
Abstract
Macroautophagy (hereafter referred to as autophagy) degrades various intracellular constituents to regulate a wide range of cellular functions, and is also closely linked to several human diseases1,2. In selective autophagy, receptor proteins recognize degradation targets and direct their sequestration by double-membrane vesicles called autophagosomes, which transport them into lysosomes or vacuoles3. Although recent studies have shown that selective autophagy is involved in quality/quantity control of some organelles, including mitochondria and peroxisomes4, it remains unclear how extensively it contributes to cellular organelle homeostasis. Here we describe selective autophagy of the endoplasmic reticulum (ER) and nucleus in the yeast Saccharomyces cerevisiae. We identify two novel proteins, Atg39 and Atg40, as receptors specific to these pathways. Atg39 localizes to the perinuclear ER (or the nuclear envelope) and induces autophagic sequestration of part of the nucleus. Atg40 is enriched in the cortical and cytoplasmic ER, and loads these ER subdomains into autophagosomes. Atg39-dependent autophagy of the perinuclear ER/nucleus is required for cell survival under nitrogen-deprivation conditions. Atg40 is probably the functional counterpart of FAM134B, an autophagy receptor for the ER in mammals that has been implicated in sensory neuropathy5. Our results provide fundamental insight into the pathophysiological roles and mechanisms of ‘ER-phagy’ and ‘nucleophagy’ in other organisms.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




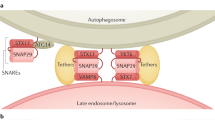
Similar content being viewed by others


References
Nakatogawa, H., Suzuki, K., Kamada, Y. & Ohsumi, Y. Dynamics and diversity in autophagy mechanisms: lessons from yeast. Nature Rev. Mol. Cell Biol. 10, 458–467 (2009)
Schneider, J. L. & Cuervo, A. M. Autophagy and human disease: emerging themes. Curr. Opin. Genet. Dev. 26, 16–23 (2014)
Stolz, A., Ernst, A. & Dikic, I. Cargo recognition and trafficking in selective autophagy. Nature Cell Biol. 16, 495–501 (2014)
Okamoto, K. Organellophagy: eliminating cellular building blocks via selective autophagy. J. Cell Biol. 205, 435–445 (2014)
Khaminets, A. et al. Regulation of endoplasmic reticulum turnover by selective autophagy. Nature http://dx.doi.org/10.1038/nature14498 (2015)
Noda, N. N., Ohsumi, Y. & Inagaki, F. Atg8-family interacting motif crucial for selective autophagy. FEBS Lett. 584, 1379–1385 (2010)
Farré, J. C., Burkenroad, A., Burnett, S. F. & Subramani, S. Phosphorylation of mitophagy and pexophagy receptors coordinates their interaction with Atg8 and Atg11. EMBO Rep. 14, 441–449 (2013)
Noda, T. & Ohsumi, Y. Tor, a phosphatidylinositol kinase homologue, controls autophagy in yeast. J. Biol. Chem. 273, 3963–3966 (1998)
Hamasaki, M., Noda, T., Baba, M. & Ohsumi, Y. Starvation triggers the delivery of the endoplasmic reticulum to the vacuole via autophagy in yeast. Traffic 6, 56–65 (2005)
Schuck, S., Gallagher, C. M. & Walter, P. ER-phagy mediates selective degradation of endoplasmic reticulum independently of the core autophagy machinery. J. Cell Sci. 127, 4078–4088 (2014)
Kirisako, T. et al. Formation process of autophagosome is traced with Apg8/Aut7p in yeast. J. Cell Biol. 147, 435–446 (1999)
Friedman, J. R. & Voeltz, G. K. The ER in 3D: a multifunctional dynamic membrane network. Trends Cell Biol. 21, 709–717 (2011)
Voeltz, G. K., Prinz, W. A., Shibata, Y., Rist, J. M. & Rapoport, T. A. A class of membrane proteins shaping the tubular endoplasmic reticulum. Cell 124, 573–586 (2006)
Takeshige, K., Baba, M., Tsuboi, S., Noda, T. & Ohsumi, Y. Autophagy in yeast demonstrated with proteinase-deficient mutants and conditions for its induction. J. Cell Biol. 119, 301–311 (1992)
Kvam, E. & Goldfarb, D. S. Nucleus-vacuole junctions and piecemeal microautophagy of the nucleus in S. cerevisiae . Autophagy 3, 85–92 (2007).
