
Abstract
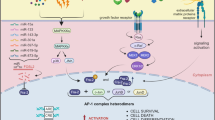
The transcription factor AP-1 (activator protein-1) is involved in cellular proliferation, transformation and death. Using mice and cells lacking AP-1 components, the target-genes and molecular mechanisms mediating these processes were recently identified. Interestingly, the growth-promoting activity of c-Jun is mediated by repression of tumour suppressors, as well as upregulation of positive cell cycle regulators. Mostly, c-Jun is a positive regulator of cell proliferation, whereas JunB has the converse effect. The intricate relationships between the different Jun proteins, their activities and the mechanisms that mediate them will be discussed.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout


Similar content being viewed by others


References
Angel, P. & Karin, M. The role of Jun, Fos and the AP-1 complex in cell-proliferation and transformation. Biochem. Biophys. Acta 1072, 129–157 (1991).
Chimenov, Y. & Kerppola, T. K. Close encounters of many kinds: Fos–Jun interactions that mediate transcription regulatory specificity. Oncogene 6, 533–542 (2001).
Ryseck, R. P. & Bravo, R. cJun, JunB, and JunD differ in their binding affinities to the AP-1 and CRE consensus sequences: effect of Fos proteins. Oncogene 6, 533–542 (1991).
Schutte, J. et al. JunB inhibits and cFos stimulates the transforming and transactivating activities of cJun. Cell 59, 987–997 (1989).
Chiu, R., Angel, P. & Karin, M. JunB differs in its biological properties from, and is a negative regulator of c-Jun. Cell 59, 979–986 (1989).
Karin, M. The regulation of AP-1 activity by mitogen-activated protein kinases. J. Biol. Chem. 270, 16483–16486 (1995).
Karin, M., Liu, Z.-G. & Zandi, E. AP-1 function and regulation. Curr. Opin. Cell Biol. 9, 240–246 (1997).
Schreiber, M. et al. Control of cell cycle progression by c-Jun is p53 dependent. Genes Dev. 13, 607–619 (1999).
Passegue, E. & Wagner, E. F. JunB suppresses cell proliferation by transcriptional activation of p16(INK4a) expression. EMBO J. 19, 2969–2979 (2000).
Shaulian, E. et al. The mammalian UV response: c-Jun induction is required for exit from p53-imposed growth arrest. Cell 103, 897–907 (2000).
Pessah, M. et al. c-Jun interacts with the corepressor TG-interacting factor (TGIF) to suppress Smad2 transcriptional activity. Proc. Natl Acad. Sci. USA 98, 6198–6120 (2001).
Hsu, J. C., Cressman, D. E. & Taub, R. Promoter-specific trans-activation and inhibition mediated by JunB. Cancer Res. 53, 3789–3794 (1993).
Chang, L. & Karin, M. Mammalian MAP kinase signalling cascades. Nature 410, 37–40 (2001).
Hill, C. S., Wynne, J. & Treisman, R. Serum-regulated transcription by serum response factor (Srf) – a novel role for the DNA binding domain. EMBO J. 13, 5421–5432 (1994).
Gruda, M. C., Kovary, K., Metz, R. & Bravo, R. Regulation of Fra-1 and Fra-2 phosphorylation differs during the cell cycle of fibroblasts and phosphorylation in vitro by MAP kinase affects DNA binding activity. Oncogene 9, 2537–2547 (1994).
Kato, Y. et al. BMK1/ERK5 regulates serum-induced early gene expression through transcription factor MEF2C. EMBO J. 16, 7054–7066 (1997).
Cavigelli, M., Dolfi, F., Claret, F. X. & Karin, M. Induction of c-fos expression through JNK-mediated TCF/Elk-1 phosphorylation. EMBO J. 14, 5957–5964 (1995).
Gupta, S., Campbell, D., Dérijard, B. & Davis, R. J. Transcription factor ATF2: regulation by the JNK signal transduction pathway. Science 267, 389–393 (1995).
Devary, Y., Gottlieb, R. A., Lau, L. & Karin, M. Rapid and preferential activation of the c-jun gene during the mammalian UV response. Mol. Cell. Biol. 11, 2804–2811 (1991).
