
Abstract
The presence of phosphorylated histone H2AX (γ-H2AX) is associated with the local activation of DNA-damage repair pathways. Although γ-H2AX deregulation in cancer has previously been reported, the molecular mechanism involved and its relationship with other histone modifications remain largely unknown. Here we find that the histone methyltransferase SUV39H2 methylates histone H2AX on lysine 134. When H2AX was mutated to abolish K134 methylation, the level of γ-H2AX became significantly reduced. We also found lower γ-H2AX activity following the introduction of double-strand breaks in Suv39h2 knockout cells or on SUV39H2 knockdown. Tissue microarray analyses of clinical lung and bladder tissues also revealed a positive correlation between H2AX K134 methylation and γ-H2AX levels. Furthermore, introduction of K134-substituted histone H2AX enhanced radio- and chemosensitivity of cancer cells. Overall, our results suggest that H2AX methylation plays a role in the regulation of γ-H2AX abundance in cancer.
Similar content being viewed by others

Introduction
The structural subunit of chromatin is the nucleosome, which consist of a histone octameric core constituted of four different histone types: H2A, H2B, H3 and H4. These nuclear histones can undergo a variety of chemical modifications such as acetylation, methylation, ubiquitination, sumoylation, poly ADP-ribosylation and phosphorylation1. The combination of these dynamic modifications form the so-called ‘histone code’, which influences gene expression, the DNA-damage response (DDR) and DNA repair2,3. Histone H2AX is a member of the histone H2A family and accounts for ~10% of total H2A molecules in normal human fibroblasts. However, the amounts of H2AX significantly vary between cell types4,5,6. H2AX plays a critical role in the DDR following induction of double-strand breaks (DSBs). When DSBs occur, H2AX accumulates near the DNA breakage sites and is quickly phosphorylated by members of the phosphatidyl-inositol-3-kinase-related kinases family, including ataxia telangiectasia-mutated (ATM), ataxia telangiectasia and Rad3-related (ATR) and DNA-activated protein kinase7. This phosphorylated form of histone H2AX is referred to as γ-H2AX and is a marker of DNA damage. γ-H2AX accumulates at sites of damaged chromatin within seconds of the formation of a DSB and triggers the accumulation of several components involved in the DDR signalling cascade8,9. In addition to phosphorylation, ubiquitination of H2AX has also been reported10. Several studies have highlighted the functions of RING finger ubiquitin ligases, RNF2, RNF8 and RNF168, in promoting accumulation of repair proteins at DSBs in an MDC1 (mediator of DNA-damage checkpoint protein 1)-dependent manner11,12,13,14. A number of in vitro and in vivo studies have demonstrated that phosphorylation and ubiquitination of H2AX play a central role in regulating various cellular responses to DSBs, including DNA repair and cell cycle checkpoints15,16. Furthermore, as DSBs are the most deleterious DNA damages that cause genomic instability and enhance the risk of tumorigenesis, deregulation of γ-H2AX seems to be linked to human cancer17,18.
Suppressor of Variegation 3–9 Homologue 2 (SUV39H2), also known as KMT1B19, is a SET-domain-containing methyltransferase that selectively methylates H3K9. The expression of Suv39h2—the murine homologue of human SUV39H2—is restricted to the testis in adult tissues and the endogenous Suv39h2 protein was shown to be enriched at heterochromatin during the first meiotic prophase and in the early stages of spermatogenesis. During mid-pachytene, Suv39h2 specifically accumulates with chromatin of the silenced sex chromosomes present in the XY body. In addition, Suv39h2 histone methyltransferase activity appears to play an important role in regulating higher-order chromatin dynamics during male meiosis20. On the other hand, the biological significance of SUV39H2 deregulation in human tumorigenesis is almost unknown. In the present study, we demonstrate that SUV39H2 methylates histone H2AX, and that the H2AX methylation is critical for γ-H2AX accumulation in human cancer.
Results
SUV39H2 methylates H2AX and is overexpressed in cancer
To investigate the possible function of methylation in the regulation of H2AX, we conducted in vitro methyltransferase assays against H2AX using a variety of histone methyltransferases. SUV39H2, a SET-domain-containing histone methyltransferase reported to methylate H3K9 (refs 20, 21), was found to be able to methylate histone H2AX (Fig. 1a). The histone methyltransferase activity of SUV39H2 appears to play an important role in regulating chromatin structure and dynamics, whereas the biological significance of SUV39H2 deregulation in human tumorigenesis is still largely unexplored. Hence, we investigated the role of SUV39H2 and its relation to H2AX modification in human cancers.

(a) In vitro methyltransferase analysis of SUV39H2. Recombinant histone H2AX and 3H-SAM were incubated in the presence or absence of recombinant SUV39H2, and the reaction products were analysed by SDS–PAGE followed by fluorography (upper panel) and stained for total protein(lower panel). (b) SUV39H2 mRNA levels in 14 lung cancer cases (NSCLC: 9 cases; SCLC: 5 cases) and 16 normal tissues. (c) Quantitative real-time PCR analysis was performed in 14 lung cancer samples and 16 normal tissues (the brain, breast, colon, oesophagus, eye, heart, liver, pancreas, rectum, spleen, stomach, kidney, bladder, testis, placenta and lung) and the result is shown by box-whisker plot. For statistical analysis, Kruskal–Wallis (*P<0.05) and Student’s t-test (**P<0.05) were performed. (d) Representative cases for positive SUV39H2 expression in lung cancer tissues and normal adult tissues. ADC, adenocarcinoma; SCC, squamous cell carcinoma. Original magnification, × 100 (lung × 200). (e) Expression levels of SUV39H2 in 65 normal lung samples, 19 large cell lung carcinoma samples, 45 lung adenocarcinoma samples and 27 squamous cell lung carcinoma samples. SUV39H2 is overexpressed in all three types of lung cancer. Expression profile is derived from Oncomine database.
We first examined expression levels of SUV39H2 in 16 normal and 14 lung cancer tissues (9 non-small-cell lung carcinoma (NSCLC) cases and 5 SCLC cases) using quantitative real-time PCR analysis, and found that SUV39H2 was significantly upregulated in cancer cells compared with that in normal tissues (Fig. 1b,c). We subsequently conducted immunohistochemical analysis of SUV39H2 in lung cancer and normal tissues, and found that SUV39H2 was overexpressed in 217 out of 328 archival NSCLC cases in accordance with the Oncomine database, while no staining was observed in normal organs, except for the testis (Fig. 1d,e). Expression profile analysis by complementary DNA microarray using a large number of clinical cases also revealed overexpression of SUV39H2 in cervical, bladder, oesophageal and prostate cancers, as well as in osteosarcomas and soft tissue sarcomas (Supplementary Table 1).
SUV39H2 is critical for chemo- and radiosensitivity
To investigate the role of SUV39H2 in human cancer and the relationship between SUV39H2 and γ-H2AX, we examined the effect of SUV39H2 knockdown on radio- and chemosensitivity of cancer cells, because γ-H2AX is considered as a key regulator of the DNA repair system after DSBs, which causes chemo/radio-resistance of cancer cells6. The lung squamous carcinoma RERF-LC-AI cells overexpressing SUV39H2 (Supplementary Fig. 1) was treated with control small interfering RNA (siRNA) or two different siRNAs directed to SUV39H2 for 48 h, followed by irradiation with 3 or 6 Gy of ionizing radiation. Subsequent cell viability analysis revealed that SUV39H2 knockdown enhanced the sensitivity of RERF-LC-AI cells to radiation (Supplementary Fig. 2a). We also examined the effect of SUV39H2 knockdown on chemosensitivity. RERF-LC-AI cells were transfected with control siRNA and either of two independent SUV39H2 siRNAs, and then treated with various concentrations of cisplatin or doxorubicin 48 h after siRNA transfection. Cell viability IC50 values after 48 h cisplatin treatment were calculated to be ~10 μM for the control siRNA, and 0.38 μM and 0.21 μM for siSUV39H1#1 and siSUV39H1#2, respectively. Those for the doxorubicin treatment were 2.21 μM for the control siRNA, and 0.096 and 0.066 μM for siSUV39H1#1 and siSUV39H1#2, respectively (Supplementary Fig. 2b), further supporting SUV39H2 as playing a role in chemosensitivity.
