
Abstract
Previous studies have shown that the expression of cardiac angiotensin II(ANG II) type 1 (AT1) and type 2 (AT2) receptors are developmentally regulated, although factors modulating these receptors have not been well investigated. The present study was designed 1) to characterize the ontogeny of cardiac AT1 and AT2 gene expression during the last third trimester of gestation in fetal sheep and newborn lambs, 2) to determine the influence of ANG II on modulating cardiac AT1 and AT2 gene expression during fetal life, and 3) to investigate the role of AT1 receptor activity on the regulation of AT1 and AT2 mRNA levels during fetal cardiac development. Using sheep AT1 and AT2 cDNA probes, we demonstrated that cardiac AT1 gene expression is relatively unchanged during fetal (90-135 d of gestation, term 145 d) and newborn life. In contrast, cardiac AT2 mRNA expression was high during fetal development and decreased rapidly after birth. Continuous i.v. infusion of ANG II (9.5 nM/h) for 24 h, which raised ANG II levels from 84± 9 to 210 ± 21 pg/mL had no effect on the expression of cardiac AT1 or AT2 mRNA, but increased adrenal and decreased liver AT1 mRNA levels. Administration of the AT1 receptor antagonist losartan (1.2 mg kg-1 h-1) significantly decreased arterial blood pressure in fetuses at 110- and 135-d, but not 95-d gestation. Except for increased AT1 receptor gene expression in the right atrium at 95-and 135-d gestation, and left ventricle at 110-d gestation, cardiac AT1 and AT2 mRNA levels were unaltered by AT1 receptor blockade. In summary, this study demonstrates that cardiac AT2 but not AT1 receptor gene expression is regulated by the transition from fetal to newborn life. Neither ANG II nor blockade of AT1 receptors significantly alter the expression of AT1 or AT2 mRNA in the fetal heart. Endogenous ANG II also appears to significantly contribute to the maintenance of blood pressure homeostasis during the final third of gestation in fetal lambs.
Similar content being viewed by others
Main
Studies in newborn animals and human infants have shown that the activity of the RAS increases at birth(1,2). This increased activity has previously been shown to be important in regulating several of the cardiovascular and autonomic adaptations which occur during the transition from fetal to newborn life(1,3). Recently, it has been suggested that the RAS also exerts a major influence of cellular growth and organ differentiation(4,5). Accumulating evidence indicates that, during neonatal and adult life, ANG II has a growth-promoting effect on the heart, including stimulating protein synthesis and cellular hypertrophy and inducing growth promoting genes in cardiac myocytes(5–7).
ANG II exerts its effects via specific receptors, classified as type 1(AT1) and type 2 (AT2) based on selective antagonism by peptidic and nonpeptidic ligands(8,9). There appear to be important developmental changes in the distribution and expression of the two distinct ANG II receptors. In developing rats, both in situ hybridization and autoradiographic studies have shown that AT2 receptor expression is high in developing embryos, is localized to fetal mesenchymal tissues, and decreases late in development and with postnatal maturation(4,10). On the other hand, AT1 receptors appear later in fetal development and are localized primarily to areas related to blood pressure regulation and fluid homeostasis(4,10,11). Despite the apparent importance of the ANG II in maintaining physiologic homeostasis early in life, few studies have investigated the factors regulating developmental changes in the expression of cardiac ANG II receptor genes. These changes may be important in regulating growth and functional maturation of the heart(5). The present study was therefore designed to characterize the ontogeny of cardiac AT1 and AT2 gene expression in the developing ovine heart and to determine the influence of ANG II and AT1 receptor blockade in modulating cardiac AT1 and AT2 gene expression during the last trimester of gestation.
METHODS
Animal preparation and surgical procedures. Studies were performed on fetal and newborn sheep of Dorset and Suffolk mixed breeding, obtained from a local source. Gestational ages of the fetuses were based on the induced ovulation technique(12). Fetal weight was estimated according to the following formula: weight (kg) = 0.0961 × gestational age (d) - 9.2228, r = 0.85(13). All procedures were performed within the regulations of the Animal Welfare Act and the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and were approved by the University of Iowa Animal Care and Use Committee.
Surgical preparation and experimental protocols. In the first series of studies, we determined maturational changes in cardiac AT1 and AT2 mRNA levels during fetal development and the early postnatal period in sheep. For these studies, pregnant ewes were anesthetized with 12 mg/kg thiopental sodium (Abbot Laboratories) and maintained with a mixture of halothane (1%), oxygen (33%), and nitrous oxide (66%). Under sterile conditions, the uterus was exteriorized and the fetus was exposed. Samples of fetal cardiac left and right ventricular free wall and left and right atrium were obtained at 90, 115, and 135 d of gestation (term 145 d). Similar procedures were performed in anesthetized lambs at 2 d, 7 d, and 8 wk of life(n = 6-12 animals for each group). Because expression of ANG II receptors and mRNA may differ within different regions of the ventricle, tissues from only left and right ventricle apical free walls were used for mRNA analysis in this and subsequent studies. After being removed, the cardiac tissue was snap frozen in liquid nitrogen and stored at -70°C.