Schonthal, A., Herrlich, P., Rahmsdorf, H. J. & Ponta, H. Requirement for fos gene expression in the transcriptional activation of collagenase by other oncogenes and phorbol esters. Cell 54, 325–334 (1988).
Hibi, M., Lin, A., Smeal, T., Minden, A. & Karin, M. Identification of an oncoprotein-responsive and UV-responsive protein kinase that binds and potentiates the c-Jun activation domain. Genes Dev. 7, 2135–2148 (1993).
Dérijard, B. et al. JNK1: A protein kinase stimulated by UV light and Ha-Ras that binds and phosphorylates the c-Jun activation domain. Cell 76, 1025–1037 (1994).
Han, J., Jiang, Y., Li, Z., Kravchenko, V. V. & Ulevitch, R. J. Activation of the transcription factor MEF2C by the MAP kinase p38 in inflammation. Nature 386, 296–299 (1997).
Shaulian, E. & Karin, M. AP-1 in cell proliferation and survival. Oncogene 20, 2390–2400 (2001).
Kovary, K. & Bravo, R. The jun and fos protein families are both required for cell cycle progression in fibroblasts. Mol. Cell. Biol. 11, 4466–4472 (1991).
Brown, J. R. et al. Fos family members induce cell cycle entry by activating cyclin D1. Mol. Cell Biol. 18, 5609–5619 (1998).
Brusselbach, S. et al. Cell proliferation and cell cycle progression are not impaired in fibroblasts and ES cells lacking c-Fos. Oncogene 10, 79–86 (1995).
Thepot, D. et al. Targeted disruption of the murine junD gene results in multiple defects in male reproductive function. Development 127, 143–153 (2000).
Behrens, A., Sibilia, M. & Wagner, E. F. Amino-terminal phosphorylation of c-Jun regulates stress-induced apoptosis and cellular proliferation. Nature Genet. 21, 326–329 (1999).
Johnson, R. S., van Lingen, B., Papaioannou, V. E. & Spiegelman, B. W. A null mutation at the c-jun locus causes embryonic lethality and retarded cell growth in culture. Genes Dev. 7, 1309–1317 (1993).
Eferl, R. et al. Functions of c-jun in liver and heart development. J. Cell Biol. 145, 1049–1061 (1999).
Szabowski, A. et al. c-Jun and JunB antagonistically control cytokine-regulated mesenchymal–epidermal interaction in skin. Cell 103, 745–755 (2000).
Wisdom, R., Johnson, R. S. & Moore, C. c-Jun regulates cell cycle progression and apoptosis by distinct mechanisms. EMBO J. 18, 188–197 (1999).
Xiong, Y. et al. p21 is a universal inhibitor of cyclin kinases. Nature 366, 701–704 (1993).
El-Deiry, W. S. et al. WAF1, a potential mediator of p53 tumor suppression. Cell 75, 817–825 (1993).
Clark, W. et al. v-Jun overrides the mitogen dependence of S-phase entry by deregulating retinoblastoma protein phosphorylation and E2F-pocket protein interactions as a consequence of enhanced cyclin E–cdk2 catalytic activity. Mol. Cell Biol. 20, 2529–2542 (2000).
Hennigan, R. F. & Stambrook, P. J. Dominant negative c-jun inhibits activation of the cyclin D1 and cyclin E kinase complexes. Mol. Biol. Cell 12, 2352–2363 (2001).
Schutte, J., Minna, J. D. & Birrer, M. Deregulated expression of human {Ic-jun} transforms primary rat embryo cells in cooperation with and activated c-Ha-{Iras} gene and transforms Rat-la cells as a single gene. Proc. Natl Acad. Sci. USA 86, 2257–2261 (1989).
Schorpp-Kistner, M., Wang, Z. Q., Angel, P. & Wagner, E. F. JunB is essential for mammalian placentation. EMBO J. 18, 934–948 (1999).
Bakiri, L., Lallemand, D., Bossy-Wetzel, E. & Yaniv, M. Cell cycle-dependent variations in c-Jun and JunB phosphorylation: a role in the control of cyclin D1 expression. EMBO J. 19, 2056–2068 (2000).
