
Abstract
The decreased contractility of gastric antral smooth muscle in the neonate has been attributed to reduced levels of activator calcium. It is generally accepted that calcium-dependent myosin light chain phosphorylation (MLCP) is the key step in the initiation of force development in smooth muscle. In this study, we investigated the relationship between MLCP and force development in gastric antral smooth muscle from neonatal (4-6 d old) and adult rabbits. We tested the hypothesis that the reduced force development of circular smooth muscle from the neonate would be accompanied by decreased levels of MLCP, as compared with data from adult animals. Full thickness muscle strips oriented parallel to the circular muscle layer were examined for their contractile response to acetylcholine (ACh) (10-8 M to 10-3 M) or 10-4 M ACh only. In the latter study, tissues were rapidly frozen in a dry ice-acetone slurry for subsequent MLCP determination. MLCP was determined at times corresponding to 5, 10, 15, 30, and 60 s of stimulation. For each age group, maximal active force developed at an ACh concentration of 10-4 M and was significantly greater in tissues from adults (1.86 ± 0.24 N/m2, adult; 0.95 ± 0.05 N/m2, neonate; p < 0.05). In contrast, no significant differences were observed with respect to basal or agonist-stimulated levels of MLCP. The data suggest that factors other than levels of MLCP contribute to the reduced force-generating capacity of antral smooth muscle from the neonate.
Similar content being viewed by others

Main
Delayed gastric emptying and abnormal antral motility patterns are common in preterm and newborn infants(1–4). Although the factors involved in the postnatal maturation of gastric motility remain to be fully elucidated, we and others have reported that antral smooth muscle from newborn animals differs from that of the adult with respect to maximal active force development(5–9), sensitivity to agonist stimulation(8), involvement/mobilization of extracellular and intracellular calcium stores(6–9), and the characteristics of cell membrane calcium channels(7). These findings have led to the hypothesis that the postnatal maturation of gastric motility involves, at least in part, an overall increase in the force-generating capacity(contractility) of antral smooth muscle.
An essential first step in smooth muscle contraction is the calcium-dependent phosphorylation of the 20-kD regulatory myosin light chain by the enzyme, myosin light chain kinase (for reviews, seeRefs. 10 and 11). MLCP is brought about by the interaction of calcium, the calcium-activated protein calmodulin, and a calcium-calmodulin-dependent protein responsible for MLCP called myosin light chain kinase. Phosphorylation of the 20-kD myosin light chain on each myosin head brings about a conformational change which results in actin activation of the myosin-Mg2+-ATPase, and subsequent crossbridge cycling. In the muscle cell, crossbridge cycling produces a mechanical output, which is expressed either as force development or shortening, depending upon the physical constraints on the muscle.
It is well documented that agonist-mediated increases in cytosolic calcium lead to rapid increases in MLCP and subsequent force development in vascular and tracheal smooth muscle(10, 11). Although it is also recognized that increases in cytoplasmic calcium levels initiate contraction in gastrointestinal smooth muscle, the relationship between MLCP and force development is less well studied. However, the data available suggest that MLCP is essential to the contractile event and that a correlation exists between levels of MLCP and force output by the muscle(12–17). The purpose of this study was to evaluate the relationship between active force development and MLCP in circular smooth muscle from the gastric antrum of adult and neonatal rabbits. Specifically, we tested the hypothesis that the reduced stress-generating capacity of antral smooth muscle from neonatal rabbits can be accounted for by reduced levels of MLCP.
METHODS
Tissue preparation. Antral muscle strips were obtained from neonatal (4-6 d old) and adult (female, 5-6 kg) New Zealand White rabbits as described previously(5). The stomachs were removed through a midline incision, opened along the greater curvature, and rinsed free of residual food contents using room temperature (23°C), oxygenated(100% O2) HEPES buffer solution of the following composition (in mM): 137.5 NaCl, 5.0 KCl, 1.0 MgCl2, 2.5 CaCl2, and 10.0 glucose, at pH 7.4. The antrum was cut free from the remainder of the stomach and pinned mucosal side up in a dissecting dish filled with fresh room temperature HEPES buffer solution. With the aid of a stereomicroscope, full thickness muscle strips (1 mm × 5-10 mm) were prepared by careful separation of the mucosa from the underlying muscle. The microscope permitted clear visualization of the muscle bundles and ensured that the long axis of the muscle strip was parallel to the circular muscle layer.