In a second series of studies, we investigated the effects of exogenous ANG II on the expression of fetal left and right ventricle AT1 and AT2 mRNAs in nine pairs of chronically instrumented twin fetal sheep at 129 ± 2 d of gestation. For these studies, the ewe was placed under general anesthesia as described above, the uterus was opened, and polyethylene catheters were placed into the femoral arteries and veins of both twin fetuses. Fetal incisions were closed, the fetus was returned to the uterus, and all surgical incisions were closed. Catheters were exteriorized through a s.c. tunnel and placed in a cloth pouch on the ewe's flank. Ampicillin sodium (Wyeth Laboratories, Philadelphia, PA) was administered to the ewe intramuscularly (2 g) before surgery and every 24 h after surgery for three doses, and infused into the amniotic cavity (2 g) immediately after surgery. Animals were returned to individual pens and allowed free access to food and water.
After a 72-h recovery period from surgery, one of each set of twins was administered a continuous i.v. infusion of ANG II (Sigma Chemical Co., St. Louis, MO) at a rate of 10 µg/h (9.5 nM/h) delivered in a volume at 1 mL/h (3.2 ± 0.1 µg kg-1 min-1). The other twin served received 0.9% NaCl at 1 mL/h (vehicle for ANG II) and served as a control. Because, in sheep, there are virtually no vascular anastomoses between twin placentae(14), an exchange of hormone is unlikely. The infusions were carried out with a portable peristaltic infusion pump (Cormed, Inc., Middleport, NY) secured on the back of the ewe in a specially designed jacket that allowed the animals to move freely during the infusion. Fetal arterial blood pressure and heart rate and amniotic pressure were monitored in each fetus for 30 min before and at the end of the 24-h infusion of ANG II. Fetal arterial blood was sampled before and 24 h after starting the ANG II infusion for determination of arterial pH, PO2, PCO2, hematocrit, PRA, and plasma ANG II concentration. At the end of the 24-h infusion, the ewes were again anesthetized, and the fetal heart was removed, and the left and right ventricle free wall was isolated, snap frozen in liquid nitrogen, and stored at -70°C.
In a third series of studies, we investigated the effects of the selective AT1 receptor antagonist, losartan (a generous gift of DuPont/Merck, Wilmington, DE), on fetal cardiac AT1 and AT2 gene expression in twin fetal sheep at 95, 110, and 135 d of gestation (n = 7-9 at each gestational age). The surgical preparation was identical to that described for the second series of studies. After a 72-h recovery period from surgery, one of each set of twin received a continuous i.v. infusion of losartan (10 mg/kg bolus followed by 20 µg kg-1 min-1, 1 mL kg-1 h-1) for 48 h. The other twin served as a saline-treated control. Fetal arterial blood pressure, fetal heart rate, and amniotic pressure were monitored in each fetus for 30 min before and at the end of the 48-h losartan infusion. Fetal arterial blood was obtained from each fetus before and 48 h after starting the losartan infusion for determination of arterial pH, PO2, PCO2, hematocrit, and plasma ANG II concentration. At the end of the 48-h infusion, the ewes were anesthetized, the fetal heart was removed, and the left and right ventricle free wall was isolated, snap frozen in liquid nitrogen, and stored at -70°C.
Analytical procedures. Arterial blood for pH, PCO2, and PO2 was collected anaerobically in heparinized syringes, and measurements were immediately determined using an IL-1303 pH blood gas analyzer (Laboratory Instruments, Schaumburg, IL) at 39.5°C. Hematocrit was determined using a meter caliper. Measurements of plasma ANG II concentration, and PRA were performed by RIA (University of Iowa Cardiovascular Center RIA Core Facility, Donna B. Farley, Director).
Isolation of RNA and preparation of probes. Total cellular RNA was isolated using TRI-REAGENT (Molecular Research Center, Cincinnati, OH). RNA was quantified spectrophotometrically by absorbance at 260 nm. RNA samples were stored as an ethanol precipitate at -70°C until further analysis.
The sheep AT1 receptor partial cDNA sequence from nucleotide 114-783 and AT2 receptor partial cDNA sequence from nucleotide 142-921 were isolated as previously described(15,16). The plasmids pAT1 and pAT2 were then treated with the restriction enzymes NdeI and HindIII, respectively, and purified by phenol extraction and ethanol precipitation. The resultant linear plasmids were used to generate labeled antisense RNA probes using T7 RNA polymerase (U.S. Biochemical Corp., Cleveland, OH) and[δ-32P]UTP (Amersham Corp, Arlington Heights, IL).