Passegue, E., Jochum, W., Schorpp-Kistner, M., Mohle-Steinlein, U. & Wagner, E. F. Chronic myeloid leukemia with increased granulocyte progenitors in mice lacking junB expression in the myeloid lineage. Cell 104, 21–32 (2001).
Passegue, E., Jochum, W., Behrens, A., Ricci, R. & Wagner, E. JunB can substitute for c-Jun in mouse development and cell proliferation. Nature Genet. 30, 158–166 (2002).
Weitzman, J. B., Fiette, L., Matsuo, K. & Yaniv, M. JunD protects cells from p53-dependent senescence and apoptosis. Mol. Cell 6, 1109–1119 (2000).
Sherr, C. J. Tumor surveillance via the ARF-p53 pathway. Genes Dev. 12, 2984–2991 (1998).
Agarwal, S. K. et al. Menin interacts with the AP1 transcription factor JunD and represses JunD-activated transcription. Cell 96, 143–152 (1999).
Pfarr, C. M. et al. Mouse JunD negatively regulates fibroblast growth and antagonizes transformation by ras. Cell 76, 747–760 (1994).
Maki, Y., Bos, T. J., Davis, C., Starbuck, M. & Vogt, P. K. Avian sarcoma cirus 17 carries the Jun oncogene. Proc. Natl Acad. Sci. USA 84, 2848–2852 (1987).
van Straaten, F., Muller, R., Curran, T., Van Beveren, C. & Verma, I. M. Complete nucleotide sequence of a human c-onc gene: deduced amino acid sequence of the human c-fos protein. Proc. Natl Acad. Sci. USA 80, 3183–3187 (1983).
Vandel, L. et al. Stepwise transformation of rat embryo fibroblasts: c-Jun, JunB, or JunD can cooperate with Ras for focus formation, but a c-Jun-containing heterodimer is required for immortalization. Mol. Cell. Biol. 16, 1881–1888 (1996).
Johnson, R., Spiegelman, B., Hanahan, D. & Wisdom, R. Cellular transformation and malignancy induced by ras require c-jun. Mol. Cell. Biol. 16, 4504–4511 (1996).
Behrens, A., Jochum, W., Sibilia, M. & Wagner, E. F. Oncogenic transformation by ras and fos is mediated by c-Jun N-terminal phosphorylation. Oncogene 19, 2657–2663 (2000).
Hartl, M., Reiter, F., Bader, A. G., Castellazzi, M. & Bister, K. JAC, a direct target of oncogenic transcription factor Jun, is involved in cell transformation and tumorigenesis. Proc. Natl Acad. Sci. USA 98, 13601–13606 (2001).
Fu, S., Bottoli, I., Goller, M. & Vogt, P. K. Heparin-binding epidermal growth factor-like growth factor, a v-Jun target gene, induces oncogenic transformation. Proc. Natl Acad. Sci. USA 96, 5716–5721 (1999).
Miller, A., Curran, T. & Verma, I. cFos can induce cellular transformation: novel mechanism of activating a cellular oncogene. Cell 36, 51–60 (1984).
Mechta, F., Lallemand, D., Pfarr, C. M. & Yaniv, M. Transformation by ras modifies AP1 composition and activity. Oncogene 14, 837–847 (1997).
Bakin, A. V. & Curran, T. Role of DNA 5-methylcytosine transferase in cell transformation by fos. Science 283, 387–390 (1999).
Toft, D. J., Rosenberg, S. B., Bergers, G., Volpert, O. & Linzer, D. I. Reactivation of proliferin gene expression is associated with increased angiogenesis in a cell culture model of fibrosarcoma tumor progression. Proc. Natl Acad. Sci. USA 98, 13055–13059 (2001).
van Dam, H. et al. Autocrine growth and anchorage independence: two complementing Jun-controlled genetic programs of cellular transformation. Genes Dev. 12, 1227–1239 (1998).
Ruther, U., Garber, C., Komitowski, D., Muller, R. & Wagner, E. F. Deregulated c-fos expression interferes with normal bone development in transgenic mice. Nature 325, 412–416 (1987).
