
Abstract
Reliably predicting how coral calcification may respond to ocean acidification and warming depends on our understanding of coral calcification mechanisms. However, the concentration and speciation of dissolved inorganic carbon (DIC) inside corals remain unclear, as only pH has been measured while a necessary second parameter to constrain carbonate chemistry has been missing. Here we report the first carbonate ion concentration ([CO32−]) measurements together with pH inside corals during the light period. We observe sharp increases in [CO32−] and pH from the gastric cavity to the calcifying fluid, confirming the existence of a proton (H+) pumping mechanism. We also show that corals can achieve a high aragonite saturation state (Ωarag) in the calcifying fluid by elevating pH while at the same time keeping [DIC] low. Such a mechanism may require less H+-pumping and energy for upregulating pH compared with the high [DIC] scenario and thus may allow corals to be more resistant to climate change related stressors.
Similar content being viewed by others

Introduction
Atmospheric CO2 has increased from 280 parts per million (ppm) during pre-industrial times to 400 ppm today1. This increase has led to invasion of CO2 into the surface ocean, which has shifted the acid–base equilibrium and modified the dissolved inorganic carbon (DIC) species composition such that the proton concentration ([H+]) has increased, and pH and [CO32−] have decreased; a process commonly known as ocean acidification (OA)2,3. Since OA reduces calcification rates in many, though not all, corals4,5,6,7,8, it is particularly detrimental to the health of coral reefs and their associated ecosystem functions and biogeochemical processes4,7,9. The decline in coral calcification rates is thought to be linked to reductions in seawater [CO32−] and thus Ωarag. However, because calcification occurs in a semi-isolated compartment where conditions are largely controlled by the coral10, the exact mechanism by which OA reduces calcification is difficult to decipher11. To understand and predict how corals might respond to current and future CO2 increases, climate warming and other stressors, we need a better understanding of coral calcification mechanisms, particularly how a high pH and Ωarag state is achieved inside the calcifying fluid and the sources as well as speciation and concentration of DIC that sustain calcification5,11,12,13,14,15,16.
In order to precipitate CaCO3 in a place separated from seawater by multiple layers of tissue, corals ultimately must be able to import Ca2+ ions and DIC from seawater and transport them to the site of calcification. These processes remain poorly understood and controversial. Although proposed routes of ion and molecule import differ in their details, potential import processes can be broadly grouped into three categories: (1) passive CO2 diffusion, (2) active bicarbonate and other ion transport and (3) whole seawater import though the time scale of each of these processes will differ10,16,17,18,19. Which of these three processes dominates the import pathway for DIC species remains unknown in part because a complete characterization of the internal carbonate chemistry of corals is lacking. In addition to the import of necessary ions, corals must raise the aragonite saturation state of the calcifying fluid to obtain rapid rates of calcification18. It is widely believed that this is achieved through a H+ efflux pump that creates high pH and, via chemical equilibrium and kinetic limitations, high [CO32−] in the calcifying space5,19. The H+ efflux pump is proposed to be coupled to Ca2+ influx via a Ca–H ATPase10. This theoretical model for coral calcification is considered the foundation for hypotheses explaining the response of calcification to future acidification5, yet direct measurements of at least two of the several carbonate system parameters to fully substantiate this conceptual model for coral calcification are lacking. Based on either indirect geochemical tracer evidence12 or sensor-based pH data combined with the assumption of either active HCO3− transport or whole seawater import13,20, several studies have also suggested that corals concentrate DIC to roughly twice the seawater value at the site of calcification. Others, however, have assumed that coral internal [DIC] is similar to or lower than that in seawater5,21. Furthermore, while a linkage between H+-pumping and carbon species transportation has recently been suggested12, confirmation by direct measurements is required. In summary, despite decades of research, our knowledge of the internal carbonate chemistry of corals is still limited and direct evidence is needed to definitively constrain coral calcification mechanisms.
Microelectrodes are a powerful tool for the direct measurement of chemical concentrations within organisms. The reliability and accuracy of pH and other microelectrodes have been well-established in the literature and in our work (see Methods section below). Previous pH microelectrode studies have shown that pH increases near the coral surface and in the upper region of the polyp’s gastric cavity due to photosynthesis, but decreases deeper in the cavity (coelenteron)22. Most importantly, a high pH has been measured in the calcifying fluid using both pH microelectrodes (pH: 9.3–10.1) and pH sensitive dyes (pH: 8.5–9.0) (refs 5, 13, 23). These experiments showed that corals increase the pH of the calcifying fluid to achieve a high Ωarag, which facilitates rapid rates of calcification. However, to fully characterize internal coral carbonate chemistry, pH alone is not sufficient and another carbonate system parameter such as [CO32−], [DIC], or total alkalinity (TA) must be assumed12,13 or preferably directly measured.