Tsukada, M. & Ohsumi, Y. Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae . FEBS Lett. 333, 169–174 (1993)
Park, Y. E. et al. Autophagic degradation of nuclear components in mammalian cells. Autophagy 5, 795–804 (2009)
Shoji, J. Y., Kikuma, T., Arioka, M. & Kitamoto, K. Macroautophagy-mediated degradation of whole nuclei in the filamentous fungus Aspergillus oryzae . PLoS ONE 5, e15650 (2010)
Mijaljica, D. & Devenish, R. Nucleophagy at a glance. J. Cell Sci. 126, 4325–4330 (2013)
Lu, K., Psakhye, I. & Jentsch, S. Autophagic clearance of polyQ proteins mediated by ubiquitin-Atg8 adaptors of the conserved CUET protein family. Cell 158, 549–563 (2014)
Brachmann, C. B. et al. Designer deletion strains derived from Saccharomyces cerevisiae S288C: a useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast 14, 115–132 (1998)
Jones, E. W., Zubenko, G. S. & Parker, R. R. PEP4 gene function is required for expression of several vacuolar hydrolases in Saccharomyces cerevisiae . Genetics 102, 665–677 (1982)
Robinson, J. S., Klionsky, D. J., Banta, L. M. & Emr, S. D. Protein sorting in Saccharomyces cerevisiae: isolation of mutants defective in the delivery and processing of multiple vacuolar hydrolases. Mol. Cell. Biol. 8, 4936–4948 (1988)
Janke, C. et al. A versatile toolbox for PCR-based tagging of yeast genes: new fluorescent proteins, more markers and promoter substitution cassettes. Yeast 21, 947–962 (2004)
Nakatogawa, H., Ishii, J., Asai, E. & Ohsumi, Y. Atg4 recycles inappropriately lipidated Atg8 to promote autophagosome biogenesis. Autophagy 8, 177–186 (2012)
Sikorski, R. S. & Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae . Genetics 122, 19–27 (1989)
James, P., Halladay, J. & Craig, E. A. Genomic libraries and a host strain designed for highly efficient two-hybrid selection in yeast. Genetics 144, 1425–1436 (1996)
Mochida, K., Ohsumi, Y. & Nakatogawa, H. Hrr25 phosphorylates the autophagic receptor Atg34 to promote vacuolar transport of α-mannosidase under nitrogen starvation conditions. FEBS Lett. 588, 3862–3869 (2014)
Suzuki, S. W., Onodera, J. & Ohsumi, Y. Starvation induced cell death in autophagy-defective yeast mutants is caused by mitochondria dysfunction. PLoS ONE 6, e17412 (2011)
Noda, N. N. et al. Structural basis of target recognition by Atg8/LC3 during selective autophagy. Genes Cells 13, 1211–1218 (2008)
Tanaka, C. et al. Hrr25 triggers selective autophagy-related pathways by phosphorylating receptor proteins. J. Cell Biol. 207, 91–105 (2014)
Noda, T., Matsuura, A., Wada, Y. & Ohsumi, Y. Novel system for monitoring autophagy in the yeast Saccharomyces cerevisiae . Biochem. Biophys. Res. Commun. 210, 126–132 (1995)
Acknowledgements
We thank the members of our laboratory for materials, discussions, and technical and secretarial support; Biomaterial Analysis Center, Technical Department at Tokyo Institute of Technology for DNA sequencing service; T. Yoshihisa and T. Endo for providing antibodies. This work was supported in part by Grants-in-Aid for Scientific Research 25111003 (to H.N.), 25711005 (to H.N.), and 23000015 (to Y.Oh.) from the Ministry of Education, Culture, Sports, Science and Technology of Japan.