An 18 S rRNA probe was used to confirm equal loading and transfer of RNA. The 18 S rRNA probe was prepared from an 18 S cDNA clone corresponding to a 82-bp fragment of a highly conserved region of the human 18 S rDNA gene(Ambion Inc., Austin, TX).
Northern blot hybridization. Aliquots of 10 µg of RNA as measured by absorbance at 260 nm were fractioned by 1% formaldehyde-agarose gel electrophoresis. After electrophoresis, RNA was transferred to a 0.45-mm Nytran filter. The filters were prehybridized for 1 h at 60°C in a solution of 50% deionized formamide, 5× SSPE, 5× Denhardt's reagent, 0.5% SDS, and 200 µg/mL denatured sheared salmon sperm DNA. Hybridization of filters was carried out with fresh hybridization buffer solution containing 2 × 106 cpm/mL of the appropriate radiolabeled probe. The hybridization reaction was carried out at 60°C for 12-18 h.
Filters were washed with three low stringency washes (1× SSPE, 0.5% SDS) at 68°C and a high stringency wash (0.1× SSPE, 0.5% SDS) at 65°C. Hybridization signals were detected and quantitated using an AMBIS 4000 Radioanalytic Imaging System (AMBIS Inc., San Diego CA). The AMBIS 4000 simultaneously images and quantitates the radioisotopic signal generated by32 P on the filters as previously described(15). Background counts above each lane were determined and subtracted from the total counts generated in each region of interest to yield a net count value. The washed filters were additionally exposed to Kodak XAR film at -70°C.
Data analysis. For quantitation of mRNA abundance, all individual tissue samples were analyzed together on a single Northern blot hybridization to control for day-to-day variations in hybridization efficiency. Northern blots were done in triplicate. Abundances of AT1 and AT2 mRNA and 18 S rRNA were expressed as total net radioactive counts after subtracting background counts for each sample. AT1 and AT2 32P net counts were normalized by the corresponding 18 S rRNA net count.
Statistical analyses of differences between hemodynamic and arterial blood values were performed using two-way ANOVA, factoring for age and treatment group. Ontogenic differences in AT1 or AT2 mRNA abundance within individual heart chambers were analyzed by one-way ANOVA. Changes in mRNA levels in response to infusion of losartan or ANG II were determined by a multiway ANOVA, factoring for twinship, treatment group, and chamber. Post hoc testing was performed by the Student-Newman-Keuls method. Statistical significance is defined as p < 0.05, and results are presented as mean ± SE.
RESULTS
Cardiac AT1 and AT2 mRNA expression in fetal and newborn sheep. Cardiac AT1 mRNA expression was unchanged in the right ventricle and left and right atria during the last trimester of gestation and during the first 8 wk of postnatal life (Figs. 1 and 2). The pattern of AT1 gene expression slightly differed in the left ventricle compared with other cardiac chambers, being lower at 7 d of life compared with those at 115 d of gestation.

Developmental changes in cardiac AT1 and AT2 gene expression in fetal and newborn sheep. Representative autoradiograms of a Northern blot of total left and right ventricle RNA hybridized with a sheep AT1 and AT2 RNA probe and a human 18 S rRNA probe.

Developmental changes in cardiac AT1 gene expression in fetal and newborn sheep. Graphs summarize results of cardiac AT1 mRNA levels (expressed as net counts of32 P and normalized for 18 S) in fetal (term 145 d of gestation) and newborn sheep: *p < 0.05 compared with 115-d fetus. n = 7-10 in each group. Values are means ± SE.
In contrast to the relatively stable levels of AT1 mRNA expression in fetal and newborn animals, the expression of cardiac AT2 receptor gene was high during the third trimester of fetal development and decreased rapidly after birth (Figs. 1 and 3). This decrease was more pronounced (p < 0.05) in the left ventricle and atrium compared with the right ventricle and atrium, respectively, at 2 d of life, but not at other postnatal ages.

Developmental changes in cardiac AT2 gene expression in fetal and newborn sheep. Graphs summarize results of cardiac AT2 mRNA levels (expressed as net counts of32 P and normalized for 18 S) in fetal (term 145 d of gestation) and newborn sheep: *p < 0.05 compared with all fetal values,†p < 0.05 compared with 135-d fetus. n = 7-10 in each group. Values are means ± SE.