Lysine 134 on histone H2AX is dimethylated by SUV39H2
We next examined the relationship between SUV39H2-dependent histone H2AX methylation and γ-H2AX production. We first performed liquid chromatography-tandem mass spectrometry (MS/MS) to identify the site of methylation for histone H2AX, and found that lysine 134 was dimethylated by SUV39H2 (Fig. 2a). To validate this result, we prepared recombinant H2AX-K134A—where lysine 134 is substituted for alanine—and performed an in vitro methyltransferase assay (Fig. 2b). The intensity of the band corresponding to histone H2AX methylation was significantly diminished in the K134A mutant compared with that of wild-type H2AX (H2AX-WT), suggesting that lysine 134 of histone H2AX is a novel substrate of SUV39H2. Lysine 134 is located in a peptide region unique to H2AX among the H2A family members and is located near serine 139. Ser139-phosphorylated H2AX is known as γ-H2AX (Fig. 2c). As lysine 134 is highly conserved from Danio rerio to Homo sapiens, we postulated that methylation of this site might be critical for the regulation of histone H2AX functions and the production of γ-H2AX (Fig. 2d).

(a) The MS/MS spectrum corresponding to the dimethylated histone H2AX peptide (residues 128–142). A 28-Da increase indicates dimethylated Lys 134. Score and Expect show Mascot Ion Score and Expectation value in Mascot Database search results are shown, respectively. (b) Validation of K134 methylation on histone H2AX. Recombinant histone H2AX-WT or H2AX-K134A proteins and 3H-SAM were incubated in the presence of recombinant SUV39H2, and the reaction products were analysed by SDS–PAGE followed by fluorography. The membrane was stained with Ponceau S (lower panel). (c) Amino acid sequence alignment of human histone H2A family. Lysine 134 is located in the unique sequence portion of H2AX. (d) Amino acid sequence alignment of H2AX unique sequence portion. (e) Determination of the titre and specificity of the anti-dimethylated K134 H2AX antibody analysed by enzyme-linked immunosorbent assay. (f) Validation of the anti-dimethylated K134 H2AX antibody. Recombinant H2AX-WT or H2AX-K134A proteins and SAM were incubated in the presence or absence of recombinant SUV39H2, and the reaction products were analysed by WB analysis. The intensity of each H2AX K134 dimethylation signal was normalized by the corresponding H2AX. SAHH, S-adenosyl-L-homocysteine hydrolase. Results are the mean±s.d. (n=3). (g) Immunocytochemical analysis of HeLa cells. Cells were stained with an anti-FLAG antibody (green), an anti-H2AX K134me2 antibody (red) and 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI (blue)). Non-transfected HeLa cells were used as negative control. Scale bars, 10 μm. (h) 293T cells were co-transfected with a FLAG-H2AX-WT or a FLAG-H2AX-K134A, and an empty vector (HA-Mock) or HA-SUV39H2. The samples were immunoblotted with anti-dimethylated K134 H2AX and anti-FLAG antibodies following IP with anti-FLAG. Results are the mean±s.d. (n=3).
To investigate this possibility, we generated an antibody against a synthetic peptide containing dimethylated K134 and confirmed its affinity and specificity by enzyme-linked immunosorbent assay (Fig. 2e). To verify the quality of the H2AX K134-methylation antibody, we prepared both H2AX-WT and H2AX-K134A recombinant proteins, and performed in vitro methyltransferase assays using recombinant SUV39H2 protein as an enzyme source. The methylation signal of H2AX-WT was clearly observed in the presence of SUV39H2, while the signal was absent when H2AX-K134A protein was used as a substrate (Fig. 2f). Using this antibody, we performed immunocytochemical analysis after transfection of a SUV39H2 expression vector into HeLa cells, to further verify SUV39H2-dependent H2AX K134 methylation in vivo, and detected a strong staining of K134-methylated H2AX in SUV39H2-overexpressing HeLa cells (Fig. 2g). Furthermore, we expressed FLAG-H2AX-WT or a FLAG-H2AX K134A, along with an influenza hemagglutinin (HA) control or HA-SUV39H2, into 293T cells. Western blot analysis of FLAG-immunoprecipitated protein with anti-K134-methylated H2AX exhibited a specific signal when both H2AX-WT and SUV39H2 proteins were co-expressed, but no signal was detected in H2AX K134A mutant (Fig. 2h). Taken together, these data indicate that SUV39H2 methylates histone H2AX on lysine 134 both in vitro and in vivo.
H2AX K134 methylation is critical for γ-H2AX production
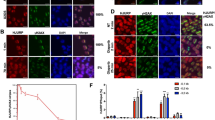
To examine the physiologic significance of H2AX K134 methylation for γ-H2AX production, we transfected either of siNegative control (siNC; control), siEGFP (control), SUV39H2-siRNA#1 or SUV39H2-siRNA#2 into RERF-LC-AI cells. Forty-eight hours after transfection, the cells were treated with 1 μM doxorubicin for 2 h. Western blot analysis detected high levels of γ-H2AX in the cells treated with the control siRNAs, but significantly less γ-H2AX was present when the cells were pre-treated with siSUV39H2s (Fig. 3a,b). The observed decrease in γ-H2AX production following SUV39H2 knockdown appeared not to be caused by changes in quantity or activity of ATM and ATR kinases, as the amounts of ATM and ATR—responsible for H2AX serine 139 phosphorylation—as well as those of phospho-ATM (active-form of ATM) and phospho-Chk2 (Checkpoint kinase 2, a substrate of ATM and ATR) were unchanged. Importantly, given that the suppression of H2AX K134 dimethylation following knockdown of SUV39H2 was clearly observed in the endogenous H2AX proteins after immunoprecipitation (IP) with an H2AX antibody (Fig. 3c), SUV39H2 appears to be a fundamental regulator of endogenous H2AXK134 dimethylation. In addition, we confirmed that doxorubicin-induced DSBs did not affect SUV39H2 expression and H2AX K134 methylation levels (Fig. 3d). To validate the effect of SUV39H2 on γ-H2AX production, we performed immunocytochemical analysis using the anti-γ-H2AX antibody and detected significant decrease of γ-H2AX levels following knockdown of SUV39H2 (Fig. 3e,f), consistent with the western blotting (WB) results. These data suggest that SUV39H2 plays a critical role in γ-H2AX activation. We next prepared Suv39h2-null (Suv39h2−/−) mouse embryonic fibroblast (MEF) cells from Suv39h2 knockout mouse22, to confirm the knockdown experiments. Both Suv39h2−/− and Suv39h2-WT MEFs were treated with 1 μM doxorubicin for 2 h before lysis. Western blot analysis confirmed significantly lower levels of γ-H2AX activity in Suv39h2−/− MEFs compared with that in Suv39h2-WT MEFs (Supplementary Fig. 3). Moreover, we examined laser-induced γ-H2AX activity in Suv39h2−/− and Suv39h2-WT MEFs, and confirmed that Suv39h2−/− cells showed reduced γ-H2AX compared with Suv39h2-WT cells (Supplementary Fig. 4).