Here we use CO32− microelectrodes together with pH microelectrodes to directly measure two necessary system parameters and calculate concentrations of all other carbonate species within corals. Our novel findings are that [CO32−] and carbonate saturation state inside the coral calcifying fluid are high and calcifying fluid DIC concentration is low.
Results
pH and [CO32−] depth profiles
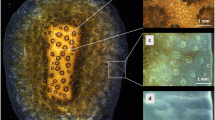
pH and CO32− microelectrodes were used to characterize the interior of coral polyps (see Methods section below) under ambient seawater pH and in the light (500 μmol photons m−2 s−1), when photosynthesis stimulates high rates of calcification. We advanced a microelectrode inside the polyp’s gastric cavity at a resolution of 50 μm per step (5 μm in one case) parallel to the polyp wall until it broke on the solid CaCO3 base (Fig. 1a,b). Repeated micro-profile measurements from seawater into the lower part of the coelenteron revealed that the profiles were reproducible (Fig. 1b, Supplementary Figs 1a,b and 2) even though polyp contraction sometimes affected the observed depths (Supplementary Fig. 2) and introduced noise in the electrode data.

(a) Close-up view of the coral T. reniformis showing the tip of a microelectrode as it enters the polyp. (b) Repeated pH microelectrode profile readings inside a T. reniformis coral polyp. Between the last point of the second insert and the next reading (which is a noise-free reading and is marked as ×), the microelectrode tip broke. pH microelectrodes were calibrated on the NBS scale, which is about 0.1–0.15 unit greater than that on the total scale. Error bars (s.d.) are defined in the Methods section. Positive depths are inside the coral polyp while negative depths are in the overlying seawater. Error bars are smaller than the symbols in the overlying water.
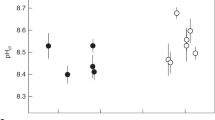
We obtained several full-depth profiles of pH and CO32− through the polyp mouth, the coelenteron, and into the calcifying fluid for three tropical coral species: Orbicella faveolata, Turbinaria reniformis and Acropora millepora. For O. faveolata, pH (in National Bureau of Standards, NBS, scale) increased from 8.1 and [CO32−] from 180 μmol kg−1 in the overlying seawater to 8.3–8.5 and ∼300 μmol kg−1, respectively, in the upper part of the polyp due to photosynthesis (Fig. 2a,b). Deeper in the polyps the pH decreased to a value of 7.7, which was consistent with earlier studies13,22,24. Correspondingly, in the deeper portion of the polyp, [CO32−] decreased to 100 μmol kg−1. Then, within a short distance, pH increased sharply to as high as 9.7, confirming previous work5,13,23, while [CO32−] increased six fold to 600 μmol kg−1 before the electrode tip broke. We interpret this rapid increase in pH and [CO32−] as evidence that the microelectrodes obtained measurements within the calcifying fluid prior to breaking.

(a) pH and (b) [CO32−] of Orbicella faveolata corals. (c) pH and (d) [CO32−] of Turbinaria reniformis and Acropora millepora corals. Dashed black lines indicate the top of the polyp mouths. Colored dashed lines roughly indicate the location of the calcifying fluid and differ in depth for each polyp.
Similar profiles were measured inside coral polyps of T. reniformis with highest pH in the calcifying fluid ranging from 8.8 to 9.3 (Fig. 2c) and highest [CO32−] from 1000 to 1400 μmol kg−1 (Fig. 2d). In A. millepora, the electrode advancement step was reduced to 5 μm due to the smaller polyps compared with the other species, resulting in a very fine-scale pH profile, which demonstrated a sharp increase in pH from 8.15 to 8.65 within only a short distance of 10–60 μm (Fig. 2c). Similarly, we observed a 15-fold increase in [CO32−] from ∼100 μmol kg−1 in the coelenteron to ∼1500 μmol kg−1 in the calcifying fluid within a distance of 50 μm in this species (Fig. 2d).
Composite depth profiles of CO2, DIC, TA and Ωarag
Our methods did not allow simultaneous profiling of pH and [CO32−] at the same spots due to both technical challenges and high heterogeneity inside a coral polyp. Therefore, we constructed composite profiles of pH and [CO32−] based on the range of our data across all three species and calculated [CO2], DIC, TA and Ωarag profiles (Supplementary Table 1), assuming the DIC system was in chemical equilibrium (see Methods section below). These composite profiles were then used to generate a more complete picture and a conceptual model of coral calcification.
