
Abstract
Interleukin-10 (IL-10) is an important regulatory cytokine required to control allergy and asthma. IL-10-mediated regulation of T cell-mediated responses was previously thought to occur indirectly via antigen-presenting cells. However, IL-10 can act directly on regulatory T cells and T helper type 17 (Th17) cells. In the context of allergy, it is therefore unclear whether IL-10 can directly regulate T helper type 2 (Th2) cells and whether this is an important regulatory axis during allergic responses. We sought to determine whether IL-10 signaling in CD4+ Th2 cells was an important mechanism of immune regulation during airway allergy. We demonstrate that IL-10 directly limits Th2 cell differentiation and survival in vitro and in vivo. Ablation of IL-10 signaling in Th2 cells led to enhanced Th2 cell survival and exacerbated pulmonary inflammation in a murine model of house dust mite allergy. Mechanistically, IL-10R signaling regulated the expression of several genes in Th2 cells, including granzyme B. Indeed, IL-10 increased granzyme B expression in Th2 cells and led to increased Th2 cell death, identifying an IL-10-regulated granzyme B axis in Th2 cells controlling Th2 cell survival. This study provides clear evidence that IL-10 exerts direct effects on Th2 cells, regulating the survival of Th2 cells and severity of Th2-mediated allergic airway inflammation.
Similar content being viewed by others


INTRODUCTION
Interleukin-10 (IL-10) is a potent regulator of inflammatory responses1 and plays a critical role in controlling allergic airway inflammation.2 Compared with nonasthmatics, asthmatic individuals have been reported to have reduced levels of IL-10 in bronchoalveolar lavage fluid,3 with decreased secretion of IL-10 from alveolar macrophages.4 Furthermore, polymorphisms in the IL10 gene resulting in low IL-10 production have been associated with severe asthma,5 suggesting that IL-10 is a major determinant in the development and severity of airway allergy. In murine models, Il10–/– mice develop enhanced allergic responses to a variety of allergens6, 7, 8 and develop increased eosinophilic airway inflammation. These studies highlight a clear non-redundant role for IL-10 in controlling allergen-induced airway disease. To date, the only preventative treatment for allergic asthmatics is allergen-specific immunotherapy.9 Although the precise mechanisms are unclear, successful allergen-specific immunotherapy correlates with the development of IL-10-secreting antigen-specific T cells,10 suggesting that the balance between IL10-secreting cells and pathogenic cells is an important determinant in the development of allergic disease.11
Despite the central role of IL-10 in controlling allergic responses, the target of IL-10 signaling in this setting has not been clearly described. Classically, the effects of IL-10 on T-cell responses were thought to occur indirectly through antigen-presenting cells (APCs).12, 13 Recent reports, however, indicate that IL-10 can signal directly on subsets of T cells, including memory/effector T cells in a model of colitis,14 T helper type 17 (Th17) cells during intestinal inflammation,15 and regulatory T (Treg) cells for the prevention of Th17-mediated colitis.16 In addition, in vitro data suggested that IL-10 can directly regulate IL-5 and IL-4 in cultured T cells.17, 18 Therefore, we hypothesized that IL-10 signaling directly in T helper type 2 (Th2) cells could contribute to regulation of Th2-mediated allergic airway responses. Here, we investigated the role of IL-10 signaling in Th2 cells in a mouse model of house dust mite (HDM)-induced allergic airway inflammation and showed that IL-10 signaling in Th2 cells regulated the severity of allergic airway responses. RNA sequencing (RNA-Seq) identified that IL-10 regulated several genes in Th2 cells including Granzyme b. We found that IL-10 regulated granzyme B in Il10–/– cells that correlated with increased Th2 cell death. Finally, adoptive transfer of Il10r-deficient Il4gfp+ Th2 cells alone was sufficient to orchestrate severe allergic airway inflammation in wild-type (WT) mice, identifying that IL-10-mediated regulation of Th2 cells in vivo is an important and previously overlooked axis of immune regulation, preventing the development of severe airway inflammation.