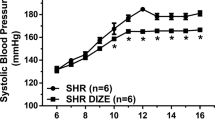
Effects of ANG II infusion on cardiac AT1 and AT2 mRNA expression in third trimester fetal sheep. Because circulating levels of ANG II transiently increase at birth, we sought to determine whether this effect contributes to the rapid down-regulation of cardiac AT2 mRNA which occurs after the transition from fetal to newborn life. Nine pairs of chronically instrumented fetal sheep (129 ± 2 d of gestation) were studied; one of each set of twins receiving a continuous i.v. infusion of ANG II (10 µg/h, 9.5 nM/h) for 24 h. The effects of ANG II infusion on arterial blood gas values, hematocrit, systemic hemodynamics PRA, and plasma ANG II concentration in this group of animals have previously been reported(16). Briefly, there were no differences in arterial blood gas values, hematocrit, or heart rate in either group before or after the 24-h infusion period. After 24 h of infusion, PRA was slightly lower in the ANG II-treated group (2.05 ± 0.67) compared with control 3.05± 1.13 ng ANG I mL-1 h-1, whereas plasma ANG II was increased (210 ± 21 versus 95 ± 9 pg/mL). Mean arterial blood pressure was also significantly higher in the ANG II group 24 h after infusion compared with the control animals (76 ± 5versus 54 ± 3 mm Hg, respectively).
The effects of ANG II infusion on cardiac AT1 and AT2 mRNA levels are presented in Figures 4 and 5. Northern blot hybridization demonstrated no significant changes in left or right ventricle AT1 or AT2 mRNA levels in ANG II-treated fetuses when compared with twin matched control fetuses. To determine whether tissue-specific effects of ANG II infusion on AT1 receptor mRNA expression existed, we also determined changes in AT1 mRNA levels in fetal liver and adrenal glands. In contrast to the lack of change in cardiac tissue, AT1 mRNA levels significantly decreased (p < 0.05) in adrenal glands (2350 ± 340 versus 1558 ± 384 net counts) with ANG II infusion but increased (p < 0.05) in liver (1199 ± 209 versus 2018 ± 322 net counts). A 27 ± 11% decrease (p < 0.05) in fetal renal cortical AT1 mRNA in response to ANG II infusion in this group of animals has previously been reported(16).

ANG II regulation of AT1 and AT2 gene expression. Autoradiograms of Northern blots of total cardiac ventricle mRNA hybridized with a sheep AT1 and AT2 RNA probe and a human 18 S rRNA probe. C, control;AII, ANG II infusion.

ANG II regulation of AT1 and AT2 gene expression. Summary data for left and right ventricle AT1 and AT2 mRNA levels (expressed as net counts of32 P and normalized for 18 S) in control and ANG II-infused fetal sheep. Nine pairs of twin fetuses were studied. Values are means ± SE.
Effects of losartan on cardiac AT1 and AT2 mRNA expression in third trimester fetal sheep. These studies were designed to determine whether blockade of AT1 receptors with losartan, and the accompanying decrease in arterial pressure, would alter the expression of cardiac AT1 and AT2 mRNA. The effects of losartan infusion of fetal arterial blood values, mean arterial blood pressure, and heart rate are presented in Table 1. No significant changes in arterial pH, PCO2, PO2, hematocrit, or heart rate were observed in either control or losartan-treated fetuses in any age group. Significant decreases in fetal arterial pressure in response to losartan were observed in animal at 110 d (40 ± 1 versus 33 ± 1 mm Hg) and 135 d of gestation (43 ± 1 versus 32 ± 1 mm Hg) but not at 95 d of gestation.
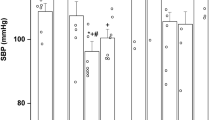
After the 48-h infusion of losartan, statistically significant increases in right atrium AT1 mRNA levels were seen in 95- and 135-d gestation fetuses (Fig. 6). However, except for an increase in left ventricular AT1 mRNA at 110 d (178 ± 21% of control,p < 0.05), infusion of losartan had no significant effect on AT1 mRNA levels in other cardiac chambers at any gestational age. Additionally, cardiac AT2 receptor gene expression was unaltered by AT1 receptor blockade at all three gestational ages determined during the third trimester of gestation (Fig. 6).

Effect of AT1 receptor blockade on cardiac AT1 (top panels) and AT2(bottom panels) mRNA levels during fetal development. Results represent changes in left atrium (LA), left ventricle(LV), right atrium (RA), and right ventricle(RV) AT1 mRNA levels in response to losartan (20 µg kg-1 m-1 i.v. for 48 h) expressed as a percentage of control ± SE. Studies were done in twin fetuses at 95, 110, and 135 d of gestation (term 145 d), n = 7-9 at each gestational age.*p < 0.05 compared with the control value.
DISCUSSION
The results of this study demonstrate that cardiac AT1 and AT2 mRNA expression are differentially regulated during the last trimester of gestation in fetal sheep and postnatal development. AT1 mRNA levels remain relatively constant in all four cardiac chambers during this period of maturation, whereas AT2 mRNA levels rapidly decrease during the 1st wk of postnatal life. In contrast to other fetal tissues, cardiac AT1 and AT2 mRNA expression is not altered by increased circulating ANG II levels. Furthermore, we observed that during the third trimester of gestation, blockade of AT1 receptors has no significant effect on cardiac AT2 mRNA abundance and no consistent effect on AT1 mRNA levels (increased expression only in right atrium at 95 and 135 d and in left ventricle at 110 d). Finally, based on changes in blood pressure and PRA, the results suggest that AT1 receptors become functional during the mid third trimester in fetal sheep.