(a) Effects of SUV39H2 knockdown on the γ-H2AX levels. RERF-LC-AI cells were transfected with two control siRNAs (siNC and siEGFP) and SUV39H2 siRNAs (#1 and #2), and were treated with 1 μM of doxorubicin 48 h after transfection with siRNAs. Cells were harvested 2 h after treatment with doxorubicin, followed by SDS–PAGE. Western blotting was performed using anti-SUV39H2, anti-SUV39H1, anti-dimethylated K134 H2AX, anti-γ-H2AX, anti-H2AX, anti-phospho-ATM, anti-ATM, anti-ATR, anti-phosph-Chk2 and anti-α-Tubulin (internal control) antibodies. (b) The intensity of γ-H2AX levels was normalized by α-Tubulin and averaged. Results are the mean±s.d. (n=3). P-values were calculated using Student’s t-test (*P<0.05). (c) RERF-LC-AI cells were transfected with two control siRNAs (siNC and siEGFP) and SUV39H2 siRNAs (#1 and #2), and were treated with 1 μM of doxorubicin 48 h after transfection with siRNAs. Cells were harvested 2 h after treatment with doxorubicin and IP was conducted using an anti-H2AX antibody. Input and immunoprecipitated samples were immunoblotted with anti-SUV39H2, anti-α-Tubulin, anti-dimethylated K134 H2AX and anti-H2AX antibodies. (d) RERF-LC-AI and HeLa cells were treated with 1 μM of doxorubicin for 2 h before lysis. The samples were immunoblotted with anti-SUV39H2, anti-H2AXK134me2 and anti-α-Tubulin antibodies. The intensity of SUV39H2 and H2AK134me2 levels was normalized by α-Tubulin expression levels. Results are the mean±s.d. (n=3). (e) Immunocytochemical analysis of RERF-LC-AI cells. Cells were transfected with control siRNA and SUV39H2 siRNA, and were treated with 1 μM of doxorubicin 48 h after transfection with siRNAs for 1 h. Cells were stained with an anti-SUV39H2 antibody (green), an anti-γ-H2AX antibody (red) and 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI (blue)) 2 h after treatment with doxorubicin. Scale bar, 10 μm. (f) γ-H2AX intensity examined in e was quantified with the Image J software and averaged. Results are the means±s.d. of ten independent cells. P-values were calculated using Student’s t-test (***P<0.001).
To further evaluate the effect of H2AX K134 methylation on γ-H2AX production in vivo, we co-expressed FLAG-H2AX-WT or FLAG-H2AX-K134A with HA-SUV39H2 in HeLa cells before treatment with doxorubicin. IP and WB analysis show that γ-H2AX was induced to a higher level in the presence of H2AX-WT, compared with two H2AX-K134A or K134R (Fig. 4a,b and Supplementary Fig. 5). In addition, we found that the amount of phospho-ATM and ATR bound to H2AX were decreased in K134-substituted H2AX. Furthermore, we co-expressed H2AX-WT or H2AX-K134A with SUV39H2 in H2AX−/− MEFs to validate the relationship between H2AX K134 methylation and γ-H2AX production. First, we confirmed that overexpressed H2AX-WT and H2AX-K134A were appropriately incorporated into chromatin of H2AX−/− MEF cells by immunocytochemistry (ICC; Supplementary Fig. 6), and then conducted WB analysis following purification of the chromatin fraction (Supplementary Fig. 7). γ-H2AX production in H2AX−/− MEF cells overexpressing H2AX-K134A was clearly suppressed compared with H2AX−/− MEF cells overexpressing H2AX-WT, confirming the critical role of H2AX K134 dimethylation in γ-H2AX production.

(a) FLAG-H2AX-WT or a FLAG-H2AX-K134A were co-expressed with HA-SUV39H2 and treated with 1 μM of doxorubicin 48 h post transfection. The samples were immunoblotted with anti-H2AXK134me2, anti-γ-H2AX, anti-FLAG, anti-phospho ATM and ATR antibodies after immunoprecipitating with anti-FLAG M2 agarose. Input protein levels of phospho-ATM, ATR, α-Tubulin and HA are also shown. (b) Quantification of WB results shown in a. γ-H2AX, phospho ATM and ATR levels after doxorubicin treatment normalized by FLAG or α-Tubulin expression levels and averaged. Results are the mean±s.d. (n=3). (c) In vitro kinase and pull-down assay. Biotin-tagged unmodified H2AX peptides or biotin-tagged K134 dimethylated H2AX peptides were mixed with enzyme source (whole-cell lysates of HeLa cells). Peptides were precipitated and immunoblotted with an anti-γ-H2AX antibody. (d) The intensity of γ-H2AX signal was normalized by peptide amount and averaged. Results are the means±s.d. (n=3). (e) Comparison of ATR and phospho-ATM amounts bound to unmodified-H2AX peptides and K134-dimethylated H2AX peptides. Biotin-tagged peptides were mixed with 5 μl of enzyme source (whole-cell lysates of HeLa cells). Peptides were precipitated and immunoblotted with anti-ATR and anti-phospho-ATM antibodies. Results are the means±s.d. (n=3). (f) The schematic drawing of strategy for in vitro kinase assays. (g) Biotin-tagged unmodified H2AX peptides, biotin-tagged K134-dimethylated H2AX peptides and biotin-tagged K134R H2AX peptides were mixed with enzyme source (immunoprecipitated ATM) in the presence or absence of KU-55933. The samples were immunoblotted with an anti-γ-H2AX antibody.
To explore the relationship between H2AX K134 methylation and Η2ΑX S139 phosphorylation in more detail, we prepared biotin-conjugated unmodified H2AX and K134 dimethylated H2AX peptides encompassing amino acids 121–142 (Fig. 4c), and conducted in vitro kinase and pull-down assays. Biotin-conjugated peptides were mixed with lysates from HeLa cells treated with doxorubicin and pulled down with Dynabeads M-280 streptavidin followed by WB analysis (Fig. 4c,d). The K134 dimethylated H2AX peptide exhibited a much higher level of H2AX S139 phosphorylation than the unmodified H2AX peptide. We also observed that the amount of phospho-ATM and ATR bound to the K134-dimethylated H2AX peptide was significantly higher compared with the unmodified peptide, consistent with the above data (Fig. 4e). Moreover, we also conducted the in vitro kinase assay of H2AX peptides using immunoprecipitated ATM as the source of enzyme (Fig. 4f). The K134-dimethylated peptide showed notably higher H2AX S139 phosphorylation compared with the unmodified and K134R peptides (Fig. 4g). In addition, KU-55933, an ATM kinase inhibitor, diminished H2AX S139 phosphorylation of the K134-dimethylated peptide. These results further support our hypothesis that dimethylated K134 is critical for H2AX S139 phosphorylation.
We next examined the effect of H2AX K134 methylation on the localization of the tumour suppressor p53-binding protein 1 (TP53BP1)—one of the important mediators within the DSB repair machinery—following treatment with doxorubicin. As previously observed1,6, TP53BP1 formed foci that co-localized with γ-H2AX in the WT MEFs; however, it was widely distributed in the Suv39h2−/− MEFs (Fig. 5a). Similarly, TP53BP1 localization was lost in RERF-LC-AI cells when SUV39H2 was knocked down (Fig. 5b).

(a) Immunocytochemical analysis of MEF-WT and MEF-Suv39h2−/−. Cells were treated with 1 μM of doxorubicin for 1 h and stained with an anti-γ-H2AX antibody (red), an anti-TP53BP1 antibody (green) and 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI; blue). Scale bar, 10 μm. (b) Immunocytochemical analysis of RERF-LC-AI cells after knockdown of SUV39H2. Cells were transfected with siNC and siSUV39H2. After 48 h, siRNA-transfected cells were treated with 1 μM of doxorubicin for 1 h and stained with an anti-SUV39H2 antibody (red), an anti-TP53BP1 antibody (green) and DAPI (blue). Scale bar, 10 μm. (c) Immunohistochemical stainings of K134-methylated H2AX and γ-H2AX in clinical lung tissues. Typically stained normal and tumour tissues are shown. Detailed clinical information and data are described in Supplementary Table 2. (d) Correlation of staining between K134-methylated H2AX and γ-H2AX was statistically calculated using Spearman’s correlation coefficient by ranks with ties. (e) H2AXK134 methylation and γ-H2AX were co-expressed in clinical bladder tissues. Immunohistochemical stainings of K134-methylated H2AX and γ-H2AX in clinical bladder tissues. Typically stained normal and tumour tissues are shown. Detailed clinical information and data are described in Supplementary Table 3. (f) Correlation of staining between K134-methylated H2AX and γ-H2AX in clinical bladder tissues was statistically calculated using Spearman’s correlation coefficient by ranks with ties.
Correlation of H2AX K134 methylation and γ-H2AX in cancer
To examine the relation between H2AX K134 methylation and γ-H2AX levels in clinical tissues, we conducted immunohistochemical analysis using tissue microarray comprising 154 cases of lung cancer. As shown in Fig. 5c, the signal intensity of both H2AX K134 methylation and γ-H2AX was notably higher in lung cancer tissues compared with normal tissues. We calculated the Spearman’s correlation coefficient by ranks with ties, to clarify the statistical correlation between the two histone H2AX modifications. The Spearman’s ρ-value of 0.823 (P<0.0001; Fig. 5d and Supplementary Table 2) supports a direct correlation between H2AX K134 methylation and γ-H2AX levels. This result was also confirmed in clinical bladder tissues (Fig. 5e,f and Supplementary Table 3).