RESULTS
IL-10R signaling in T cells regulates Th2 cell differentiation and HDM-induced airway allergy
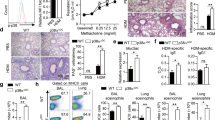
To investigate whether IL-10 signaling in T cells was important during airway allergy, we conditionally deleted the IL-10 receptor-α (IL-10RA) subunit in T cells by crossing Il10rafl/fl mice with Cd4Cre mice.19 Mice were sensitized to HDM via the intraperitoneal route and were challenged by intratracheal delivery of HDM to establish a localized T cell-driven airway inflammatory response (Figure 1a). HDM-challenged mice with Il10ra-deficient T cells had significantly increased cellular infiltration and eosinophilia in the alveolar spaces, compared with Il10ra-sufficient mice (Figure 1b,c). Mice lacking IL-10 signaling in T cells also developed more severe lung pathology with increased inflammation (Figure 1d), mucus secretion, Muc5ac gene expression (Figure 1e,f), and elevated serum IgE (Figure 1g). The frequency and total number of CD4+ T cells expressing IL-4, IL-5, and IL-13 were also significantly increased in the lung and lung-draining mediastinal lymph nodes of Cd4CreIl10rafl/fl mice (Figure 2a–d and Supplementary Figure S1A–D online). Although IL-10 can also regulate both type-1 and type-17 responses following allergen challenge in the airways,7, 20 we found that the frequency of IFNγ+ and IL-17A+ T cells in the lung were significantly reduced, with numbers of interferon-γ (IFNγ)- and IL-17A-producing CD4+ cells unaffected by the absence of IL-10 signaling in T cells (Figure 2e,f and Supplementary Figure S1C,D). The proportion of Foxp3+ Treg cells in the local lymph nodes were largely unaffected; however, we did observe an increase in Foxp3+ Treg cells in the lung of Cd4CreIl10rafl/fl mice (Supplementary Figure S1E,F). Overall, these data identify a role for IL-10 signaling in T cells during HDM-induced airway allergy.

Interleukin-10 receptor (IL-10R) signaling in T cells regulates house dust mite (HDM)-mediated airway inflammation. (a) Schematic of HDM sensitization and challenge. Cd4CreIl10rafl/fl or Cd4CreIl10ra+/+ mice were sensitized with HDM or phosphate-buffered saline (PBS) and alum intraperitoneally (i.p.) at days 0 and 14, challenged with HDM intratracheally (i.t.) on days 21 and 24, and harvested on day 25. (b) Total number of cells recovered from bronchoalveolar lavage (BAL). (c) Differential counts on Giemsa-stained BAL cells. (d) Representative hematoxylin and eosin (H&E)-stained lung sections and inflammation scores from HDM-challenged mice. (e) Representative Alcian blue/periodic acid–Schiff (AB-PAS)-stained lung sections and mucus scores from HDM-challenged mice. (f) Expression of Muc5ac in lung tissue. (g) Serum IgE in PBS or HDM-challenged mice. Data are representative of four separate experiments with Mann–Whitney tests used to determine significance with P⩽0.05 considered significant, marked with an asterisk (*); n=4–6 mice per group.

Interleukin-10 receptor (IL-10R) signaling in T cells limits T helper type 2 (Th2) differentiation in vivo and in vitro. (a–f) Cd4CreIl10rafl/flIl4gfp or Cd4CreIl10ra+/+Il4gfp mice were sensitized with house dust mite (HDM) or phosphate-buffered saline (PBS) and alum intraperitoneally (i.p.) at days 0 and 14, challenged with HDM intratracheally (i.t.) on days 21 and 24, and harvested on day 25, as in Figure 1. Percent and number of cytokine-secreting CD4+CD44hi cells in the lung and mediastinal lymph nodes (med:LN). Data representative of 2–4 separate experiments; n=4–6 mice per group. (g) Naïve cells from Cd4CreIl10rafl/flIl4gfp or Cd4CreIl10ra+/+Il4gfp mice were polarized in vitro for 7 or 14 days, as indicated, and stained for intracellular cytokines by flow cytometry. Data are representative of three separate experiments. (h) Naïve cells from Cd4CreIl10rafl/flIl4gfp or Cd4CreIl10ra+/+Il4gfp mice were harvested for cell lysates or polarized under Th2 conditions for 3 days. Total cultures were then harvested for lysates and western blot analysis for phosphorylated signal transducer and activator of transcription 3 (pSTAT3), total STAT3, pSTAT6, total STAT6, and Actin as a loading control. Data are representative of two separate experiments with densitometry used to determine the ratio of phosphorylated/total protein level. Mann–Whitney tests were used to determine significance with P⩽0.05 considered significant, marked with an asterisk (*). AU, arbitrary unit.