The muscle strips were mounted in individual 10-mL tissue baths filled with warmed (37°C), aerated HEPES buffer solution. One end was attached via an inelastic wire to an isometric force transducer (Grass, FT-03C), and the other end attached to a glass rod that could be raised or lowered by adjustment of fine micrometer. After a 1-h equilibration period, the muscle strips were stretched in 0.5-mm increments and stimulated with ACh (10-4 M) until the length at which maximal active stress develops was achieved(5). A permanent record of the force developed by each muscle strip was obtained using a multichannel recorder.
Response to ACh stimulation. Each muscle strip was examined for its contractile response to ACh (10-8 M to 10-3 M). ACh was added to the baths in microliter amounts to achieve the final concentrations reported. All dose-response curves were constructed with ACh being given in random doses; no cumulative dose-response curves were obtained. The dose-response data for tissues from neonates and adults were analyzed for differences in active stress (N/m2) and ED50 (defined as the dose of ACh which elicits a one-half maximal response). Also, in some instances, chart recorder paper speed was increased to permit assessment of the relationship between time after agonist administration and the development of the contractile response. Specifically, we determined the time between addition of the agonist and the beginning of force development (lag time), and the time to one-half (T50) and maximal(T100) force development.
Myosin light chain phosphorylation determination. MLCP levels in antral smooth muscle from neonatal and adult rabbits were measured under basal (unstimulated) conditions and after ACh (10-4 M) stimulation using techniques described previously(12–14). Stimulated tissues were studied at 5, 10, 15, 30 and 60 s of stimulation. At the appropriate time, each muscle strip was rapidly frozen in a dry ice-acetone slurry and then allowed to thaw in this solution for 60 min at 22°C. This procedure denatures endogenous phosphatases and kinases thereby preserving the levels of MLCP that existed at the time of freezing. The muscle strips were homogenized at 4°C in 1% SDS, 10% (wt/vol) glycerol, and 20 mM DTT. The phosphorylated and unphosphorylated myosin light chains were resolved using a two-dimensional gel electrophoretic system involving isoelectric focusing in the first dimension and SDS-PAGE electrophoresis in the second dimension(18). MLCP was quantitated by densitometric scanning of Coomassie Blue-stained gels using a Helena Labs Quick-scan Jr. densitometer (Beaumont, TX). The amount of phosphorylated myosin light chain was calculated by dividing the area of the phosphorylated peak by the total area of the phosphorylated and unphosphorylated peaks and expressing the results as a percent myosin light chain phosphorylation. The acidic satellite forms of the phosphorylated light chains were rarely observed and therefore excluded from the analysis.
Data analysis. Upon completion of each experiment, the length and weight of each muscle strip was determined and used to calculate tissue cross-sectional area as described previously(5). All contractions were normalized for tissue cross-sectional area, and the data were reported as stress in N/m2. Values of ED50 were obtained from a probit analysis of the dose-response curves(19). Statistical analyses utilized analysis of variance and the Student'st test for unpaired observations to compare data from neonatal and adult animals. Differences were considered statistically significant whenp was <0.05. All data are presented as mean ± SEM.
RESULTS
Response to ACh stimulation. ACh stimulation of antral smooth muscle from neonates and adults produced a rapid increase in tension (phasic response) followed by reduced but sustained contraction (tonic response). No statistically significant differences existed between tissues from neonatal and adult animals with respect to lag time (3.4 ± 0.4 s, neonate; 3.2± 0.3 s, adult), the time to one-half peak response (3.9 ± 0.5 s, neonate; 4.5 ± 0.6 s, adult) or the time to peak response (11.2± 0.7 s, neonate; 12.4 ± 1.0 s, adult).