H2AX K134 methylation regulates chemo- and radiosensitivity
To further evaluate the effect of H2AX K134 methylation on γ-H2AX production and DNA repair, we established a dominant-negative experimental system. HeLa cells were transfected with either FLAG-H2AX-WT or FLAG-H2AX-K134A, with or without co-expression of HA-SUV39H2. After 48 h, cells were treated with 1 μM Doxorubicin for 2 h and harvested for WB analysis (Fig. 6a). SUV39H2 overexpression clearly enhanced both H2AX K134 dimethylation and S139 phosphorylation on the H2AX-WT proteins, but not the in K134-substituted proteins (Fig. 6a). We repeated the WB analysis (Supplementary Fig. 8) and quantified levels of H2AX K134 dimethylation and γ-H2AX (Fig. 6b), which showed a strong correlation between the histone H2AX K134 methylation and γ-H2AX. To examine whether exogenous H2AX proteins are appropriately incorporated into chromatin, HeLa cells transfected with FLAG-H2AX-WT or FLAG-H2AX-K134 were stained with anti-FLAG antibody after synchronizing the cells (Fig. 6c). The FLAG-tagged H2AX-WT and H2AX-K134A proteins that overlapped the strong 4′,6-diamidino-2-phenylindole stainings were detected in the heterochromatin regions in the nucleus23,24. In mitotic cells, we found co-localization of the exogenous H2AX proteins with condensed chromosomes, further indicating that the exogenous H2AX-WT and H2AX-K134A proteins were appropriately incorporated into the chromatin. We also confirmed the incorporation of exogenous H2AX-WT and H2AX-K134A into chromatin by WB after chromatin fractionation (Supplementary Fig. 9).

(a) HeLa cells were transfected with a FLAG-H2AX-WT, FLAG-H2AX-K134A or FLAG with HA-SUV39H2 or an HA-control. After 48 h, cells were treated with 1 μM of doxorubicin for 1 h and samples immunoblotted with anti-H2K134me2, anti-γ-H2AX, anti-H2AX, anti-HA (SUV39H2) and anti-α-Tubulin antibodies. (b) Quantification of exogenous and endogenous K134-dimethylated or γ-H2AX amount. The intensity of K134-dimethylated H2AX and γ-H2AX levels was normalized against H2AX. Results are the mean±s.d. (n=3). Total amount shows the sum of endogenous and exogenous expression levels. Results are the mean±s.d. (n=3). (c) HeLa cells were transfected with FLAG-H2AX-WT or FLAG-H2AX-K134A, and treated with 7.5 (μg ml−1) of aphidicolin to synchronize the cell cycle 48 h after transfection. Next, the cells were stained with an anti-FLAG antibody ( green) and 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI; blue) 13 h after release from G1 arrest. Scale bars, 10 μm.
Using this dominant-negative system, we also examined the effect of K134 methylation on radio- and chemosensitivity. H2AX-K134A-overexpressing HeLa cells exhibited significantly lower viability than H2AX-WT-overexpressing HeLa cells following radiation treatment (Fig. 7a,b). Sensitivity was also shown to be dose dependent by colony-formation assay (Fig. 7c and Supplementary Fig. 10). We examined the cell cycle status in the two cell lines without radiation and found no difference (Fig. 7d,e). With regard to the effect of H2AX K134 methylation on chemosensitivity, IC50s of both cisplatin and doxorubicin treatment were significantly lower in the H2AX-K134A-overexpressed HeLa cells (cisplatin: WT=4.82 μM, K134A=1.26 μM; doxorubicin: WT=4.82 μM, K134A=1.26 μM; Fig. 7f). In summary, attenuation of H2AX K134 methylation appears to enhance both radio- and chemosensitivity of cancer cells.

(a) HeLa cells were transfected with a FLAG-H2AX-WT or a FLAG-H2AX-K134A expression vector and HA-SUV39H2, then irradiated with 6 or 10 Gy of ionizing radiation 24 and 72 h post transfection. Cell viability was measured 24 h after the second irradiation. Results are the mean±s.d. (n=3). P-values were calculated using Student’s t-test (**P<0.01). n.s., not significant. (b) Expression of FLAG-H2AX-WT and FLAG-H2AX-K134A in HeLa cells from a. Samples were immunoblotted with anti-FLAG and anti-α-Tubulin antibodies. (c) Clonogenicity assays of HeLa cells transfected with H2AX-WT and H2AX-K134A. Cells were irradiated with 0, 2, 4, 6 and 10 Gy of ionizing radiation 24 h post transfection. Subsequently, the cells were cultured in EMEM with 0.8 mg ml−1 of Geneticin/G-418 for 15 days. Results are the means±s.d. (n=3). P-values=Student’s t-test (**P<0.01). (d) Detailed cell cycle kinetics in HeLa cells 48 h after transfection with FLAG-H2AX and FLAG-K134A. (e) Expression of FLAG-H2AX-WT and FLAG-H2AX-K134A in HeLa and A549 cells from d. (f) HeLa cells overexpressing FLAG-H2AX-WT or FLAG-H2AX-K134A with HA-SUV39H2 were treated with cisplatin or doxorubicin 24 h post transfection. Cell viability was measured 48 h after the drug treatment.
Discussion
In the present study, we showed that the histone methyltransferase SUV39H2 methylates histone H2AX on lysine 134, and that this is critical for the production of γ-H2AX in cancer cells (Supplementary Fig. 11). Our binding assays revealed that the lack of methylation at K134 reduces the affinity between the kinases responsible for γ-H2AX production and H2AX (Fig. 4b,e).
SUV39H2 can also methylate histone H3 lysine 9 and this methylation is considered to be important for organizing heterochromatin20. Given that chromatin structure is likely to be important for DNA repair processes, the dynamics of relevant histone modifications during this step may play important roles in organizing the process appropriately. Indeed, histone H3 K9 methylation was also reported to be a regulator of the DNA repair pathway25,26, indicating that SUV39H2 might regulate this pathway through the methylation of both histone H2AX K134 and H3 K9. Further functional analyses may clarify the dynamics of histone modifications, the involvements of other proteins and their mutual relations in the DDR following induction of DSBs in cancer cells.
It has previously been reported that constitutive endogenous γ-H2AX foci are rare in normal primary human cells and tissues27. However, cancer cells and tissues show different degrees of constitutive γ-H2AX activity in the absence of exogenously induced DSBs27,28. Moreover, γ-H2AX is also a prognostic marker for several types of human cancer, and increased γ-H2AX levels are significantly associated with poorer prognosis of triple-negative breast cancer, endometrial cancer and NSCLC28,29,30,31. These results imply that high γ-H2AX expression levels may be involved in a more aggressive, highly proliferating tumour phenotype and resistance to anti-cancer treatment30. Given that SUV39H2 is significantly upregulated in various types of tumour tissues (Supplementary Table 1), it is possible that constitutive SUV39H2 overexpression in cancer cells may cause aberrant γ-H2AX expression, and that cancer cells might acquire more malignant phenotype, including chemo- and/or radioresistance.
Importantly, γ-H2AX is considered to be a drug target32,33,34,35,36,37,38 and peptide inhibitors of γ-H2AX have already been reported as potential chemotherapeutic agents35,39. Inhibition of γ-H2AX through interference with upstream kinase activities using caffeine, wortmannin and LY294002 resulted in significantly increased tumour cell radiosensitivity40. In addition, VE-821, a novel specific inhibitor of ATR41, increased sensitivity of cancer cells to radiation and/or chemotherapy42,43,44,45. These findings suggest that regulation of γ-H2AX production through the inhibition of SUV39H2-depedent H2AX K134 methylation could be an attractive target for drug development. Indeed, as the expression level of SUV39H2 in normal tissues is low (Fig. 1d, and Supplementary Figs 12 and 13), this molecule could represent an ideal target for cancer therapy. Moreover, our knockdown experiments using Cell Counting kit-8 (CCK-8), Giemsa and fluorescence-activated cell sorting and clonogenicity assays revealed that SUV39H2 appears to possess oncogenic activity (Supplementary Fig. 14). According to our expression profile analysis, SUV39H1 is also overexpressed in some cancers. Moreover, knockdown of SUV39H2 significantly diminished the γ-H2AX production even though SUV39H1 expression levels were not changed (Fig. 3a), suggesting that SUV39H2 is a critical factor that regulates γ-H2AX production. Taking these findings into account, we propose a SUV39H2 inhibitor could be used alone or in combination with DNA-damaging agents or radiotherapy in cancer patients with aberrant SUV39H2 expression.