Author information
Authors and Affiliations
Contributions
K.M. and H.N. designed the study; K.M. performed most of the experiments with the help of H.K.; Y.Oi., Y.K. and H.H. performed mass spectrometry; K.M., Y.Oh. and H.N. analysed data. K.M. and H.N. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
Extended Data Figure 1 Interactions of Atg39 and Atg40 with Atg8 and Atg11.
a, Yeast two-hybrid assay. Cells expressing proteins fused with the Gal4 activation domain (GAD) or the Gal4 DNA-binding domain (GBD) were grown on the agar plate with or without histidine (His). Representative results of two technical replicates are shown. Growth without His indicates positive interaction. The P52A R67A mutation in Atg8 weakens its binding to the Atg8-family-interacting motif (AIM)30. b, In the experiments described in Fig. 1b, the protein band intensities were measured, and the relative ratios of co-precipitated proteins to direct targets of immunoprecipitation (coIP (%)) are shown as the mean ± s.d. (n = 3). WT, wild type. c, Cells were treated with rapamycin for 13 h, and the lysates (input) were subjected to co-immunoprecipitation analysis using anti-GFP antibody. The immunoprecipitates (IP) were analysed by immunoblotting. c, Co-immunoprecipitation efficiencies were calculated as described in b, and the results are shown as the mean ± s.d. (n = 3). d, Cells treated with rapamycin for 12 h were subjected to co-immunoprecipitation analysis as described in c. Eight residues in the Asp52–Ser59 region in Atg39 were replaced with Ala in the D52–S59A mutant. Representative results of three technical replicates are shown (c, d). *P < 0.05; **P < 0.01; ***P < 0.001 (unpaired two-tailed Student’s t-test) (b, c, d).
Extended Data Figure 2 Atg39 and Atg40 are not required for the cytoplasm-to-vacuole targeting pathway, mitophagy, pexophagy, and non-selective autophagy.
a, Cells were incubated in nitrogen starvation medium, and the processing of the proform (p) of the vacuolar aminopeptidase Ape1 into the mature form (m), which is indicative of activity of the cytoplasm-to-vacuole targeting pathway, was examined by immunoblotting using anti-Ape1 antibodies. mApe1 intensity was divided by the sum of mApe1 and pApe1 intensities, and the relative values are shown (Ape1 maturation (%)). Vacuolar degradation of GFP–Atg8, which correlates with activity of non-selective autophagy, was also investigated. The results were quantified (see Methods), and the relative values (%) are shown. b, c, Mitophagy and pexophagy were examined as described previously31. Degradation of GFP-fused proteins was quantified (see Methods), and the relative values (%) are shown. Representative immunoblots of two technical replicates are presented (a–c). d, To assess bulk degradation of the cytoplasm by non-selective autophagy, a Pho8Δ60 alkaline phosphatase (ALP) assay was performed as described previously32. ALP activities are shown as the mean ± s.d. (n = 3). AU, arbitrary units; NS, not significant; ***P < 0.001 (unpaired two-tailed Student’s t-test).
Extended Data Figure 3 The ER is degraded via selective macroautophagy under nitrogen-deprived conditions in a manner dependent on Atg39 and Atg40.
a–e, Cells treated with rapamycin or incubated in nitrogen-starvation medium were subjected to immunoblotting analysis. Pep4 and Ypt7 are required for vacuolar lytic activity and autophagosome-vacuole fusion, respectively. Atg17 is a subunit of the Atg1 kinase complex and required for starvation-induced autophagy1. Atg19 and Atg34 are autophagy receptors for vacuolar enzymes; Atg32 and Atg36 are those for mitochondria and peroxisomes, respectively. In e, atg39Δ or atg40Δ cells harbouring Atg39–HA or Atg40–HA plasmids, respectively, were analysed. Representative immunoblots of three (a, b, d, e) or four (c) technical replicates are shown. Sec63–GFP degradation (%) was quantified (see Methods), and the results are shown as the mean ± s.d. (n = 3 (b, d, e) or 4 (c)). NS, not significant; *P < 0.05; **P < 0.01; ***P < 0.001 (unpaired two-tailed Student’s t-test) (b–e). The results in c and d showed that ER-phagy requires Atg11 and the AIM-binding ability of Atg8, but not previously characterized autophagic receptors, suggesting the existence of an unknown receptor for this pathway. f, Fluorescence microscopy of cells treated with or without rapamycin. Images are representative of at least 150 cells. Arrowheads, Atg8-and Sec63-positive puncta. Scale bars, 5 μm.