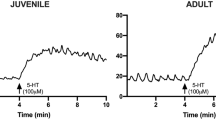
Antral smooth muscle from neonates and adult rabbits responded to increasing concentrations of ACh with increased force (Fig. 1). At each concentration above 10-6 M ACh, the force developed by muscle strips from adult rabbits was significantly greater than the force developed by tissues from neonates. For each age group, maximal active stress developed at an ACh concentration of 10-4 M and was 1.86 ± 0.24 N/m2 for the adult and 0.95 ± 0.05 N/m2 for the neonate(p < 0.05).

ACh dose-response curves of gastric antral smooth muscle from neonatal and adult rabbits. Each data point represents the mean± SE of at least 12 experiments.
Normalization of the contractile data from neonates and adult animals to 100% of their maximal responses produced virtually identical dose-response relationships (Fig. 2). No statistical difference was found between the two age groups with respect to the ED50 of the dose-response curves; 3.9 ± 1.2 × 10-6 M for the adult and 5.7 ± 1.4 × 10-6 M for the neonate (p > 0.05).

Normalization of data shown in Figure 1 to 100% of respective maximal responses. Each data point represents the mean ± SE of at least 12 experiments.
MLCP. Figure 3 summarizes the MLCP data obtained from antral smooth muscle of neonatal and adult rabbits. Basal levels of MLCP were similar in neonates and adult animals, 7.4 ± 1.4% and 8.2± 0.9%, respectively (p > 0.05). When stimulated with 10-4 M ACh, tissues from both the neonate and the adult evidenced identical time-dependent changes in MLCP. In antral smooth muscle from the neonate and the adult, MLCP reached peak levels at 10 s of stimulation(neonate, 48.7 ± 1.9%; adult, 44.6 ± 2.3%; p 0.05) and returned to near basal levels by 60 s of stimulation (neonate 13.3 ± 2.4%; adult, 12.0 ± 2.3%).

Levels of MLCP in circular smooth muscle from the gastric antrum of neonatal and adult rabbits under basal conditions and after ACh (10-4 M) stimulation. Each data point represents the mean ± SE of at least six experiments.
DISCUSSION
Smooth muscle contains an endogenous enzyme, myosin light chain kinase, capable of phosphorylating the 20-kD light chains of myosin. It is now well documented that contraction of smooth muscle is initiated by phosphorylation of the 20-kD light chain of myosin(10, 11). Phosphorylation of myosin is the direct result of an increase in the cytoplasmic concentration of free calcium which occurs as a result of agonist-mediated calcium influx and/or calcium mobilization from intracellular stores. Calcium-dependent MLCP results in actin activation of myosin ATPase activity, the development of force, and muscle shortening. Evidence exists in support of a close correlation between the levels of activator calcium, maximal active stress, and peak levels of MLCP for a number of smooth muscles(10, 20, 21).
The primary purpose of this study was to test the hypothesis that the decreased force-generating capacity of antral smooth muscle from neonatal rabbits is due in part to reduced levels of MLCP. This hypothesis is based on two assumptions. First, a close correlation exists between the amount of calcium available for activation of the contractile process and peak levels of MLCP. Second, antral smooth muscle from neonatal animals develops less active stress than tissues from adult animals in part as the result of reduced amounts of calcium available for activation of the contractile machinery. The results of the present study indicate that MLCP also is a regulatory step in antral smooth muscle contraction, in as much as unstimulated muscle had low levels of MLCP, and ACh stimulation was followed by rapid increases in MLCP. The observed basal and peak levels of MLCP are similar to values reported for gallbladder(12–14), uterine(22–24), tracheal(10, 25), and vascular(11, 21, 26) smooth muscle. However, the findings of the present study do not support our hypothesis. Although we did observe a significant difference in the maximal active stress developed by antral smooth muscle from neonates and adults, no significant differences were observed with respect to the time dependency or the levels of MLCP attained in response to ACh stimulation. The data suggest that at equivalent levels of MLCP tissues from neonatal animals develop less active stress than tissues from adult animals. Thus, as compared to the adult, gastric antral smooth muscle from the neonate is characterized by a decreased stress/MLCP ratio.
