-sensitive two site enzyme linked immunosorbent assay. J Clin Endocrinol Metab 80: 2926–2932
Dunkel L, Siimes MA, Bremner WJ 1993 Reduced inhibin and elevated gonadotropin levels in early pubertal boys with testicular defects. Pediatr Res 33: 514–518
McNeilly AS, Brooks AN, Baxter G, Webb R 1994 Sheep adrenal inhibin. In: Burger HG, Findlay K, Robertson DM, de Kretser D, Petraglia F (eds) Inhibin and Inhibin-Related Proteins. Ares-Serono Symposia, Rome, 261–269.
Schneyer AL, Sluss PM, Whitcomb RW, Martin KA, Sprengel R, Crowley WF Jr 1991 Precursors of α-inhibin modulate follicle-stimulating hormone receptor binding and biological activity. Endocrinology 129: 1987–1999
Lambert-Messerlian GM, Crowley WF Jr, Schneyer AL 1995 Extragonadal α-inhibin precursor proteins circulate in human male serum. J Clin Endocrinol Metab 80: 3043–3049
Burger HG, McLachlan RI, Bangah M, Quigg H, Findlay JK, Robertson DM, de Kretser DM, Warne GL, Werther GA, Hudson IL, Cook JJ, Fiedler R, Greco S, Yong ABW, Smith P 1988 Serum inhibin concentrations rise throughout normal male and female puberty. J Clin Endocrinol Metab 67: 689–694
Author information
Authors and Affiliations
Additional information
Supported financially by the Foundation for Paediatric Research, Helsinki, Finland.
Rights and permissions
About this article
Cite this article
Raivio, T., Perheentupa, A., McNeilly, A. et al. Biphasic Increase in Serum Inhibin B during Puberty: A Longitudinal Study of Healthy Finnish Boys. Pediatr Res 44, 552–556 (1998). https://doi.org/10.1203/00006450-199810000-00014
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-199810000-00014
This article is cited by
-
Reactive oxygen species and male reproductive hormones
Reproductive Biology and Endocrinology (2018)