Extended Data Figure 4 Atg39 and Atg40 function as ER-phagy receptors.
a, Atg39–mCherry or Atg40–mCherry was expressed under the endogenous promoter (endo.) or the CUP1 promoter for overexpression (OE) in the presence of 250 μM CuSO4. These cells were treated with rapamycin, and degradation of Sec63–GFP was examined by immunoblotting. Pgk1 serves as a loading control. Representative immunoblots of three technical replicates are presented, and the quantification results (see Methods) are shown as the mean ± s.d. (n = 3). *P < 0.05; **P < 0.01 (unpaired two-tailed Student’s t-test). The results for atg1Δ cells suggest that the rapamycin-induced decrease in Atg39–mCherry overexpressed in wild-type cells is due to its autophagic degradation. b, The co-localization of Atg39 or Atg40 with Atg11 in cells treated with rapamycin for 3 h was observed under a fluorescence microscope. Images are representative of at least 100 cells. Scale bars, 2 μm. c–e, Vacuolar degradation of Atg39–GFP and Atg40–GFP was examined by immunoblotting (c, e) and fluorescence microscopy (d). atg39Δ and atg40Δ cells carrying Atg39–GFP and Atg40–GFP plasmids, respectively, were examined in e. Representative immunoblots of three (Atg39–GFP degradation in c, e), or four (Atg40–GFP degradation in c, e) technical replicates are presented, and the quantification results are shown as the mean ± s.d. (n = 3 (Atg39–GFP degradation in c, e) or 4 (Atg40–GFP degradation in c, e)). ***P < 0.001 (unpaired two-tailed Student’s t-test) (c, e). Images are representative of at least 100 cells (d). Scale bars, 5 μm. The results in c–e suggest that vacuolar transport of Atg39 and Atg40 requires their binding to Atg11 and Atg8.
Extended Data Figure 5 Atg39 and Atg40 are differently involved in degradation of the pnER and the cER/cytoER.
Vacuolar transport of Hmg1–GFP and Rtn1–GFP was examined by fluorescence microscopy. Images are representative of at least 100 cells. Scale bars, 5 μm.
Extended Data Figure 6 Electron microscopy of the ER sequestered into autophagic bodies.
a, pep4Δ cells treated with rapamycin for 13 h were analysed by immunoelectron microscopy using anti-Kar2 antibodies. Images are representative of at least 50 cells. Arrowheads, Kar2 signals (gold particles). These images show the staining of the cortical ER (left) and the perinuclear ER/nuclear envelope (right). b–d, Electron microscopy images (without immunogold labelling) of cells incubated in nitrogen-starvation medium for 12 h (b, d) or treated with rapamycin for 12 h (c). Images are representative of at least 14 cells that contained autophagic bodies. Arrowheads, autophagic bodies containing double-ring structures (b, c) or those containing tubules/sheets (d). Scale bars, 200 nm (a–d).
Extended Data Figure 7 Model for Atg39- and Atg40-mediated ER-phagy and nucleophagy.
Atg39 localizes to the perinuclear ER (pnER)/nuclear envelope and acts to load a part of the pnER/nucleus as double-membrane vesicles, which contain intranuclear components, into autophagosomes. Atg40 mainly localizes to highly curved regions in the cortical ER (cER) and cytoplasmic ER (cytoER), and allows sequestration of these ER domains as folded tubules or sheets into autophagosomes. Whereas Atg39 is dispensable for cER/cytoER-phagy, Atg40 also contributes to pnER-phagy/nucleophagy. It is likely that both of these receptors interact with Atg11 to recruit the core Atg proteins that mediate autophagosome formation, and also with Atg8 to promote autophagic sequestration of the ER or a part of the nucleus.