To date, little information is available concerning how alterations in the physiologic status of a smooth muscle affects the stress/MLCP relationship of the muscle. Xie et al.(15) compared this relationship in colonic smooth muscle from healthy rabbits and rabbits with documented experimental colitis and observed that the decreased stress generating capacity of colonic smooth muscle from colitic rabbits was associated with decreased levels of MLCP, with no change in the stress to MLCP ratio. They explained their results by assuming that the inflammatory process reduced the amount of calcium available for activation of the contractile process. Recently, Word et al.(24) demonstrated that the increased stress-generating capacity of myometrial smooth muscle from pregnant women occurred despite reduced levels of MLCP. They observed that the stress to MLCP ratio of uterine smooth muscle was increased more than two times in tissues from pregnant women. The findings of Xie et al.(15) and Word et al.(24), as well as the results of this study suggest that factors in addition to MLCP levels must be accounted for when considering the force-generating capacity of a smooth muscle.
It is possible that differences in tissue thickness between the neonate and the adult may have affected our data and compromised our ability to determine differences in MLCP levels between the age groups. Kamm and Murphy(27) have reported that peak MLCP levels depend upon the rate of tissue activation and, as such, are affected by the thickness of the muscle strip preparation. In their study, significant increases in the thickness of an in vitro hog carotid artery preparation decreased peak MLCP values, and prolonged the lag period and the times to one-half and maximal contraction. In our study, we found that the lag time and the times to one-half maximal and maximal contraction were not different between the age groups, suggesting similar rates of tissue activation. Thus, we do not believe that our data were influenced by differences in muscle strip thickness between the neonate and the adult.
In summary, despite comparable levels of MLCP upon Ach stimulation, circular smooth muscle from the gastric antrum of neonatal rabbits developed less active stress than tissues from adult animals. Although we did not examine the factors responsible for the decreased stress to MLCP ratio in tissues from neonatal rabbits, we recently reported that the postnatal increase in antral smooth muscle contraction in the rabbit is paralleled by changes in myosin heavy chain isoform distribution(28). Ayas et al.(29) reported similar data for the fundus and antrum of the cat. They also reported that the postnatal period is associated with an increase in the cellular concentration of actin and myosin. The latter could account for increased stress development despite similar levels of MLCP. The data presently available suggest that the postnatal maturation of force in smooth muscle is a complex dynamic involving changes in contractile protein content and isoform distribution, as well as possibly changes in other cytostructural and regulatory proteins.
Abbreviations
- ACh:
-
acetylcholine
- HEPES:
-
N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid
- MLCP:
-
myosin light chain phosphorylation
References
Ittmann PI, Amarnath R, Berseth CL 1992 Maturation of antral duodenal motor activity in preterm and term infants. Dig Dis Sci 37: 14–19
Berseth CL, Ittmann PI 1992 Antral and duodenal motor responses to duodenal feeding in preterm and term infants. J Pediatr Gastroenterol Nutr 14: 182–186
Cavell B 1979 Gastric emptying in preterm infants. Acta Paediatr Scand 68: 725–730
Seigel M 1983 Gastric emptying time in premature and compromised infants. J Pediat Gastroenterol Nutr 2( suppl 1): s136–s140
Zitterman J, Ryan JP 1990 Development of gastric antral smooth muscle contractility in newborn rabbits. Am J Physiol 258:G571–G575
Paul DA, Ierardi JA, Parkman HP, Ryan JP 1994 Developmental changes in gastric fundus smooth muscle contractility and involvement of extracellular calcium in fetal and adult guinea pigs. Pediatr Res 36: 642–646