Overall, our study shows that DIC decreases slightly near the polyp surface and in the upper part of the coelenteron due to photosynthetic CO2 consumption (Fig. 3a,b). Farther down in the polyp calculated DIC increases due to respiratory input of CO2 and reaches a peak at the bottom of the coelenteron before rapidly declining again in the calcifying space (Fig. 3b). Calculated TA in the coelenteron is nearly constant throughout most of the gastric cavity space, but increases dramatically in the calcifying fluid (Fig. 3b). Nonetheless, in agreement with direct TA measurements of the coelenteron fluid25 and some model assumptions5,21, DIC and TA values in the coelenteron solution are not substantially different from those of the overlying seawater.

(a) Illustrative model and (b) concentration distributions. Dark circles are photosynthetic endosymbionts. P stands for photosynthesis, R for respiration, and sw for seawater. In (b), for DIC (red line) and TA (purple line) the calculated ranges of possible concentrations inside the calcifying fluid are provided based on calculations of all species from Supplementary Table 1.
Notably, a sharp increase in [CO2] to ∼30 μmol kg−1 occurs toward the bottom of the coelenteron solution (Fig. 3b). This increase is not only due to an increase in DIC from respiration but is substantially enhanced by the nearly invariable TA (Fig. 3b, Supplementary Table 1). Thus, we attribute this [CO2] increase to both respiration input and the conversion of CO32− and HCO3− to CO2 by H+ exported from the calcifying fluid (Fig. 3a) – a mechanism suggested earlier as being important for Symbiodinium spp. photosynthetic needs19. Inside the calcifying fluid [CO2] is nearly zero (Fig. 3b), primarily due to the fluid’s very high pH (Fig. 2a,c). This very large [CO2] difference (∼30 μmol kg−1, Supplementary Table 1), based on direct measurements for the first time, confirms recent indirect evidence from geochemical tracers12 that a high rate of CO2 molecular diffusion from the coelenteron through the tissue into the calcifying fluid provides an important source of DIC for CaCO3 precipitation.
CO2 flux into the calcifying fluid
We estimated the CO2 influx to the calcifying fluid by both a 1-D diffusion model and an exchange model between the coelenteron and the calcifying fluid21,26 (equations and parameters are given in the Methods section and Supplementary Table 1). The CO2 diffusive flux was estimated to be as high as 217–433 nmol cm−2 h−1. Alternatively, the CO2 influx was estimated by the exchange model to be 82–301 nmol cm−2 h−1 (see Methods section). This range of CO2 influx and consumption rate compares favourably with coral calcification rates measured in the past (150–1000, nmol cm−2 h−1) (refs 10, 27). Based on the observations and model calculations presented here, we postulate that the coral’s proton pumping ability is essential not only for creating a high aragonite saturation state in the calcifying fluid (favouring calcification), but also for creating a sharp CO2 gradient facilitating DIC influx to support calcification. This CO2 transport mechanism is consistent with dual isotopic evidence18,28 and geochemical proxies12 showing that respired CO2 is an essential source of DIC for calcification. This passive CO2 transport mechanism may reduce the need for the active transport of HCO3− by processes such as transmembrane or vacuolar transport though the ultimate source of DIC for calcification comes from external seawater via photosynthesis, diffusive flux and whole seawater import.
Low [DIC] in the calcifying fluid
Despite the great CO2 flux into the coral calcifying fluid, our analysis reveals that the fluid [DIC] is not that different from seawater concentrations due to the consumption of DIC during calcification, which balances the CO2 influx (Fig. 3a). Even though TA in the calcifying fluid is elevated (1.2–1.9 times that of overlying seawater) it is not as high as previously reported values12. Allison et al.12 have recently used geochemical tracers to determine pH (from B isotope ratio) and estimate DIC (from B/Ca ratio) and concluded that in order to achieve a high aragonite saturation state, corals must internally concentrate DIC. Others also assumed that corals concentrate internal DIC to about twice, or more, the concentration of seawater13,29. Our direct pH and [CO32−] measurements show that while Ωarag inside the calcifying fluid is elevated, DIC is not elevated with respect to background seawater.
Discussion
Our microelectrode profiles of pH and CO32− through coral polyps provide the first direct evidence for elevated [CO32−] in the calcifying fluid. [CO32−] (600–1500 μmol kg−1) and corresponding aragonite mineral saturation states (Ωarag=8–22) in the calcifying fluid are high and consistent with expectations based on inorganic CaCO3 formation experiments11, previous experiments12 and model frameworks5,13.