CD4 is expressed on a subset of dendritic cells and transiently expressed on CD8+ cells during T-cell development. It was therefore possible that the in vivo observations described above were a consequence of IL-10R deletion on CD4+ dendritic cells or CD8+ T cells. To determine whether IL-10 could directly regulate cytokine production in CD4+ T cells, we used fluorescence-activated cell sorting (hereafter referred to as ‘sorting’) to purify naive TCRβ+ CD4+ T cells and polarized these under Th2 conditions in the absence of APCs. We observed a higher frequency of IL-4+ and IL-13+ cells from as early as day 7 in Cd4CreIl10rafl/fl Th2 cultures compared with Cd4CreIl10ra+/+cultures, with a further increase in IL-4+, IL-13+, and IL-5+ cells after 14 days (Figure 2g), indicating that IL-10 can directly regulate Th2 cell differentiation.
IL-10 induces tyrosine phosphorylation of signal transducer and activator of transcription 3 (STAT3) in monocytes and lymphocytes.21 To determine whether STAT3 expression and phosphorylation was regulated by IL-10 in Th2 cells, we measured STAT3 in Il10ra-sufficient and Il10ra-deficient naive T cells and in Th2 committed cells following 3 days of Th2 polarization. Il10ra-sufficient and -deficient naive T cells had similar levels of total STAT3 and, as expected, very little phosphorylated STAT3 (pSTAT3). Of note, there was a reduction in total pSTAT3 in Cd4CreIl10rafl/fl Th2 cells compared with Cd4CreIl10ra+/+Th2 cells (Figure 2h and Supplementary Figure S2A), indicating that IL-10R-signaling was required for optimal STAT3 levels and phosphorylation of STAT3. WT T cells cultured under Th2 conditions in the presence of an IL-10R blocking antibody (anti-IL-10R) also had reduced pSTAT3 as compared with WT cells treated with an isotype control antibody (Supplementary Figure S2B). Il10ra-deficient T cells also secreted less IL-10 under Th2 conditions in vitro (Supplementary Figure S2C) with proportionally fewer IL-10+ cells in the lungs of mice with HDM-induced airway allergy (Supplementary Figure S1G,H), suggesting that IL-10R signaling was required for optimal IL-10 production from Th2 cells. These data are in line with a previous report demonstrating that IL-10R-signaling was required for IL-10 production by Treg cells.16 STAT6 was largely unaffected in Il10ra-deficient T cells (Figure 2h).
Tissue-associated Th2 cells preferentially express Il10 and Il10ra
To determine the spatial expression of the IL-10R on Th2 cells in vivo, we sensitized and challenged Il4gfpreporter mice22 with HDM and sorted Th2 committed Il4gfp+cells from the lung tissue, local mediastinal lymph nodes, and spleen following HDM challenge. Th2 cells from the lung tissue had the highest relative expression of both Il10 and Il10ra as compared with Th2 cells from the spleen or mediastinal lymph nodes (Figure 3a,b). Th2 cells in the lung also had a slightly higher expression of Il10ra compared with other T cells in the lung (Supplementary Figure S3A). To determine whether Il10 and Il10r were coregulated in lung-derived Th2 cells, we crossed Il4gfp-reporter mice with Il10-reporter mice (10BiT).23 These dual reporter mice allowed us to sort-purify Il10-expressing Th2 cells (Il10BiT+Il4gfp+) and nonexpressing Th2 cells (Il10BiT–Il4gfp+) from the allergic lung (Figure 3c). As expected, Il10BiT+ Th2 cells had elevated expression of Il10 mRNA (Figure 3d); however, both Il10BiT+ and Il10BiT– Th2 cells had comparable Il10ra expression (Figure 3e), suggesting that Il10 and Il10r are differentially regulated in Th2 cells. The common IL-10 receptor (Il10Rb), which pairs with various cytokine receptors for IL-10, IL-22, IL-26, IL-28, and IFN1 signaling, was not differentially regulated in any of the purified Th2 cells analyzed (data not shown).