Hyman PE, Martin MG, Tomomasa T, Jing J, Snape WJ Jr 1989 Development of calcium channels in gastric smooth muscle. Pediatr Res 25: 600–604
Tomomasa T, Yagi H, Kimura S, Snape WJ Jr, Hyman PE 1989 Developmental changes in agonist-mediated gastric smooth muscle contraction in the rabbit. Pediatr Res 26: 459–461
Hillemeier AC, Bitar KN, Biancani P 1991 Developmental characteristics of the kitten antrum. Gastroenterology 101: 339–343
De Lanerolle P, Paul RJ 1991 Myosin phosphorylation/dephosphorylation and the regulation of airway muscle contractility. Am J Physiol 261:L1–L14
Murphy RA 1994 What is special about smooth muscle? The significance of covalent crossbridge regulation. FASEB J 8: 311–318
Washabau RJ, Wang MB, Dorst C, Ryan JP 1994 Role of myosin light chain phosphorylation in guinea pig gallbladder smooth muscle contraction. Am J Physiol 266:G469–G474
Washabau RJ, Wang MB, Ryan JP 1991 Myosin light chain phosphorylation and contraction of guinea pig gallbladder smooth muscle. Am J Physiol 261:G952–G957
Washabau RJ, Wang MB, Dorst C, Ryan JP 1991 Effect of muscle length on isometric stress and myosin light chain phosphorylation in gallbladder smooth muscle. Am J Physiol 260:G920–G924
Xie YN, Gerthoffer WT, Reddy SN, Cominelli F, Eysselein VE, Snape WJ Jr 1992 An abnormal rate of actin myosin crossbridge cycling in colonic smooth muscle associated with experimental colitis. Am J Physiol 262:G921–G926
Gerthoffer WT, Murphy KA, Mangini J, Boman S, Lattanzio FA Jr 1991 Myosin phosphorylation and calcium in tonic and phasic contractions of colonic smooth muscle. Am J Physiol 260:G959–G964
Weisbrodt NW, Murphy RA 1985 Myosin phosphorylation and contraction of feline esophageal smooth muscle. Am J Physiol 249:C9–C14
O'Farrell PH 1975 High resolution two-dimensional electrophoresis of proteins. J Biol Chem 250: 4007–4021
Tallarida RJ, Murray RB 1987 Manual of Pharmacologic Calculations with Computer Programs. Springer-Verlag, New York, pp 93–97
Hellstrand P, Nordstrom I 1993 Cross-bridge kinetics during shortening in early and sustained contraction of intestinal muscle. Am J Physiol 265: 695–703
Hartshorne DJ, Kawamura T 1992 Regulation of contraction-relaxation in smooth muscle. News Physiol Sci 7: 59–64
Janis RA, Barany K, Barany M, Sarmiento JG 1981 Association between myosin light chain phosphorylation and contraction of uterine smooth muscle. Mol Physiol 1: 3–11
Dokhac L, D'Albis A, Janmot C, Harbon S 1986 Myosin li+ght chain phosphorylation in intact rat uterine smooth muscle: role of calcium and cyclic AMP. J Muscle Res Cell Motil 7: 259–268
Word RA, Stull JT, Casey ML, Kamm KE 1993 Contractile elements and myosin light chain phosphorylation in myometrial tissues from nonpregnant and pregnant women. J Clin Invest 92: 29–37
Rembold CM 1992 Regulation of contraction and relaxation in arterial smooth muscle. Hypertension 20: 129–137
Merkel L, Gerthoffer WT, Torphy TJ 1990 Dissociation between myosin phosphorylation and shortening velocity in canine trachea. Am J Physiol 258:C524–C532
Kamm KE, Murphy RA 1985 Velocity and myosin phosphorylation transients in arterial smooth muscle: effects of agonist diffusion. Experentia 41: 101–107
Triano-Rodgers T, Garbarina R, Martin JS, Ryan JP 1994 Postnatal development of gastric smooth muscle contractility is associated with changes in myosin heavy chain isoform distribution. Pediatr Res 35: 804
Ayas M, Bitar KN, Hillemeier C 1995 Characterization of actin and myosin in the developing stomach. Pediatr Res 37: 202–206
Author information
Authors and Affiliations
Additional information
Supported in part by National Institutes of Health Grant HD21047.
Rights and permissions
About this article
Cite this article
Ierardi, J., Paul, D. & Ryan, J. Myosin Light Chain Phosphorylation in Contraction of Gastric Antral Smooth Muscle from Neonate and Adult Rabbits. Pediatr Res 39, 156–159 (1996). https://doi.org/10.1203/00006450-199601000-00024
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-199601000-00024