Methods
Cell culture and clinical tissues
NCI-H1781, A549, SK-MES-1, NCI-H2170, NCI-H520, DMS 114, 293T, HeLa, SW780 and COS-7 cells were from the American Type Culture Collection in 2001 and 2003, and were tested and authenticated by DNA profiling for polymorphic short tandem-repeat markers (Supplementary Table 4). SBC-3, SBC-5 and RERF-LC-AI cells were from the Japanese Collection of Research Bioresources in 2001 and were tested and authenticated by DNA profiling for polymorphic short tandem-repeat markers (Supplementary Table 4). ACC-LC-319 cells were from the Aichi Cancer Center in 2003, and were tested and authenticated by DNA profiling for single-nucleotide polymorphism, mutation and deletion analysis (Supplementary Table 4). The SW780 line was established in 1974 by A. Leibovitz from a grade I transitional cell carcinoma.
All cell lines were grown in monolayers in appropriate media: DMEM for RERF-LC-AI, 293T and COS-7 cells; Eagle’s minimum essential medium for SK-MES-1, SBC-3, SBC-5 and HeLa cells; RPMI1640 medium for NCI-H1781, ACC-LC-319, A549, NCI-H2170, NCI-H520 and DMS 114 cells; Leibovitz’s L-15 for SW780 cells supplemented with 10% fetal bovine serum (FBS) and 1% antibiotic/antimycotic solution (Sigma-Aldrich). SAEC cells were maintained in small airway epithelial cell basal medium supplemented with 52 μg ml−1 bovine pituitary extract, 0.5 ng ml−1 human recombinant epidermal growth factor, 0.5 μg ml−1 hydrocortisone, 0.5 μg ml−1 epinephrine, 10 μg ml−1 transferrin, 5 μg ml−1 insulin, 0.1 ng ml−1 retinoic acid, 6.5 ng ml−1 triiodothyronine, 50 μg ml−1 Gentamicin/Amphotericin-B (GA-1000) and 50 μg ml−1 fatty acid-free BSA. All cells were maintained at 37 °C in humid air with 5% CO2 condition, except for SW780 cells (without CO2). Cells were transfected with FuGENE 6 or FuGENE HD (Roche Applied Science) according to manufacturer’s protocols46,47. The use of all clinical materials in this study was approved by Ethics Committees of Institute of Medical Science at the University of Tokyo.
Antibodies
The following primary antibodies were used: anti-FLAG (rabbit, F7425; Sigma-Aldrich; dilution used in WB: 1:3,000, ICC: 1:500), anti-FLAG (mouse, M2; Sigma-Aldrich; dilution used in ICC: 1:500), anti-HA (Y-11; Santa Cruz Biotechnology; dilution used in WB: 1:500), anti-SUV39H1 (#8729; Cell Signaling Technology; dilution used in WB: 1:500), anti-H2AX (07-627; Millipore; dilution used in WB: 1:500, IP: 1:250), anti-γ-H2AX (05-636; Millipore; dilution used in WB: 1:500, ICC: 1:250, immunohistochemistry: 1:250), anti-ATM (ab2629; Abcam; dilution used in WB: 1:500), anti-phospho-ATM (ab81292; Abcam; dilution used in WB: 1:500), anti-ATR (#2790; Cell Signaling Technology; dilution used in WB: 1:500), anti-phospho-Chk2 (#2661; Cell Signaling Technology; dilution used in WB: 1:500), anti-α-Tubulin (T6199; Sigma-Aldrich; dilution used in WB: 1:500), anti-TP53BP1 (Clone BP13; Millipore; dilution used in ICC: 1:250) for RERF-LC-AI cells, anti-TP53BP1 (#4937; Cell Signaling Technology; dilution used in ICC: 1:250) for mouse MEF cells, anti-H2A (ab13923; Abcam; dilution used in WB: 1:1,000), anti-histone H3 (ab1791; Abcam; dilution used in WB: 1:1,000), anti-histone H4 (#2592S; Cell Signaling Technology; dilution used in WB: 1:1,000), anti-ATM (Ab-3; Calbiochem; dilution used in IP: 1:100) for IP and anti-ACTB (#2592S; Cell Signaling Technology; dilution used in WB: 1:1,000). An anti-SUV39H2 antibody (Sigma-Aldrich; dilution used in WB: 1:250, ICC: 1:250) and an anti-K134 di-methylated H2AX antibody (Sigma-Aldrich; dilution used in WB: 1:500, ICC: 1:250, immunohistochemistry: 1:250) were produced in rabbit immunized with a synthetic peptide.
Quantitative real-time PCR
For quantitative real-time PCR reactions, specific primers for all GAPDH (housekeeping gene), SDH (housekeeping gene) and SUV39H2 were designed (primer sequences in Supplementary Table 5). PCR reactions were performed using the LightCycler 480 System (Roche Applied Science) and ViiA 7 real-time PCR system (Life Technologies, Carlsbad, CA), following the manufacturer’s protocol. Messenger RNA levels were normalized to GAPDH and SDH expression.
In vitro methyltransferase assay
For the in vitro methyltransferase assay, recombinant Histone H2AX (#14-576, Millipore) was incubated with recombinant SUV39H2 enzyme using 1 μCi S-adenosyl-L-[methyl-3H]-methionine (SAM; PerkinElmer) as the methyl donor in a mixture of 10 μl of methylase activity buffer (50 mM Tris-HCl at pH8.8, 10 mM dithiothreitol and 10 mM MgCl2), for 2 h at 30 °C48,49,50,51. Proteins were resolved on a 5–20 % SDS–PAGE gel (Ready Gel; Bio-Rad) and visualized by MemCode Reversible Stain (Thermo Scientific) and fluorography.
Mass spectrometry
Histone H2AX samples that reacted with SUV39H2 were separated on SDS–PAGE and stained with Simply Blue Safe Stain (Life Technologies). The excised Histone H2AX bands were reduced in 10 mM tris (2-carboxyethyl) phosphine (Sigma-Aldrich) with 50 mM ammonium bicarbonate (Sigma-Aldrich) for 30 min at 37 °C and alkylated in 50 mM iodoacetamide (Sigma-Aldrich) with 50 mM ammonium bicarbonate for 45 min in the dark at 25 °C. Trypsin GOLD (Promega) solution was added with the enzyme to protein ratio at 1/50 (w/w) and incubated at 37 °C for 16 h. The resulting peptides were extracted from gel fragments and separated on a 0.1 × 200 mm homemade C18 column using 45 min linear gradient from 2% to 35% acetonitrile in 0.1% formic acid, with flow rate at 200 nl min−1. The eluting peptides were analysed with HCTultra ETD II mass spectrometer (Bruker Daltonics). The acquired MS and collision-induced dissociation (CID) MS/MS spectra were processed with Compass DataAnalysis 4.0 (Bruker Daltonics) and BioTools 3.1 software (Bruker Daltonics), followed by the database search on in-house Mascot server ver.2.3.01 (Matrix Science). We accepted the peptide identifications satisfying the Expectation value <0.05 in Mascot Database search.
Western blotting
Samples were prepared from the cells lysed with CelLytic M cell lysis reagent (Sigma-Aldrich) containing a complete protease inhibitor cocktail (Roche Applied Science)51, and whole cell lysates or IP products were transferred to nitrocellulose membrane. Protein bands were detected by incubating with horseradish peroxidase (HRP)-conjugated antibodies (GE Healthcare) and visualizing with enhanced chemiluminescence (GE Healthcare). We declare that our blots were evenly exposed in each membrane and that the blots are not clopped to the bands.