Extended Data Figure 8 Atg39-dependent degradation of part of the nucleus.
a, Src1–GFP degradation assay for cells before rapamycin treatment. The results for cells treated with rapamycin are shown in Fig. 4a. b, Vacuolar transport of Src1–GFP was examined by fluorescence microscopy. Images are representative of at least 100 cells. Scale bars, 5 μm. c, Autophagic degradation of Nop1–GFP was examined by immunoblotting. Representative results of two (a) or three (c) technical replicates are shown. Nop1–GFP degradation was quantified (see Methods), and the results are shown as the mean ± s.d. (n = 3). NS, not significant; ***P < 0.001 (unpaired two-tailed Student’s t-test). d, pep4Δ cells were treated with rapamycin for 13 h, followed by immunoelectron microscopy using anti-Nop1 antibody. Images are representative of six cells containing Nop1 signals in the nucleolus. Scale bars, 500 nm (left) and 200 nm (right). The data show that the nucleolus is specifically labelled with gold particles. Arrowheads, autophagic bodies in the vacuolar lumen.
Extended Data Figure 9 Phenotypes of atg39 and atg40 mutants under nitrogen-deprived conditions.
a, Cells were examined by electron microscopy. Images are representative of at least 14 cells. b, Cells were subjected to nitrogen starvation, and dead cells were stained with phloxine B. Images are representative of at least 600 cells. DIC, differential interference contrast microscopy images. c, Dead cells were counted (see Methods), and the results are shown as the mean ± s.d. (n = 6). d, Wild-type or atg39Δ cells harbouring Atg39–GFP plasmids or the empty vector were subjected to nitrogen starvation, and dead cells were counted. The results are shown as the mean ± s.d. (n = 3). NS, not significant; *P < 0.05; ***P < 0.001 (unpaired two-tailed Student’s t-test) (c, d). e, f, Cells expressing Rtn1–GFP (e) or Sec63–GFP (f) were subjected to nitrogen starvation, and the fluorescence microscopy images were taken by focusing cell periphery (e) or centre (f). Images are representative of at least 100 (e) or 200 (f) cells. In f, the fluorescence intensity of Sec63–GFP at the periphery of cells marked with asterisks (left) was measured along cyan lines in insets and graphically shown (right). Scale bars, 500 nm (a), 5 μm (b, e, f).
Extended Data Figure 10 Domain architectures of Atg40 and reticulon-related proteins.
HHpred, a homology search tool based on structure prediction, identified reticulon-related proteins, Yop1 in S. cerevisiae, and REEP4 and REEP5 in Homo sapiens, as proteins similar to Atg40. Red bars, regions structurally similar to Atg40; green bars, reticulon-like domains; orange regions, hydrophobic segments predicted by TMHMM, a membrane protein topology prediction method. LIR, LC3-interacting region.
Supplementary information
Supplementary Table 1
This file contains a list of yeast strains used. (PDF 218 kb)
Rights and permissions
About this article

Cite this article
Mochida, K., Oikawa, Y., Kimura, Y. et al. Receptor-mediated selective autophagy degrades the endoplasmic reticulum and the nucleus. Nature 522, 359–362 (2015). https://doi.org/10.1038/nature14506
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature14506
This article is cited by
-
Selective autophagy in cancer: mechanisms, therapeutic implications, and future perspectives
Molecular Cancer (2024)
-
The nascent polypeptide-associated complex subunit Egd1 is required for efficient selective mitochondrial degradation in budding yeast
Scientific Reports (2024)
-
ALLO-1- and IKKE-1-dependent positive feedback mechanism promotes the initiation of paternal mitochondrial autophagy
Nature Communications (2024)
-
Autophagy: Regulator of cell death
Cell Death & Disease (2023)
-
The mechanisms to dispose of misfolded proteins in the endoplasmic reticulum of adipocytes
Nature Communications (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