Two classes of ANG II receptors exist in the developing myocardium, as well as in the mature heart, and are present on cardiac myocytes and fibroblasts(17). The inotropic, chronotropic, and growth-promoting effects of ANG II on the heart appear to be mediated by the AT1 receptor, whereas the physiologic role of the AT2 receptor remains speculative(7,18). The AT2 receptor is expressed early during embryonic life in the rat(4,19), predominates in the fetal mesenchyme(4), and shows a marked decrease in expression during postnatal maturation. The predominance of the AT2 receptor during fetal life has led investigators to postulate that these receptors may be involved in protein synthesis and growth(4,20). Original reports of mice with targeted disruption of the AT2 gene demonstrate that, in this species, the AT2 receptor is not required for normal embryonic and postnatal development(21,22). However, it has recently been reported that AT2-deficient mice express diverse urinary tract anomalies(23), suggesting a role for AT2 receptor activation in normal ureter and kidney development.
Studies in a number of other species have shown that cardiac angiotensin receptor expression is developmentally regulated. ANG II binding sites are present in the rat heart by embryonic d 14, increase during gestation and immediately after birth, then decrease during with postnatal maturation(24–27). Ventricular ANG II receptor density is also significantly higher in human neonatal heart compared with adult heart(27). These postnatal changes in ANG II binding sites result from down-regulation of both AT1 and AT2 receptors(25,26). In vitro studies have shown that AT1 mRNA and receptor binding levels are similar in embryonic d 19 and 1-d neonatal cultured rat cardiac fibroblasts and myocytes, whereas AT2 receptor expression significantly decreased by 1 d of life(17). The results of the present study, which demonstrate that cardiac AT2 mRNA levels are highest during fetal life and decrease after birth, provide additional evidence for developmental regulation of myocardial ANG II receptors. Interestingly, cardiac AT1A receptor mRNA levels are increased over 3-fold during the 1st wk of life in the rat compared with levels in animals 4 wk and older(25). In contrast, we found that myocardial AT1 mRNA levels are relatively consistent late in fetal development and during the first 2 mo of life. Reasons for the differences are likely related to differences in species and study design.
The distribution of ANG II receptors within the heart has also been investigated, although discrepant results have been obtained. Hunt et al.(24) found atrial ANG II receptors levels to be twice ventricular levels during the perinatal period but equalized by adulthood. Other investigators have found in adult guinea pigs and rabbits that ANG II binding remains higher in the atria than in ventricles(28,29). Receptor binding studies have indicated equivalent AT1 and AT2 receptor populations on ventricular membranes. However, in rats, in situ hybridization studies fail to detect AT2 mRNA in fetal, neonatal, or adult cardiac muscle, although expression in subendocardial atrial tissue is present during the early postnatal period(20). In the rat fetus, AT1 mRNA is diffusely localized, the highest signal being visible in the atrioventricular septum, interventricular septum, and bundle of His(30). It should be noted that, in the present study, no attempt was made to quantify expression of AT1 or AT2 mRNA for comparison among the four cardiac chambers. However, in a previous study we found in 135-d gestation fetal sheep that AT1 mRNA levels were similar among left and right atria and ventricles(31). Although it is possible that uneven distribution of ANG II receptors within the heart may alter measurements within discrete regions, it is unlikely that this spatial distribution affected our results. We obtained large pieces of tissue from ventricular free wall and both atria, excluding the septa. The samples were then homogenized separately in the process of RNA extraction. In addition, the developmental pattern of AT1 and AT2 mRNA expression were relatively consistent among the different cardiac chambers.
In contrast to the AT1 receptor, little is known about the structure, function, or regulation of the AT2 receptor. Detection of cardiac AT2 mRNA has been difficult, although expression of myocardial AT1 and AT2 receptors is relatively similar(17,24). Although other investigators have failed to detect myocardial AT2 transcripts by Northern blot or in situ hybridization(20), or have needed to use reverse transcription-polymerase chain reaction(32), we were able to detect AT2 mRNA by Northern blot analysis using total RNA prepared from ovine hearts. We examined AT2 mRNA expression early in development, when gene expression is highest, as opposed to most other investigators who have examined gene regulation in adult hearts. It is possible that, in adult ovine heart, AT2 mRNA transcripts may not be detectable by Northern analysis. Using a sheep AT2 cDNA probe labeled with 32P, we detected two transcripts for AT2, similar to that reported in a previous study(33). Although we are not aware of more than one transcript being present in other species, this certainly does not exclude the presence in sheep. Within the species, there may be post-transcriptional modification of the mRNA, alternative splicing, or non-AT2-related mRNAs. Further investigation and characterization of these two transcripts is needed to clarify this issue. Regardless, we have seen the multiple AT2 transcript repeatedly, and the ratio of the two band has been consistent among conditions.