Data from the calcifying fluid was only obtained in a small subset of the numerous profiles collected, most likely because the calcifying fluid layer is very thin (likely only a few micrometers) and existed in pockets13,16,17. In most cases, the electrode tips broke within one step (50 μm) between the bottom of the polyp gastric cavity space (coelenteron) and the hard CaCO3 skeleton. In some cases, however, the data suggest the space between the coelenteron and the skeleton could be thicker than previously thought (up to 100 μm). Alternatively, it is possible that the newly formed CaCO3 minerals are porous allowing the glass microelectrode tip to slightly penetrate the mineral surface, thus giving an inaccurate measure of the calcifying fluid thickness. It would be desirable to collect profiles with a better constraint of vertical and lateral heterogeneity of carbonate properties inside corals. Nonetheless a high pH, high [CO32−] layer was unequivocally detected right above the CaCO3 skeleton, though further work is needed to better define its thickness and assess its spatial variability. While the pH upregulation in the calcifying fluid measured by microelectrodes does overlap with those by the dye-based method13 and boron isotope method12, the highest values have been obtained from microelectrodes (9.7 here, 9.3 and 10.1 by two others, see Table 1)5,23. Differences between the approaches may result from differences in coral species, colony size30, measurement condition and location within the colony13,14, and time-scales over which the approaches integrate30.
If the very high pH of 9.0 or higher measured by microelectrodes reported here or elsewhere is typical of coral calcifying fluid, a high DIC internal solution (2 × of seawater DIC or higher) would indicate that internal Ωarag could be as high as 30 to 55 (Fig. 4a), which is inconsistent with Ωarag values inferred from abiotic experiments18. Therefore, it seems that the most reasonable internal [DIC] is about 1 × of seawater value, which is consistent with our results from the two direct carbonate parameter measurements. While further research is required to confirm our conclusion, our findings strongly shift the argument towards one of a lower calcifying fluid DIC concentration5,21.

(a) Ωarag and (b) buffer index. The buffer index is given as mmol kg−1 of H+ pumping (H+ loss or TA gain) per unit of pH upregulation. This index is calculated numerically with CO2SYS (ref. 37) (see Methods section). Here [DIC]=1800 μmol kg−1 is roughly the same as seawater value. pH is on the NBS scale.
One important difference between low versus high DIC scenarios is the necessary amount of protons that a coral must pump out (which results in an increase in TA) to raise its internal pH by one unit (in mmol-H+ per kg per pH unit). This buffer index for the high DIC scenario is at least twice that for the low DIC scenario, thus more protons must be removed in a high DIC scenario to achieve the observed calcifying fluid pH (Fig. 4b). Therefore, we conclude that the low DIC scenario would require less energy than the high DIC scenario as proton-pumping requires ATP. Comparisons of a relative change in energy requirement to upregulate internal pH between high external CO2 under future OA conditions and 400 ppm current conditions following the method of Ries5 also supported our conclusion (Supplementary Note 1 and Supplementary Table 5). As a result, if proton removal from the coelenteron is increasingly difficult as a result of higher seawater H+ concentrations under OA conditions31, the low DIC scenario should be advantageous as less H+-pumping will be required to further raise pH in the calcifying fluid.
Overall, this suggests that corals that use passive CO2 diffusion as an important pathway for DIC supply, and maintain low DIC in their calcifying fluid, may require less energy to calcify at high rates, which could potentially make them more resistant to OA and other stressors. However, at the same time, the lower buffering capacity of low DIC and moderate TA calcifying fluid means that the ability to maintain high fluid pH will be vulnerable to changes in the physiological and biochemical conditions that control H+-pumping. For example, a weakening of proton pumping during dark periods may lead a coral with low internal DIC towards decalcification more readily than otherwise expected in a high DIC scenario. In addition, a high internal DIC could allow corals to reach a high carbonate saturation state at lower internal pH (Fig. 4a)13. Therefore, additional studies using microelectrodes and other techniques in corals under OA conditions in combination with other stressors and environmental conditions (i.e., elevated temperature, nutrient levels and daytime versus nighttime) are a critical next step in elucidating coral calcification mechanisms under future ocean conditions.
Methods
Coral collection and preparation
The measurements focused on three species of corals: O. faveolata, T. reniformis and A. millepora. All species contain photosynthetic endosymbionts, often called zooxanthellae (Symbiodinium spp.), and form an aragonite skeleton. Small fragments of O. faveolata were sampled from the Florida Keys and transported to the laboratory in the University of Georgia (Florida Keys National Marine Sanctuary permit 2014–2015). Turbinaria reniformis and Acropora millepora coral fragments were provided by Reef Systems Coral Farm in New Albany, OH. The parent colonies of both species have been maintained in recirculating indoor aquaria with natural light (greenhouse, 700–1000 μmol photons m−2 s−1) and commercially available artificial seawater (Instant Ocean Reef Crystals) for over 10 years. Several fragments of each colony were shipped to the Cai lab at the University of Georgia, where they were acclimated to a closed-system laboratory aquarium with artificial seawater of salinity 35 and TA of 2.4 mmol kg−1 at 26 °C with light levels of about 200 μmol photons m−2 s−1 for at least two weeks prior to conducting pH and CO32− microelectrode profile measurements. TA in the aquarium was depleted at the time of microelectrode measurements (Supplementary Tables 2, 3 and 4) but was amended weekly.