Immunocytochemistry
Cultured cells were fixed in 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) at room temperature for 30 min, permeabilized in 0.1% Triton X-100 (Sigma-Aldrich) for 3 min on ice and blocked with 3% BSA for 1 h at room temperature49. Fixed cells were incubated with primary antibodies overnight at 4 °C, incubated with Alexa Fluor-conjugated secondary antibody (Molecular Probes, Life Technologies) and observed using a Leica confocal microscopy (SP5 Tandem Scanner Spectral 2-Photon Confocal).
Immunohistochemical analysis
SUV39H2 expression and status of histone H2AX K134 methylation status and S139 phosphorylation in clinical tissues were examined by immunohistochemical analysis49,50,52,53,54,55,56,57. EnVision+ kit/HRP kit (Dako) was applied and slides of paraffin-embedded lung tumour specimens were processed under high pressure (125 °C, 30 s) in antigen-retrieval solution, high pH 9 (S2367, Dako), treated with peroxidase-blocking reagent and then treated with protein-blocking reagent (K130, X0909, Dako). Tissue sections were incubated with a rabbit anti-SUV39H2, a rabbit anti-H2AXK134me2 polyclonal antibody and a mouse anti-γ-H2AX antibody, followed by HRP-conjugated secondary antibody (Dako). Antigen was visualized with substrate chromogen (Dako liquid DAB chromogen; Dako). Finally, tissue specimens were stained with Mayer’s haematoxylin (Hematoxylin QS; Vector Laboratories, Burlingame, CA, USA) for 20 s to discriminate the nucleus from the cytoplasm. The intensity of H2AXK134 methylation and H2AXS139 phosphorylation was evaluated using the following criteria: strong positive (scored as 2+), brown staining in >50% of tumour cells completely obscuring cytoplasm; weak positive (1+), any lesser degree of brown staining appreciable in tumour cell cytoplasm; and absent (scored as 0), no appreciable staining in tumour cells. Cases were accepted as strongly positive if two or more investigators independently defined them as such.
Fluorescence-activated cell sorting
SW780 and A549 cells were treated with siSUV39H2 or control siRNAs (siEGFP and siNC) and cultured in a CO2 incubator at 37 °C for 72 h. Aliquots of 1 × 105 cells were collected by trypsinization and stained with propidium iodide following the manufacturer’s instructions (Cayman Pharma, Neratovice, Czech Republic). Cells were analysed by FACScan (Beckman Coulter, Brea, CA) with MultiCycle for Windows software (Beckman Coulter) for detailed cell cycle status58. The percentages of cells in G0/G1, S and G2/M phases of the cell cycle were determined from at least 20,000 ungated cells.
Coupled cell cycle and cell proliferation assay
A 5′-bromo-2′-deoxyuridine (BrdU) flow kit (BD Pharmingen, San Diego, CA) was used to determine the cell cycle kinetics and to measure the incorporation of BrdU into DNA of proliferating cells48,54,59,60,61. The assay was performed according to the manufacturer’s protocol. Briefly, cells were cultured in the presence of 10 μM of BrdU for 30 min at 37 °C. Next, both floating and adherent cells were pooled from triplicate wells per treatment point, fixed in a solution containing paraformaldehyde and the detergent saponin, and incubated for 1 h with DNase at 37 °C (30 μg per sample). Fluorescein isothiocyanate-conjugated anti-BrdU antibody (1:50 dilution in Wash buffer; BD Pharmingen) was added and incubation continued for 20 min at room temperature. Cells were washed in Wash buffer and total DNA was stained with 7-amino-actinomycin D (20 μl per sample), followed by flow cytometric analysis using FACScan (Beckman Coulter) and total DNA content (7-amino-actinomycin D) was determined by using CXP Analysis Software Ver. 2.2 (Beckman Coulter).
Immunoprecipitation
Transfected 293T cells or HeLa cells were lysed with CelLytic M cell lysis reagent (Sigma-Aldrich) containing a complete protease inhibitor cocktail (Roche Applied Science). In a typical IP reaction, 300 μg of whole-cell extract was incubated with an optimum concentration of primary antibody. After the beads had been washed three times in 1 ml of TBS buffer (pH 7.6), proteins that bound to the beads were eluted by boiling in Lane Marker Reducing Sample Buffer (Thermo Scientific).
siRNA transfection
siRNA oligonucleotide duplexes were purchased from Sigma-Aldrich for targeting the human SUV39H2 transcript. siNC was used as a control siRNA. The siRNA sequences are described in Supplementary Table 6. siRNA duplexes (100 nM final concentration) were transfected into bladder and lung cancer cell lines with Lipofectamine 2000 (Life Technologies)62,63,64.
Mouse embryonic fibroblast
Mouse Suv39h2 WT and Suv39h2−/− MEF cells were established by Dr Thomas Jenuwein group22. Cells were cultured with DMEM supplemented with 10% FBS, 1% antibiotic/antimycotic solution (Sigma-Aldrich), 2 mM L-glutamine, 0.1 mM β-mercaptoethanol, 1 × non-essential amino acid solution (Life Technologies) and sodium pyruvate. Phenotype of MEF cells were confirmed by CCK-8 (Dojindo) and coupled cell cycle and cell proliferation assay. Mouse H2AX−/− MEF cells were established by Dr Andre Nussenzweig group9 and cells were cultured with DMEM supplemented with 10% FBS and 1% antibiotic/antimycotic solution (Sigma-Aldrich). We confirmed that exogenous FLAG-tagged H2AX-WT and H2AX-K134A proteins were appropriately incorporated into chromatin (Supplementary Fig. 6).
In vitro kinase and pull-down assay
HeLa cells were lysed with CelLytic M cell lysis reagent (Sigma-Aldrich) containing protease and phosphatase inhibitors 2 h after treatment with 1 μM of doxorubicin and used as enzyme source. Biotin-tagged unmethylated H2AX peptide (Biotin-SATVGPKAPSGGKKATQASQEY) or biotin-tagged methylated H2AX peptide (Biotin-SATVGPKAPSGGK[me2K]ATQASQEY) is mixed with different amount of doxorubicin-induced HeLa cell lysates in the kinase buffer (50 mM HEPES, pH 7.5, 50 mM potassium chloride, 5 mM magnesium chloride, 10% glycerol, 1 mM ATP and 1 mM dithiothreitol) for 90 min at 30 °C. Peptides were precipitated with Dynabeads M-280 streptavidin (Life Technologies) and the samples were separated on SDS–PAGE using the Tris-Tricine Precast Gel (456-3063, Bio-Rad). Subsequently, WB was performed using an anti-γ-H2AX antibody (Millipore). The nitrocellulose membrane for the WB was stained with MemCode Reversible Stain (Thermo Scientific) to quantify the amount of peptides.
Dominant-negative experiments
HeLa cells were cultured for 24 h and transfected with a FLAG-H2AX-WT expression vector, a FLAG-H2AX-K134A expression vector or a FLAG-Mock (Empty) vector and an HA-SUV39H2 expression vector using FuGENE HD transfection reagent (Roche Applied Science). Cells were harvested 48 h after transfection and lysed with CelLytic M cell lysis reagent (Sigma-Aldrich) containing a complete protease inhibitor cocktail (Roche Applied Science). Samples were separated by standard SDS–PAGE and subsequently immunoblotted with anti-H2AK134me2, anti-γ-H2AX antibody, anti-H2AX, anti-FLAG and anti-α-Tubulin antibodies. Endogenous or exogenous H2AX K134 methylation and γ-H2AX levels were quantified by GS-800 calibrated imaging densitometer (Bio-Rad).