In addition to characterizing developmental regulation of cardiac AT1 and AT2 mRNA expression, we sought to determine the regulatory effects of ANG II receptor stimulation and AT1 receptor blockade on levels of AT1 and AT2 mRNA. Our results demonstrate that an increase in circulating ANG II for 24 h did not alter AT1 or AT2 mRNA levels in fetal sheep heart. In vitro, ANG II up-regulates AT1 receptor gene expression in cultured neonatal rat cardiomyocytes and fibroblasts(34). However, in adult rats, physiologic states that increase endogenous ANG II levels have no consistent effect of AT1 receptor gene expression(35). It is possible that longer exposure to elevated circulating levels of ANG II may have altered AT1 and AT2 gene expression. In the present study, there appeared to be a downward trend in both ventricular AT1 and AT2 receptor mRNA levels in response to ANG II (Figs. 4 and 5), although the magnitude of these changes did not reach statistical significance.
The effects of ANG II on AT1 and AT2 mRNA expression appear to be tissue-specific, because, in the present study, infusion of ANG II increased AT1 mRNA levels in fetal liver, but decreased expression in adrenal and kidney within these same animals(16). These results differ from previous studies in adult rats that found ANG II infusion increased expression of AT1 mRNA in adrenal gland but not in the kidney(36); unfortunately, changes in the heart were not explored. It should also be noted that we investigated the effects of a single infusion dose of ANG II in one age group. Different results may have been obtained if different age groups of doses of ANG II were studied. The precise mechanism(s) for ANG II-induced regulation of AT1 receptors and mRNA are currently unknown. It has been suggested that activation of protein kinases C and A may be involved in up-regulation of AT1 receptor mRNA and binding sites(37,38). On the other hand, ANG II-induced down-regulation of AT1 mRNA in cultured mesangial cells appears independent of protein kinase A and protein kinase C(39). Fetal sheep kidney AT2 mRNA expression is also down-regulated by ANG II infusion(16), whereas no effects were detected on cardiac AT2 mRNA levels. Similar tissue-specific regulation of several components of the RAS has previously been reported in a number of different species(31,36,40).
Studies examining blockade of the renin-angiotensin system on cardiac AT1 receptor expression have been inconsistent. In adult rat, specific AT1 receptor blockade either up-regulates(41), down-regulates(42), or has no effect(35) on cardiac AT1 mRNA levels. In the fetal lamb, losartan had little effect on ventricular AT1 or AT2 mRNA levels (except left ventricle at 110-d gestation). This finding is in accord with our previous finding in pulmonary artery-banded fetal lambs that losartan treatment for 1 wk does not alter expression of left or right ventricle AT1 receptor protein or mRNA or AT2 mRNA(33). Because losartan produced a significant decrease in fetal arterial blood pressure, one could argue that the accompanying hemodynamic effects may have masked gene regulating effects of AT1 receptor blockade. Surprisingly, right atrial AT1 mRNA levels were elevated in losartan-treated animals in two of three age groups studied. Reasons for this chamber-specific effect are unknown but suggest that AT1 receptors within the heart may be differentially regulated.
In summary, cardiac AT2 gene expression is high during the third trimester of gestation and decreases rapidly after birth, whereas expression of AT1 receptor mRNA remains relatively constant during this time. Activation of AT1 and AT2 receptors late during fetal development fails to alter expression of these receptors. Inhibition of AT1 receptors produced a significant decrease in blood pressure in 110- and 135-d fetuses, but not 95-d, demonstrating that the RAS contributes to cardiovascular regulation during the mid third trimester in fetal lambs. Furthermore, inhibition of AT1 receptors has little effect on ventricular AT1 or AT2 mRNA levels during fetal development, suggesting that developmental changes in AT1 receptor activity are not involved in the regulation of AT2 receptor expression during cardiac development.
Abbreviations
- ANG II:
-
angiotensin II
- AT1:
-
angiotensin type 1 receptor
- AT2:
-
angiotensin type 2 receptor
- PRA:
-
plasma renin activity
- RAS:
-
renin-angiotensin system
References
Davidson D 1987 Circulating vasoactive substances and hemodynamic adjustments at birth in lambs. J Appl Physiol 63: 676–684.
Guillery EN, Robillard JE 1993 The renin-angiotensin system and blood pressure regulation during infancy and childhood. In: Rocchini AP (ed), The Pediatric Clinics of North America: Childhood Hypertension. WB Saunders, Philadelphia, 61–77.
Segar JL, Mazursky JE, Robillard JE 1994 Changes in ovine renal sympathetic nerve activity and baroreflex function at birth. Am J Physiol 267: H1824 H1832
Grady EF, Sechi LA, Griffin CA, Schambelan M, Kalinyak JE 1991 Expression of AT2 receptors in the developing rat fetus. J Clin Invest 88: 921–933.