Ruther, U., Komitowski, D., Schubert, F. R. & Wagner, E. F. c-fos expression induces bone tumors in transgenic mice. Oncogene 4, 861–865 (1989).
Saez, E. et al. c-fos is required for malignant progression of skin tumors. Cell 82, 721–732 (1999).
Young, M. R. et al. Transgenic mice demonstrate AP-1 (activator protein-1) transactivation is required for tumor promotion. Proc. Natl Acad. Sci. USA 96, 9827–9832 (1999).
Jochum, W., Passegue, E. & Wagner, E. F. AP-1 in mouse development and tumorigenesis. Oncogene 20, 2401–2412 (2001).
Wulf, G. M. et al. EMBO J. 20, 3459–3472 (2001).
Smeyne, R. Continuous c-Fos expression procedes programmed cell death in vivo. Nature 363, 166–169 (1993).
Marti, A. et al. Protein kinase A and AP-1 (c-Fos/JunD) are induced during apoptosis of mouse mammary epithelial cells. Oncogene 9, 1213–1223 (1994).
Buttyan, R., Zakeri, Z., Lockshin, R. & Wolgemuth, D. Cascade induction of c-fos, c-myc, and heat shock 70K transcripts during regression of the rat ventral prostate gland. Mol. Endocrinol. 2, 650–657 (1988).
Estus, S., et al. Altered gene expression in neurons during programmed cell death: identification of c-jun as necessary for neuronal apoptosis. J. Cell Biol. 127, 1717–1727 (1994).
Ham, J. et al. A c-Jun dominant negative mutant protects sympathetic neurons against programmed cell death. Neuron 14, 927–939 (1995).
Colotta, F., Polentarutti, N., Sironi, M. & Mantovani, A. Expression and involvement of c-fos and c-jun protooncogenes in programmed cell death induced by growth factor deprivation in lymphoid cell lines. J. Biol. Chem. 267, 18278–18283 (1992).
Karin, M. Mitogen-activated protein kinase cascades as regulators of stress responses. Ann. NY Acad. Sci. 851, 139–146 (1998).
Bossy-Wetzel, E., Bakiri, L. & Yaniv, M. Induction of apoptosis by the transcription factor c-Jun. EMBO J. 16, 1695–1709 (1997).
Preston, G. A. et al. Induction of apoptosis by c-Fos protein. Mol. Cell Biol. 16, 211–218 (1996).
Le-Niculescu, H. et al. Withdrawal of survival factors results in activation of the JNK pathway in neuronal cells leading to Fas ligand induction and cell death. Mol. Cell. Biol. 19, 751–763 (1999).
Xia, Z., Dickens, M., Raingeaud, J., Davis, R. J. & Greenberg, M. E. Opposing effects of ERK and JNK-p38 MAP kinases on apoptosis. Science 270, 1326–1331 (1995).
J., W., Neame, S. J., Paquet, L., Bernard, O. & Ham, J. Dominant-negative c-Jun promotes neuronal survival by reducing BIM expression and inhibiting mitochondrial cytochrome c release. Neuron 29, 629–643 (2001).
Hafezi, F. et al. The absence of c-fos prevents light-induced apoptotic cell death of photoreceptors in retinal degeneration in vivo. Nature Med. 3, 346–349 (1997).
Whitfield, J., Neame, S. J., Paquet, L., Bernard, O. & Ham, J. Dominant-negative c-Jun promotes neuronal survival by reducing BIM expression and inhibiting mitochondrial cytochrome c release. Neuron 29, 629–643 (2001).
Yang, D. et al. Absence of excitotoxicity-induced apoptosis in the hippocampus of mice lacking the Jnk3 gene. Nature 389, 865–870 (1997).
De Smaele, E. et al. Induction of gadd45β by NF-κB downregulates pro-apoptotic JNK signalling. Nature 414, 308–313 (2001).
Tang, G. et al. Inhibition of JNK activation through NF-κB target genes. Nature 414, 313–317 (2001).
Xu, X., Raber, J., Yang, D., Su, B. & Mucke, L. Dynamic regulation of c-Jun N-terminal kinase activity in mouse brain by environmental stimuli. Proc. Natl Acad. Sci. USA 94, 12655–12660 (1997).