Microelectrode construction and properties
pH microelectrodes32,33 and CO32− microelectrodes34,35 were constructed as previously described. We made our microelectrodes with a flat tip of diameter between 10 and 15 μm (Supplementary Fig. 3), which enabled us to make pH measurements along a profile into the coral polyp. pH microelectrodes were calibrated with three commercial pH NBS standards (pH=4, 7 and 10) at 26 °C to determine their response slopes. CO32− microelectrodes were calibrated in CO32− standard solutions pumped sequentially into a sealed chamber where an electrode was hosted35. Carbonate concentrations were set at approximately 50, 150, 250, 350, 500 and 1,000 μmol kg−1 by adding controlled amounts of HCl and NaHCO3 into filtered seawater collected from the Gulf of Mexico35. Standard solutions were then bubbled with room air for 24 h to minimize further exchange of CO2 with air during the experiment. Finally, CO32− concentrations were precisely calculated by CO2SYS with TA and pH measured by Gran titration and a commercial Ross glass electrode, respectively.
Data collection
In brief, a coral fragment was transferred from the incubation tank together with the tank water into a small measurement chamber that was kept at 26 °C and stirred with a magnetic stirrer (Supplementary Fig. 4). The coral fragment was allowed to acclimate in the small container for 1 h under 500 μmol photons m−2 s−1 until the microelectrode reading was stable in the chamber water (Fig. 1b & Supplementary Fig. 1). The slopes of pH and CO32− microelectrodes were calibrated immediately before use, and chamber water pH measured by a commercial electrode and measured TA values were used as the reference point before beginning the profiles. Measurement conditions are provided in Supplementary Tables 2,3 and 4. The point at which the microelectrode entered the polyp (defined as depth=0) was observed under a microscope. Typically the coral reacted by closing its mouth making this point unambiguous. The electrode was then advanced through the polyp’s gastric cavity in increments of 5–50 μm until the microelectrode broke on the skeleton. The microelectrode was held at each step until a stable reading was obtained. The pH reading was considered stable when drift was <0.2 mV or 0.003 pH min−1 (or <0.2 mV min−1 for CO32−) or a total of 3 min had passed, whichever was first. A 3-min limit was established because our pH microelectrodes had a much shorter response time32,35 but, in some cases, a stable reading was not realistic due to polyp movement.
Several replicate profiles were obtained to ensure that the microelectrodes were not fouled or otherwise damaged by profiling through the corals. In these tests a profile was obtained about half way through the coral coelenteron, at which point the electrode was returned to the overlying seawater, and the seawater readings were compared with values obtained before profiling was started. Then, the profile was resumed, repeating a portion of the earlier profile to ensure agreement between the two (Fig. 1b and Supplementary Figs 1 and 2). However, this practice cannot be extended into the calcifying fluid as an electrode is destined to be damaged and a repeating profile cannot be collected.
Finally, based on electrode calibration result, stability test and repeated measurements as shown above, we assign errors as ±0.02 pH unit for the overlying seawater, ±0.04 pH unit for the coelenteron and ±0.1 pH unit for the calcifying fluid. For [CO32−], a maximum uncertainty of ±10% is established based on our experiences and literature35,36.
Carbonate chemistry calculation
While the CO2SYS program37 is popularly used for carbonate equilibrium calculation based on two of the four parameters (TA, DIC, pH and pCO2), it does not accept the pH and [CO32−] pair as input parameters. However, measured pH and [CO32−] data allowed us to calculate concentrations of other DIC species, [DIC] and the TA in a straightforward way. For example,

where, K1 and K2 are the dissociation constants of the carbonic acid38. All equations are described in Supplementary Note 2. As in previous studies, we assumed that the coral’s internal salinity was the same as that of the external seawater salinity (that is corals are osmoconformers) for our thermodynamic calculation5,12,21. While a lower internal salinity would lead to a higher calculated DIC from the same observed pH and [CO32−], for example, calculated [DIC] is 0.8–12% higher at salinity=30 than that at 35, conclusion derived here holds well within a reasonable salinity range (see more details in Supplementary Note 3). In this work, pH in NBS scale is used. However, when needed, pH in total proton scale is converted to NBS scale roughly by adding 0.15 unit, pHNBS=pHT+0.15, according to CO2SYS simulation37 and Dickson39.