Beinlich CJ, White GJ, Baker KM, Morgan HE 1991 Angiotensin II and left ventricular growth in newborn pig heart. J Mol Cell Cardiol 23: 1031–1038.
Aceto JF, Baker KM 1990 [Sar1]angiotensin II receptor-mediated stimulation of protein synthesis in chick heart cells. Am J Physiol 258:H806–H813.
Sadoshima JI, Izumo S 1993 Molecular characterization of angiotensin II-induced hypertrophy of cardiac myocytes and hyperplasia of cardiac fibroblasts. Circ Res 73: 413–423.
Smith RD, Chiu AT, Wong PC, Herblin WF, Timmermans PBMWM 1992 Pharmacology of nonpeptide angiotensin II receptor antagonists. Annu Rev Pharmacol Toxicol 32: 135–165.
Chiu AT, Herblin WF, Ardecky RJ, McCall DE, Carini DJ, Duncia JV, Pease LJ, Wexler RR, Wong PC, Johnson AL, Timmermans PBMWM 1989 Identification of angiotensin II receptor subtypes. Biochem Biophys Res Commun 165: 196–203.
Tsutsumi K, Saavedra JM 1991 Characterization and development of angiotensin II receptor subtypes (AT1 and AT2) in rat brain. Am J Physiol 261:R209–R216.
Tsutsumi K, Saavedra JM 1991 Quantitative autoradiography reveals different angiotensin II receptor subtypes in selected rat brain nuclei. J Neurochem 56: 348–351.
Jennings JJ, Crowley JP 1972 The influence of mating management on fertility in ewes following progesterone-PMS treatment. Vet Rec 90: 495–498.
Robillard JE, Weitzman RE 1980 Developmental aspects of the fetal renal response to exogenous arginine vasopressin. Am J Physiol 238:F407–F414.
Mellor DJ 1969 Vascular anastomosis and fusion of foetal membranes in multiple pregnancy in the sheep. Res Vet Sci 10: 361–369.
Robillard JE, Schutte BC, Page WV, Fedderson JA, Porter CC, Segar JL 1994 Ontogenic changes and regulation of renal angiotensin II type 1 (AT1) receptor gene expression during fetal and newborn life. Pediatr Res 36: 755–762.
Robillard JE, Page WV, Mathews MS, Schutte BC, Nuyt A-M, Segar JL 1995 Differential gene expression and regulation of renal angiotensin II receptor subtypes (AT1 and AT2) during fetal life in sheep. Pediatr Res 38: 896–904.
Matsubara H, Kanasaki M, Murasawa S, Tsukaguchi Y, Nio Y, Inada M 1994 Differential gene expression and regulation of angiotensin II receptor subtypes in rat cardiac fibroblasts and cardiomyocytes in culture. J Clin Invest 93: 1592–1601.
Schunkert H, Sadoshima J-I, Cornelius T, Kagaya Y, Weinberg EO, Izumo S, Riegger G, Lorell BH 1995 Angiotensin II-induced growth responses in isolated adult rat hearts. Evidence for load-independent induction of cardiac protein synthesis by angiotensin II. Circ Res 76: 489–497.
Tsutsumi K, Saavedra JM 1992 Heterogeneity of angiotensin II AT2 receptors in the rat brain. Mol Pharmacol 41: 290–297.
Shanmugam S, Corvol P, Gasc J-M 1996 Angiotensin II type 2 receptor mRNA expression in the developing cardiopulmonary system of the rat. Hypertension 28: 91–97.
Ichiki T, Labosky PA, Shlota C, Okuyama S, Imagawa Y, Fogo A, Niimura F, Ichikawa I, Hogan BLM, Inagami T 1995 Effects on blood pressure and exploratory behaviour of mice lacking angiotensin II type-2 receptor. Nature 377: 748–750.
Hein L, Barsh GS, Pratt RE, Dzau VJ, Kobilka BK 1995 Behavioural and cardiovascular effects of disrupting the angiotensin II type-2 receptor gene in mice. Nature 377: 744–747.
Nishimura H, Yerkes E, Schulman M, Fogo A, Kon V, Brigid MLH, Brock JW, Tadashi I, Iekuni I 1997 The angiotensin type 2 (AT2) receptor null mutant mice: a model of the diverse spectrum of congenital urinary tract anomalies in humans. Pediatr Res 41: 282A
Hunt RA, Ciuffo GM, Saavedra JM, Tucker DC 1995 Quantification and localisation of angiotensin II receptors and angiotensin converting enzyme in the developing rat heart. Cardiovac Res 29: 834–840.
Suzuki J, Matsubara H, Urakami M, Inada M 1993 Rat angiotensin II (type 1A) receptor mRNA regulation and subtype expression in myocardial growth and hypertrophy. Circ Res 73: 439–447.