Interleukin-10 (IL-10) and the IL-10 receptor-α are highly expressed in lung T helper type 2 (Th2) cells during house dust mite (HDM) challenge. (a, b) Il4gfp reporter mice were sensitized and challenged with HDM. At day 25, as in Figure 1a, CD4+TCRβ+Il4gfp+ cells were sorted from mediastinal lymph nodes (med:LN), spleens, and lungs. RNA was harvested and analyzed by reverse transcription-PCR (RT-PCR) for expression of Il10 and Il10ra, and expressed relative to Hprt control. Data are representative of two independent experiments; sorted cells were pooled from four mice in each experiment. (c–e) Il4gfpFoxp3rfpIl10BiT mice were sensitized and challenged with HDM. At day 25, CD4+TCRβ+Il4gfp+Il10BiT+or CD4+TCRβ+Il4gfp+Il10BiT− cells were sorted from lungs (c), harvested for RNA, and analyzed by RT-PCR for expression of Il10 and Il10ra, relative to Hprt. Data are representative of two independent experiments; sorted cells were pooled from five mice per group.
IL-10-induced granzyme B in Th2 cells contributes to Th2 cell death
To identify potential mechanisms of how IL-10R signaling regulated Th2 cells during allergic airway inflammation, we crossed Cd4CreIl10rafl/fl or Cd4CreIl10ra+/+ mice with Il4gfp reporter mice,22 allowing us to sort-purify Il10ra-sufficient and Il10ra-deficient Il4gfp+ Th2 cells from the lung tissue of allergic mice (Figure 4a,b). RNA was extracted from enriched and sort-purified lung-derived Il4gfp+Th2 cells and used for transcriptional analysis using RNA-Seq. The transcriptome of Il10ra-sufficient and Il10ra-deficient Il4gfp+ Th2 cells were surprisingly similar with only 36 genes significantly different (P<0.05, >2-fold change, Figure 4c). Applying a fivefold change filter between Il10ra-sufficient and Il10ra-deficient Il4gfp+ Th2 cells and a minimum read count of 20, we identified 6 genes that were significantly different between Il10ra-sufficient and Il10ra-deficient Il4gfp+ Th2 cells (Figure 4d). These genes included Il10ra and Gzmb (granzyme B), which were downregulated in Il10ra-deficient Il4gfp+ Th2 cells, and Bc049352, Tmprss13, Scn4b, and Fxyd6, which were upregulated in Il10ra-deficient Il4gfp+ Th2 cells (Figure 4e).

Interleukin-10 (IL-10) regulates inhibitory receptor pathways and chemokine expression in pulmonary T helper type 2 (Th2) cells. (a) Experimental schematic. (b) CD4+TCRβ+Il4gfp+cells were sorted from the lungs of house dust mite (HDM)-challenged Cd4CreIl10rafl/flIl4gfp or Cd4CreIl10ra+/+Il4gfp mice on day 25. (c) RNA sequencing (RNA-seq) was performed on sorted cells. Plot indicating number of normalized reads for each gene in Cd4CreIl10rafl/flIl4gfp+(y axis) against Cd4CreIl10ra+/+Il4gfp+ Th2 cells (x axis). (d) Plot showing genes that were significantly different between Il10rafl/flIl4gfp+and Il10ra+/+Il4gfp+ Th2 cells. Data presented as fold change difference between Il10rafl/fl and Il10ra+/+ Il4gfp+ Th2 cells (x axis) with normalized read count (y axis). Black circles depict genes with >20 reads that were significantly increased in knockout (KO) cells and black triangles depict genes significantly decreased in KO cells. (e) Six genes that were fivefold different between Il10ra-sufficient and Il10ra-deficient Il4gfp+ Th2 cells with a minimum read count of 20. P≤0.05 considered significant, marked with an asterisk (*).