Although internal [Ca2+] was reported to be about 10% higher than that in seawater23, as it was the only reported value and as seawater [Ca2+] is very high (∼10.3 mM), we assumed that [Ca2+] in the calcifying fluid is the same as in seawater in calculating the saturation state Ω. Since [CO32−] is directly measured in this work, the uncertainty in Ω calculation is about the same as that in [Ca2+], roughly within 10%, and it does not affect the conclusions and discussion of this work.
Calculation of CO2 flux into the calcifying fluid
In a 1-D diffusion model, CO2 molecular diffusional flux is defined as:

where, D is the diffusion coefficient, and Δx is the distance corresponding to the concentration gradient. We assume D determined in seawater40 is applicable to the coral interior space. We calculated CO2 flux using the Δ[CO2] (Supplementary Table 1) and the corresponding distance between the two data points in the coelenteron and the calcifying fluid, respectively, as observed with our microelectrode (50–100 μm).
Using the calculated [CO2] gradient (29.8 μmol kg−1) from Supplementary Table 1, CO2 flux ( ) from the coelenteron into the calcifying fluid was calculated. Note the [CO2] gradient determined by the microelectrode data is not affected much by the uncertainty caused by relatively large pH and [CO32−] ranges inside the calcifying fluid as [CO2] is nearly 0 (that is an uncertainty of [CO2] between 0.5 and 0.1 μmol kg−1 only changes [CO2] gradient from 29.9 to 30.3 μmol kg−1) (Supplementary Table 1). In other words, a conservative internal pH value of 9.1 and a high value of 9.5 lead to similar estimates of CO2 gradient and flux.
) from the coelenteron into the calcifying fluid was calculated. Note the [CO2] gradient determined by the microelectrode data is not affected much by the uncertainty caused by relatively large pH and [CO32−] ranges inside the calcifying fluid as [CO2] is nearly 0 (that is an uncertainty of [CO2] between 0.5 and 0.1 μmol kg−1 only changes [CO2] gradient from 29.9 to 30.3 μmol kg−1) (Supplementary Table 1). In other words, a conservative internal pH value of 9.1 and a high value of 9.5 lead to similar estimates of CO2 gradient and flux.
Flux can also be calculated as the exchange rate between two adjacent ‘boxes’, the coelenteron and the calcifying fluid:

where, kco2 is the exchange coefficient (0.76–2.8 cm s−1) across biological tissues determined by an isotopic exchange method41, which has been used in two recent coral CO2 flux model simulations21,26. In principle, the two coefficients are linked by:

A lower CO2 flux calculated based on equation 3 than equation 2 may indicate that the CO2 diffusion coefficient across the coral tissue (or the coral tissue’s permeability) is lower than that of the seawater.
Amount of H+-pumping per unit pH upregulation
We define this parameter based on the buffer factor concept, discussed in details by Frankignoulle42, Hofmann et al.43 and Egleston et al.44. We take the buffer factor β defined below as a measure of how strong the calcifying fluid can buffer its pH:

We rearranged the definition to ΔTA/ΔpH=−2.3β, which represents the amount of proton pumping (or TA gain) a coral must do to upregulate its calcifying fluid pH by one unit. This was calculated numerically by following the same approach as the Revelle Factor calculation in the CO2SYS program37. We first set four DIC values and for each DIC value we allowed a range of pH changes. Thus, we derived TA values for the various DIC and pH combinations. Then, a ±1 μmol kg−1 disturbance is applied to TA values and the corresponding perturbation in pH is derived using the TA and DIC pairs as the input parameters in CO2SYS (Fig. 4b).
Energy consumption for pH upregulation
We calculated the relative energy changes of pH upregulation between different OA conditions as specified by Ries5 using the Nernst equation. The equations and results are given in the Supplementary Note 1 and Supplementary Table 5.
Additional information
How to cite this article: Cai, W-J. et al. Microelectrode characterization of coral daytime interior pH and carbonate chemistry. Nat. Commun. 7:11144 doi: 10.1038/ncomms11144 (2016).
References
IPCC. Climate Change 2013. The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (2014).
Caldeira, K. & Wickett, M. E. Anthropogenic carbon and ocean pH. Nature 425, 365–365 (2003).
Orr, J. C. et al. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 437, 681–686 (2005).
Hoegh-Guldberg, O. & Bruno, J. F. The impact of climate change on the world’s marine ecosystems. Science 328, 1523–1528 (2010).