Sechi LA, Chandi AG, Grady EF, Kalinyak JE, Schambelan M 1992 Characterization of angiotensin II receptor subtypes in rat heart. Circ Res 71: 1482–1489.
Urata H, Healy B, Stewart RW, Bumpus FM, Husain A 1989 Angiotensin II receptors in normal and failing human hearts. J Clin Endocrinol Metab 69: 54–66.
Baker KM, Campanile CP, Trachte GJ, Peach MJ 1984 Identification and characterization of the rabbit angiotensin II myocardial receptor. Circ Res 54: 286–293.
Baker KM, Singer HA 1988 Identification and characterization of guinea pig angiotensin II ventricular and atrial receptors: coupling to inositol phosphate production. Circ Res 62: 896–904.
Shanmugam S, Monnot C, Corvol P, Gasc J 1994 Distribution of type 1 angiotensin II receptor subtype messenger RNAs in the rat fetus. Hypertension 23: 137–141.
Segar JL, Bedell K, Page WV, Mazursky JE, Nuyt A-M, Robillard JE 1995 Effect of cortisol on gene expression of the renin-angiotensin system in fetal sheep. Pediatr Res 37: 741–746.
Nio Y, Matsubara H, Murasawa S, Kanasaki M, Inada M 1995 Regulation of gene transcription of angiotensin II receptor subtypes in myocardial infarction. J Clin Invest 95: 46–54.
Segar JL, Scholz TD, Bedell KA, Smith OM, Huss DJ, Guillery EN 1997 Angiotensin AT1 receptor blockade fails to attenuate pressure-overload cardiac hypertrophy in fetal sheep. Am J Physiol 273:R1501–R1508.
Everett AD, Heller F, Fisher A 1996 AT1 receptor gene regulation in cardiac myocytes and fibroblasts. J Mol Cell Cardiol 28: 1727–1736.
Bruna RD, Ries S, Himmelstoss C, Kurtz A 1995 Expression of cardiac angiotensin II AT1 receptor genes in rat hearts is regulated by steroids but not by angiotensin II. J Hypertens 13: 763–769.
Iwai N, Inagami T 1992 Regulation of the expression of the rat angiotensin II receptor mRNA. Biochem Biophys Res Commun 182: 1094–1099.
Lu D, Sumners C, Raizada MK 1994 Regulation of angiotensin II type 1 receptor mRNA in neuronal cultures of normotensive and spontaneously hypertensive rat brains by phorbol esters and forskolin. Biochem Biophys Res Commun 62: 2079–2084.
Smith RD 1994 Protein kinase C rapidly up-regulates the number of AT1 angiotensin receptors on cultured rat intestinal epithelial (RIE-1) cells. FEBS Lett 348: 51–54.
Makita N, Iwai N, Inagami T, Badr KF 1992 Two distinct pathways in the down-regulation of type-1 angiotensin II receptor gene in rat glomerular mesangial cells. Biochem Biophys Res Commun 185: 142–146.
Dzau VJ, Brody T, Ellison KE, Pratt RE, Ingelfinger JR 1987 Tissue-specific regulation of renin expression in the mouse. Hypertension 9: III36–III41
Everett AD, Tufro-McReddie A, Fisher A, Gomez RA 1994 Angiotensin receptor regulates cardiac hypertrophy and transforming growth factor-β1 expression. Hypertension 23: 587–592.
Kitami Y, Okura T, Marumoto K, Wakamiya R, Hiwada K 1992 Differential gene expression and regulation of type-1 angiotensin II receptor subtypes in the rat. Biochem Biophys Res Commun 188: 446–452.
Acknowledgements
The authors gratefully acknowledge the assistance of Mark A. Hart in the preparation of this manuscript.
Author information
Authors and Affiliations
Additional information
Supported by a Research Training Grant in Cardiology HL-07413 (M.E.S), and by Clinical Investigator Development Award HL-02865 and a Grant-in-Aid from the American Heart Association (J.L.S.).
Rights and permissions
About this article
Cite this article
Samyn, M., Petershack, J., Bedell, K. et al. Ontogeny and Regulation of Cardiac Angiotensin Types 1 and 2 Receptors during Fetal Life in Sheep. Pediatr Res 44, 323–329 (1998). https://doi.org/10.1203/00006450-199809000-00010
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-199809000-00010
This article is cited by
-
Cellular and RAS Changes in the Hearts of Young Obese Rats
Pediatric Cardiology (2011)
-
Maturation of the angiotensin II cardiovascular response in the embryonic White Leghorn chicken (Gallus gallus)
Journal of Comparative Physiology B (2010)
-
Maternal nutrient restriction and the fetal left ventricle: Decreased angiotensin receptor expression
Reproductive Biology and Endocrinology (2005)