Radiosensitivity and chemosensitivity experiments
With regard to the knockdown experiment, we cultured RERF-LC-AI cells for 24 h and transfected control siRNA (siNC) and two independent SUV39H2 siRNAs into the cells. After 48 h, the cells were irradiated with 3 or 6 Gy of ionizing radiation by cabinet X-ray system (Newco), and cell viability was measured by CCK-8 (Dojindo) 24 h after irradiation. In addition, RERF-LC-AI cells were treated with various concentrations of cisplatin or doxorubicin 48 h after treatment with siRNAs, and the cells were cultured for additional 48 h. Subsequently, cell viability was measured by CCK-8 and the SigmaPlot software (Systat Software) was used to calculate IC50. As for the dominant-negative experiment, HeLa cells were transfected with a FLAG-H2AX-WT or a FLAG-H2AX-K134A expression vector with an HA-SUV39H2 expression vector using FuGENE HD (Roche Applied Science) reagent. After 24 and 72 h, transfected HeLa cells were irradiated with 6 or 10 Gy of ionizing radiation by cabinet X-ray system (Newco) and cell viability was measured by CCK-8 (Dojindo) 24 h after the second irradiation. In addition, vector-transfected HeLa cells were treated with various concentrations of cisplatin or doxorubicin 24 h after the transfection, and the cells were cultured for additional 48 h. Next, cell viability was measured by CCK-8. IC50 was calculated with the SigmaPlot software.
Colony formation assays
HeLa cells, cultured in EMEM with 10% FBS, were transfected with a p3xFLAG-H2AX-WT vector or a p3xFLAG-H2AX-K134A mutant vector, and an HA-SUV39H2 vector using FuGENE HD. Cells were irradiated with 0, 2, 4, 6 and 10 Gy of ionizing radiation by cabinet X-ray system 24 h after the transfection. Subsequently, the cells were cultures in the medium containing 0.8 mg ml−1 of Geneticin/G-418 for 15 days. The colony number was calculated by GS-800-calibrated densitometer and results are the means±s.d. of three independent experiments. P-values were calculated using Student’s t-test.
Statistical analysis
Each experiment has been repeated at least three times. Values were presented as the mean plus or minus s.d. Statistical analyses were performed using unpaired Student’s t-test (two groups) and Kruskal–Wallis one-way analysis of variance (more than two groups). Significant difference between groups was noted when P-value was <0.05.
Additional information
How to cite this article: Sone, K. et al. Critical role of lysine 134 methylation on histone H2AX for γ-H2AX production and DNA repair. Nat. Commun. 5:5691 doi: 10.1038/ncomms6691 (2014).
References
Moore, J. D. & Krebs, J. E. Histone modifications and DNA double-strand break repair. Biochem. Cell Biol. 82, 446–452 (2004).
Lowndes, N. F. & Toh, G. W. DNA repair: the importance of phosphorylating histone H2AX. Curr. Biol. 15, R99–R102 (2005).
Pinto, D. M. & Flaus, A. Structure and function of histone H2AX. Subcell. Biochem. 50, 55–78 (2010).
Bartova, E., Krejci, J., Harnicarova, A., Galiova, G. & Kozubek, S. Histone modifications and nuclear architecture: a review. J. Histochem. Biochem. 56, 711–721 (2008).
Bonner, W. M. et al. GammaH2AX and cancer. Nat. Rev. Cancer 8, 957–967 (2008).
Rogakou, E. P., Pilch, D. R., Orr, A. H., Ivanova, V. S. & Bonner, W. M. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J. Biol. Chem. 273, 5858–5868 (1998).
Harper, J. W. & Elledge, S. J. The DNA damage response: ten years after. Mol. Cell 28, 739–745 (2007).
Celeste, A. et al. Histone H2AX phosphorylation is dispensable for the initial recognition of DNA breaks. Nat. Cell Biol. 5, 675–679 (2003).
Celeste, A. et al. Genomic instability in mice lacking histone H2AX. Science 296, 922–927 (2002).
Messick, T. E. & Greenberg, R. A. The ubiquitin landscape at DNA double-strand breaks. J. Cell Biol. 187, 319–326 (2009).
Stewart, G. S. et al. The RIDDLE syndrome protein mediates a ubiquitin-dependent signaling cascade at sites of DNA damage. Cell 136, 420–434 (2009).
Mailand, N. et al. RNF8 ubiquitylates histones at DNA double-strand breaks and promotes assembly of repair proteins. Cell 131, 887–900 (2007).
Doil, C. et al. RNF168 binds and amplifies ubiquitin conjugates on damaged chromosomes to allow accumulation of repair proteins. Cell 136, 435–446 (2009).
Pan, M. R., Peng, G., Hung, W. C. & Lin, S. Y. Monoubiquitination of H2AX protein regulates DNA damage response signaling. J. Biol. Chem. 286, 28599–28607 (2011).
Bassing, C. H. et al. Increased ionizing radiation sensitivity and genomic instability in the absence of histone H2AX. Proc. Natl Acad. Sci. USA 99, 8173–8178 (2002).
Fernandez-Capetillo, O., Lee, A., Nussenzweig, M. & Nussenzweig, A. H2AX: the histone guardian of the genome. DNA Repair 3, 959–967 (2004).
McKinnon, P. J. & Caldecott, K. W. DNA strand break repair and human genetic disease. Annu. Rev. Genomics Hum. Genet. 8, 37–55 (2007).
Jeggo, P. A. & Lobrich, M. DNA double-strand breaks: their cellular and clinical impact? Oncogene 26, 7717–7719 (2007).
Allis, C. D. et al. New nomenclature for chromatin-modifying enzymes. Cell 131, 633–636 (2007).
O'Carroll, D. et al. Isolation and characterization of Suv39h2, a second histone H3 methyltransferase gene that displays testis-specific expression. Mol. Cell Biol. 20, 9423–9433 (2000).
Garcia-Cao, M., O’Sullivan, R., Peters, A. H., Jenuwein, T. & Blasco, M. A. Epigenetic regulation of telomere length in mammalian cells by the Suv39h1 and Suv39h2 histone methyltransferases. Nat. Genet. 36, 94–99 (2004).
Peters, A. H. et al. Loss of the Suv39h histone methyltransferases impairs mammalian heterochromatin and genome stability. Cell 107, 323–337 (2001).
Daury, L. & Trouche, D. Analysis of histone deposition on specific DNA elements in living mammalian cells. BioTechniques 35, 326–332 (2003).
Gertych, A., Oh, J. H., Wawrowsky, K. A., Weisenberger, D. J. & Tajbakhsh, J. 3-D DNA methylation phenotypes correlate with cytotoxicity levels in prostate and liver cancer cell models. BMC Pharmacol. Toxicol. 14, 11 (2013).
Ayrapetov, M. K., Gursoy-Yuzugullu, O., Xu, C., Xu, Y. & Price, B. D. DNA double-strand breaks promote methylation of histone H3 on lysine 9 and transient formation of repressive chromatin. Proc. Natl Acad. Sci. USA 111, 9169–9174 (2014).
Sun, Y. et al. Histone H3 methylation links DNA damage detection to activation of the tumour suppressor Tip60. Nat. Cell Biol. 11, 1376–1382 (2009).
Sedelnikova, O. A. & Bonner, W. M. GammaH2AX in cancer cells: a potential biomarker for cancer diagnostics, prediction and recurrence. Cell Cycle 5, 2909–2913 (2006).
Nagelkerke, A. et al. Constitutive expression of gamma-H2AX has prognostic relevance in triple negative breast cancer. Radiother. Oncol. 101, 39–45 (2011).
Brunner, A. H. et al. Expression of gamma-H2AX in endometrial carcinomas: an immunohistochemical study with p53. Gynecol. Oncol. 121, 206–211 (2011).
Matthaios, D. et al. gamma-H2AX expression detected by immunohistochemistry correlates with prognosis in early operable non-small cell lung cancer. OncoTargets Ther. 5, 309–314 (2012).
Jackson, J. P., Lindroth, A. M., Cao, X. & Jacobsen, S. E. Control of CpNpG DNA methylation by the KRYPTONITE histone H3 methyltransferase. Nature 416, 556–560 (2002).
Dickey, J. S. et al. H2AX: functional roles and potential applications. Chromosoma 118, 683–692 (2009).
Helleday, T., Petermann, E., Lundin, C., Hodgson, B. & Sharma, R. A. DNA repair pathways as targets for cancer therapy. Nat. Rev. Cancer 8, 193–204 (2008).
Hickson, I. et al. Identification and characterization of a novel and specific inhibitor of the ataxia-telangiectasia mutated kinase ATM. Cancer Res. 64, 9152–9159 (2004).