Kenney, A. M. & Kocsis, J. D. Peripheral axotomy induces long-term c-Jun amino-terminal kinase-1 activation and activator protein-1 binding activity by c-Jun and junD in adult rat dorsal root ganglia in vivo. J. Neurosci. 18, 1318–1328 (1998).
Herdegen, T. et al. Lasting N-terminal phosphorylation of c-Jun and activation of c-Jun N-terminal kinases after neuronal injury. J. Neurosci. 18, 5124–5135 (1998).
Tournier, C. et al. Requirement of JNK for stress-induced activation of the cytochrome c-mediated death pathway. Science 288, 870–874 (2000).
Kolbus, A. et al. c-Jun-dependent CD95-L expression is a rate-limiting step in the induction of apoptosis by alkylating agents. Mol. Cell. Biol. 20, 575–582 (2000).
Verheij, M. et al. Requirement for ceramide-initiated SAPK/JNK signalling in stress-induced apoptosis. Nature 380, 75–79 (1996).
Ivanov, V. N. et al. Cooperation between STAT3 and c-jun suppresses Fas transcription. Mol. Cell 7, 517–528 (2001).
Owen-Schaub, L. B. et al. Wild-type human p53 and a temperature-sensitive mutant induce Fas/APO-1 expression. Mol. Cell. Biol. 15, 3032–3049 (1995).
Smith, M. L. & Fornace, A. J. Jr. p53-mediated protective responses to UV irradiation. Proc. Natl Acad. Sci. USA 94, 12255–12257 (1997).
Bissonnette, N. & Hunting, D. J. p21-induced cycle arrest in G(1) protects cells from apoptosis induced by UV-irradiation or RNA polymerase II blockage. Oncogene 16, 3461–3469 (1998).
Gniadecki, R., Hansen, M. & Wulf, H. C. Resistance of senescent keratinocytes to UV-induced apoptosis. Cell. Mol. Biol. 46, 121–127 (2000).
Hilberg, F., Aguzzi, A., Howells, N. & Wagner, E. F. c-jun is essential for normal mouse development and hepatogenesis. Nature 365, 179–181 (1993).
Kuan, C. Y. et al. The Jnk1 and Jnk2 protein kinases are required for regional specific apoptosis during early brain development. Neuron 22, 667–676 (1999).
Liu, Z.-G., Hu, H., Goeddel, D. V. & Karin, M. Dissection of TNF receptor 1 effector functions: JNK activation is not linked to apoptosis, while NF-κB activation prevents cell death. Cell 87, 565–576 (1996).
Natoli, G. et al. Activation of SAPK/JNK by TNF receptor 1 through a noncytotoxic TRAF2-dependent pathway. Science 275, 200–203 (1997).
Nishina, H. et al. Stress-signaling kinase Sek1 protects thymocytes from apoptosis mediated by CD95 and CD3. Nature 385, 350–353 (1997).
Rebollo, A. et al. Bcl-3 expression promotes cell survival following interleukin-4 deprivation and is controlled by AP1 and AP1-like transcription factors. Mol. Cell. Biol. 20, 3407–3416 (2000).
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Shaulian, E., Karin, M. AP-1 as a regulator of cell life and death. Nat Cell Biol 4, E131–E136 (2002). https://doi.org/10.1038/ncb0502-e131
Issue Date:
DOI: https://doi.org/10.1038/ncb0502-e131
This article is cited by
-
The siRNA-mediated knockdown of AP-1 restores the function of the pulmonary artery and the right ventricle by reducing perivascular and interstitial fibrosis and key molecular players in cardiopulmonary disease
Journal of Translational Medicine (2024)
-
Role of Fra-2 in cancer
Cell Death & Differentiation (2024)
-
Regulatory mechanisms and therapeutic potential of JAB1 in neurological development and disorders
Molecular Medicine (2023)
-
Novel, thalidomide-like, non-cereblon binding drug tetrafluorobornylphthalimide mitigates inflammation and brain injury
Journal of Biomedical Science (2023)
-
The role of c-Jun for beating cardiomycyte formation in prepared embryonic body
Stem Cell Research & Therapy (2023)