Ries, J. B. A physicochemical framework for interpreting the biological calcification response to CO2-induced ocean acidification. Geochim. Cosmochim. Acta 75, 4053–4064 (2011).
Shamberger, K. E. F. et al. Diverse coral communities in naturally acidified waters of a Western Pacific reef. Geophys. Res. Lett. 41, 499–504 (2014).
Langdon, C. et al. Effect of elevated CO2 on the community metabolism of an experimental coral reef. Global Biogeochem. Cycles 17, 1011 (2003).
Schoepf, V. et al. Coral energy reserves and calcification in a high-CO2 world at two temperatures. PLoS ONE 8, e75049 (2013).
Doney, S. C. The growing human footprint on coastal and open-ocean biogeochemistry. Science 328, 1512–1516 (2010).
Allemand, D. et al. Biomineralisation in reef-building corals: from molecular mechanisms to environmental control. C. R. Palevol 3, 453–467 (2004).
Cohen, A. L. & Holcomb, M. Why corals care about ocean acidification: Uncovering the mechanism. Oceanography 22, 118–127 (2009).
Allison, N., Cohen, I., Finch, A. A., Erez, J. & Tudhope, A. W. Corals concentrate dissolved inorganic carbon to facilitate calcification. Nat. Commun. 5, 5741 (2014).
Venn, A., Tambutté, E., Holcomb, M., Allemand, D. & Tambutté, S. Live tissue imaging shows reef corals elevate pH under their calcifying tissue relative to seawater. PLoS ONE 6, e20013 (2011).
Holcomb, M. et al. Coral calcifying fluid pH dictates response to ocean acidification. Sci. Rep 4, 5207 (2014).
Gagnon, A. C. Coral calcification feels the acid. Proc. Natl Acad. Sci. 110, 1567–1568 (2013).
Tambutté, S. et al. Coral biomineralization: from the gene to the environment. J. Exp. Mar. Biol. Ecol. 408, 58–78 (2011).
Gagnon, A. C., Adkins, J. F. & Erez, J. Seawater transport during coral biomineralization. Earth Planet. Sci. Lett. 329–330, 150–161 (2012).
Cohen, A. L. & McConnaughey, T. A. Geochemical perspectives on coral mineralization. Rev. Mineral. Geochem. 54, 151–187 (2003).
McConnaughey, T. Zooxanthellae that open calcium channels: implications for reef corals. Mar. Ecol. Prog. Ser. 460, 277–287 (2012).
Zoccola, D. et al. Bicarbonate transporters in corals point towards a key step in the evolution of cnidarian calcification. Sci. Rep. 5, 9983 (2015).
Hohn, S. & Merico, A. Modelling coral polyp calcification in relation to ocean acidification. Biogeosciences 9, 4441–4454 (2012).
Kuhl, M., Cohen, Y., Dalsgaard, T., Jergensenl, B. & Revsbech, N. Microenvironment and photosynthesis of zooxanthellae in scleractinian corals studied with microsensors for O2, pH and light. Mar. Ecol. Prog. Ser. 117, 1995 (1995).
Al-Horani, F. A., Al-Moghrabi, S. M. & de Beer, D. The mechanism of calcification and its relation to photosynthesis and respiration in the scleractinian coral Galaxea fascicularis. Mar. Biol. 142, 419–426 (2003).
Barott, K. L., Venn, A. A., Perez, S. O., Tambutté, S. & Tresguerres, M. Coral host cells acidify symbiotic algal microenvironment to promote photosynthesis. Proc. Natl Acad. Sci. 112, 607–612 (2015).
Agostini, S. et al. Biological and chemical characteristics of the coral gastric cavity. Coral Reefs 31, 147–156 (2012).
Nakamura, T., Nadaoka, K. & Watanabe, A. A coral polyp model of photosynthesis, respiration and calcification incorporating a transcellular ion transport mechanism. Coral Reefs 32, 779–794 (2013).
Barnes, D. J. & Devereux, M. J. Productivity and calcification on a coral reef: A survey using pH and oxygen electrode techniques. J. Exp. Mar. Biol. Ecol. 79, 213–231 (1984).
Furla, P., Galgani, I., Durand, I. & Allemand, D. Sources and mechanisms of inorganic carbon transport for coral calcification and photosynthesis. J. Exp. Biol. 203, 3445–3457 (2000).
McCulloch, M., Falter, J., Trotter, J. & Montagna, P. Coral resilience to ocean acidification and global warming through pH up-regulation. Nat. Clim. Change 2, 623–627 (2012).
Honisch, B. et al. Assessing scleractinian corals as recorders for paleo-pH: Empirical calibration and vital effects. Geochim. Cosmochim. Acta 68, 3675–3685 (2004).