Kao, J. et al. gamma-H2AX as a therapeutic target for improving the efficacy of radiation therapy. Curr. Cancer Drug Targets 6, 197–205 (2006).
Redon, C. E. et al. Histone gammaH2AX and poly(ADP-ribose) as clinical pharmacodynamic biomarkers. Clin. Cancer Res. 16, 4532–4542 (2010).
Veuger, S. J., Curtin, N. J., Richardson, C. J., Smith, G. C. & Durkacz, B. W. Radiosensitization and DNA repair inhibition by the combined use of novel inhibitors of DNA-dependent protein kinase and poly(ADP-ribose) polymerase-1. Cancer Res. 63, 6008–6015 (2003).
Wang, D. & Lippard, S. J. Cellular processing of platinum anticancer drugs. Nat. Rev. Drug Discov. 4, 307–320 (2005).
Taneja, N. et al. Histone H2AX phosphorylation as a predictor of radiosensitivity and target for radiotherapy. J. Biol. Chem. 279, 2273–2280 (2004).
Kao, J., Lavaf, A., Lan, C. & Fu, S. Inhibition of gamma-H2AX after ionizing radiation as a biological surrogate of impaired upstream DNA damage signaling and radiosensitivity. J. Cancer Mol. 5, 49–54 (2010).
Reaper, P. M. et al. Selective killing of ATM- or p53-deficient cancer cells through inhibition of ATR. Nat. Chem. Biol. 7, 428–430 (2011).
Chaudhuri, L. et al. CHK1 and WEE1 inhibition combine synergistically to enhance therapeutic efficacy in acute myeloid leukemia ex vivo. Haematologica 99, 688–696 (2014).
Huntoon, C. J. et al. ATR inhibition broadly sensitizes ovarian cancer cells to chemotherapy independent of BRCA status. Cancer Res. 73, 3683–3691 (2013).
Prevo, R. et al. The novel ATR inhibitor VE-821 increases sensitivity of pancreatic cancer cells to radiation and chemotherapy. Cancer Biol. Ther. 13, 1072–1081 (2012).
Vavrova, J. et al. Inhibition of ATR kinase with the selective inhibitor VE-821 results in radiosensitization of cells of promyelocytic leukaemia (HL-60). Radiat. Environ. Biophys. 52, 471–479 (2013).
Hamamoto, R. et al. SMYD3 encodes a histone methyltransferase involved in the proliferation of cancer cells. Nat. Cell Biol. 6, 731–740 (2004).
Hamamoto, R. et al. Enhanced SMYD3 expression is essential for the growth of breast cancer cells. Cancer Sci. 97, 113–118 (2006).
Cho, H. S. et al. RB1 methylation by SMYD2 enhances cell cycle progression through an increase of RB1 phosphorylation. Neoplasia 14, 476–486 (2012).
Takawa, M. et al. Histone lysine methyltransferase SETD8 promotes carcinogenesis by deregulating PCNA expression. Cancer Res. 72, 3217–3227 (2012).
Cho, H. S. et al. Enhanced HSP70 lysine methylation promotes proliferation of cancer cells through activation of Aurora kinase B. Nat. Commun. 3, 1072 (2012).
Cho, H. S. et al. Demethylation of RB regulator MYPT1 by histone demethylase LSD1 promotes cell cycle progression in cancer cells. Cancer Res. 71, 1–6 (2011).
Takawa, M. et al. Validation of the histone methyltransferase EZH2 as a therapeutic target for various types of human cancer and as a prognostic marker. Cancer Sci. 102, 1298–1305 (2011).
Kogure, M. et al. Deregulation of the histone demethylase JMJD2A is involved in human carcinogenesis through regulation of the G/S transition. Cancer Lett. 336, 76–84 (2013).
Toyokawa, G. et al. Histone lysine methyltransferase Wolf-Hirschhorn Syndrome Candidate 1 is involved in human carcinogenesis through regulation of the Wnt Pathway. Neoplasia 13, 887–898 (2011).
Toyokawa, G. et al. The histone demethylase JMJD2B plays an essential role in human carcinogenesis through positive regulation of cyclin-dependent kinase 6. Cancer Prev. Res. (Phila.) 4, 2051–2061 (2011).
Toyokawa, G. et al. Minichromosome Maintenance Protein 7 is a potential therapeutic target in human cancer and a novel prognostic marker of non-small cell lung cancer. Mol. Cancer 10, 65 (2011).
Unoki, M. et al. UHRF1 is a novel molecular marker for diagnosis and the prognosis of bladder cancer. Br J Cancer 101, 98–105 (2009).
Cho, H. S. et al. Enhanced expression of EHMT2 is involved in the proliferation of cancer cells through negative regulation of SIAH1. Neoplasia 13, 676–684 (2011).
Hayami, S. et al. Overexpression of LSD1 contributes to human carcinogenesis through chromatin regulation in various cancers. Int. J. Cancer 128, 574–586 (2011).
Hayami, S. et al. Overexpression of the JmjC histone demethylase KDM5B in human carcinogenesis: involvement in the proliferation of cancer cells through the E2F/RB pathway. Mol. Cancer 9, 59 (2010).
Cho, H. S. et al. The JmjC domain-containing histone demethylase KDM3A is a positive regulator of the G1/S transition in cancer cells via transcriptional regulation of the HOXA1 gene. Int. J. Cancer 131, E179–E189 (2012).
Kang, D. et al. The histone methyltransferase Wolf-Hirschhorn syndrome candidate 1-like 1 (WHSC1L1) is involved in human carcinogenesis. Genes Chromosomes Cancer 52, 126–139 (2013).
Kogure, M. et al. The oncogenic polycomb histone methyltransferase EZH2 methylates lysine 120 on histone H2B and competes ubiquitination. Neoplasia 15, 1251–1261 (2013).
Yoshimatsu, M. et al. Dysregulation of PRMT1 and PRMT6, Type I arginine methyltransferases, is involved in various types of human cancers. Int. J. Cancer 128, 562–573 (2011).
Acknowledgements
We are grateful to Drs Yataro Daigo, Vassiliki Saloura, Gottfried Von Keudell, Deng Zhenzong, Kyohei Arita, Reem Ibrahim, Hyun-Soo Cho and Gouji Toyokawa for technical support and helpful discussion, to Dr Nicholas Shukeir for providing Suv39h2−/− MEF cells and to Dr Andre Nussenzweig for providing H2AX−/− MEF cells. We also thank the member of Nakamura laboratory in the University of Chicago for kind support.
Author information
Authors and Affiliations
Contributions
K.S., Y.N. and R.H. designed this study and performed all experiments with the help of L.P and M.N. K.U. performed the mass spectrometric analysis. T.J. provided WT and Suv39h2−/− MEF cells. K.S. and R.H. wrote this manuscript, and L.P., M.N., T.J. and Y.N. critically read the manuscript and gave valuable suggestions. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
Y. Nakamura is a stock holder and a scientific advisor of Oncotherapy Science, and also has research grants from Oncotherapy Science. All other authors declare no competing financial interests.
Supplementary information
Supplementary Information
Supplementary Figures 1-14, Supplementary Tables 1-6, and Supplementary Raw Data (PDF 1869 kb)
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Sone, K., Piao, L., Nakakido, M. et al. Critical role of lysine 134 methylation on histone H2AX for γ-H2AX production and DNA repair. Nat Commun 5, 5691 (2014). https://doi.org/10.1038/ncomms6691
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/ncomms6691
This article is cited by
-
Loss of H3K9 trimethylation alters chromosome compaction and transcription factor retention during mitosis
Nature Structural & Molecular Biology (2023)
-
Bisphenol A exposure decreases sperm production and male fertility through inhibition PCBP2 expression
Environmental Science and Pollution Research (2023)
-
Dosimetry of heavy ion exposure to human cells using nanoscopic imaging of double strand break repair protein clusters
Scientific Reports (2022)
-
Biological function and regulation of histone 4 lysine 20 methylation in DNA damage response
Genome Instability & Disease (2022)
-
Histone methyltransferase SUV39H2 regulates cell growth and chemosensitivity in glioma via regulation of hedgehog signaling
Cancer Cell International (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