Jokiel, P. L. Ocean acidification and control of reef coral calcification by boundary layer limitation of proton flux. Bull. Mar. Sci. 87, 639–657 (2011).
Zhao, P. & Cai, W.-J. pH polymeric membrane microelectrodes based on neutral carriers and their application in aquatic environments. Anal. Chim Acta 395, 285–291 (1999).
Cai, W.-J., Zhao, P. & Wang, Y. pH and pCO2 microelectrodes measurement and diffusive behavior of carbon dioxide species in coastal marine sediments. Mar. Chem. 70, 133–148 (2000).
de Beer, D. et al. A microsensor for carbonate ions suitable for microprofiling in freshwater and saline environments. Limnol. Oceanogr. Meth. 6, 532–541 (2008).
Han, C., Cai, W.-J., Wang, Y. & Ye, Y. Calibration and evaluation of a carbonate microsensor for studies of the marine inorganic carbon system. J. Oceanogr. 70, 425–433 (2014).
Lee, B. H., Shim, Y.-B. & Park, S. B. A lipophilic sol−gel matrix for the development of a carbonate-selective electrode. Anal. Chem. 76, 6150–6155 (2004).
Pierrot, D., Lewis, E. & Wallace, D. W. R. MS Excel Program Developed for CO2 System Calculations Oak Ridge National Laboratory (2006).
Millero, F. J., Graham, T. B., Huang, F., Bustos-Serrano, H. & Pierrot, D. Dissociation constants of carbonic acid in seawater as a function of salinity and temperature. Mar. Chem. 100, 80–94 (2006).
Dickson, A. G. pH buffers for sea water media based on the total hydrogen ion concentration scale. Deep-Sea Res. 40, 107–118 (1993).
Li, Y. H. & Gregory, S. Diffusion of ions in sea water and in deep-sea sediments. Geochim. Cosmochim. Acta 38, 703 (1974).
Sültemeyer, D. & Rinast, K.-A. The CO2 permeability of the plasma membrane of Chlamydomonas reinhardtii: mass-spectrometric 18O-exchange measurements from 13C18O2 in suspensions of carbonic anhydrase-loaded plasma-membrane vesicles. Planta 200, 358–368 (1996).
Frankignoulle, M. A complete set of buffer factors for acid-base CO2 system in seawater. J. Mar. Syst. 5, 111–118 (1994).
Hofmann, A., Soetaert, K., Middelburg, J. & Meysman, F. R. AquaEnv: an aquatic acid–base modelling environment in R. Aquat. Geochem. 16, 507–546 (2010).
Egleston, E. S., Sabine, C. L. & Morel, F. M. M. Revelle revisited: Buffer factors that quantify the response of ocean chemistry to changes in DIC and alkalinity. Global Biogeochem. Cycles 24, GB1002 (2010).
Acknowledgements
The authors acknowledge financial supports from the NSF EF-1041124 to A.G., 1040940 to M.W., 1041070 to W-J.C., and EF-1315944 to B.H. & W-J.C. D.Q. and X.Y. acknowledge the supports of China Scholarship Council for supporting their visits to the Cai laboratory at the University of Georgia.
Author information
Authors and Affiliations
Contributions
W-J.C., A.G., M.W., B.H. and Y.W. conceived and designed the experiments; Y.M., D.Q. and Y.W. performed the experiments; Y.Y., C.H., H.X., B.H. and Y.W. contributed materials/analysis tools; W-J.C. and Y.M. analysed the data, and H.X. and W-J.C. performed the buffer index calculation. W-J.C. prepared the manuscript. All authors contributed to discussion and writing of the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Information
Supplementary Figures 1-4, Supplementary Tables 1-5, Supplementary Notes 1-3 and Supplementary References (PDF 276 kb)
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Cai, WJ., Ma, Y., Hopkinson, B. et al. Microelectrode characterization of coral daytime interior pH and carbonate chemistry. Nat Commun 7, 11144 (2016). https://doi.org/10.1038/ncomms11144
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/ncomms11144
This article is cited by
-
Crystallographic and chemical signatures in coral skeletal aragonite
Coral Reefs (2022)
-
Recent ocean acidification trends from boron isotope (δ11B) records of coral: Role of oceanographic processes and anthropogenic CO2 forcing
Journal of Earth System Science (2022)
-
Intracellular pH regulation: characterization and functional investigation of H+ transporters in Stylophora pistillata
BMC Molecular and Cell Biology (2021)
-
Heat stress differentially impacts key calcification mechanisms in reef-building corals
Coral Reefs (2021)
-
Moderate nutrient concentrations are not detrimental to corals under future ocean conditions
Marine Biology (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