Following the report that granzyme B expression in Th2 cells was required for T-cell receptor (TCR)-induced Th2 cell death,24 we hypothesized that IL-10R signaling was required for granzyme B-mediated Th2 cell death, providing an explanation for exacerbated Th2-mediated inflammation in mice with Il10ra-deficient T cells. Granzyme B protein was also reduced in Il10ra-deficient Th2 cells generated in vitro as compared with Il10ra-sufficient Th2 cells (Figure 5a,b), in line with the RNA-Seq data (Figure 4e). Decreased granzyme B expression in Il10ra-deficient T cells correlated with reduced cell death, determined by propidium iodide (PI) incorporation (Figure 5c). Reduced granzyme B and reduced cell death in Il10ra-deficient Th2 cell cultures resulted in increased Th2 cells (Figure 5d). These data support the hypothesis that IL-10R signaling is required for granzyme B expression in Th2 cells leading to Th2 cell death. To test whether IL-10 was specifically required for granzyme B expression in Th2 cells and for Th2 cell death, we polarized Il10–/– CD4+ T cells under Th2 conditions and observed that Il10–/– T cells had reduced granzyme B, similar to Il10ra-deficient T cells. Granzyme B expression could be rescued by the addition of recombinant IL-10 (Figure 5e,f), indicating that IL-10 is both necessary and sufficient for granzyme B expression in Th2 cells. In these cultures, IL-10-induced granzyme B in recombinant IL-10-treated Il10–/– T cells correlated with increased cell death and reduced Th2 cell numbers (Figure 5g,h). Finally, granzyme B inhibitor treatment (z-AAD-CH2Cl) reduced the frequency of granzyme B+ cells and prevented IL-10-induced granzyme B expression (Figure 5i). Granzyme B inhibitor treatment also prevented IL-10-induced Th2 cell death (Figure 5j) and resulted in increased Th2 cells (Figure 5k), indicating that IL-10-induced granzyme B activity was required for IL-10-induced Th2 cell death. Taken together, these data indicate that IL-10R signaling in Th2 cells is required for granzyme B-mediated Th2 cell death.

Interleukin-10 receptor (IL-10R) signaling in T helper type 2 (Th2) cells regulates granzyme B-mediated Th2 cell death. (a, b) Naïve Cd4CreIl10rafl/flIl4gfp– or Cd4CreIl10ra+/+Il4gfp– T cells (CD4+TCRβ+CD44–) were differentiated under Th2 conditions for 7 days. Live, CD4+TCRβ+PI–Il4gfp+ cells were sorted and 105 cells per well were stimulated on flat-bottomed anti-CD3-coated plates (10 μg ml−1) for 24 h. The frequency of granzyme B+ cells was determined after 24 h of stimulation. (c, d) The total number of live and T-cell receptor (TCR)-induced dead cells was determined by propidium iodide (PI) staining. (e, f) Naïve WT (Il10+/+) and Il10–/– T cells were differentiated under Th2 conditions for 7 days. Live, CD4+TCRβ+PI– cells were sorted and 105 cells per well were stimulated on flat-bottomed anti-CD3-coated plates (10 μg ml−1) for 24 h with or without recombinant IL-10 (rIL-10, 10 ng ml−1). The frequency of granzyme B-expressing cells was determined after stimulation. (g, h) The total number of live and TCR-induced dead cells was determined by PI staining. (i) Naïve WT (Il10+/+) and Il10–/– T cells were differentiated under Th2 conditions for 7 days, as in (e) above. Live, CD4+TCRβ+PI– cells were sorted and 105 cells per well were stimulated on flat-bottomed anti-CD3-coated plates (10 μg ml−1) for 24 h with or without rIL-10 (10 ng ml−1) in the presence or absence of a granzyme B inhibitor (GrB In, 10 μM). The frequency of granzyme B-expressing cells was determined after stimulation. (j, k) The total number of live and TCR-induced dead cells was determined by PI staining. Mann–Whitney tests used to determine significance with P⩽0.05 considered significant, marked with an asterisk (*). Data are representative of two independent experiments.
IL-10R signaling, specifically in Il4gfp+ Th2 cells, regulates the magnitude of HDM-driven Th2 responses in vivo
To test whether Il10ra-deficient Th2 cells were indeed responsible for the increased airway inflammation in vivo and whether IL-10 acted directly on Th2 cells, we developed an adoptive transfer system, transferring activated congenic CD4+CD44+Il4gfp+ Th2 cells from HDM-immunized Il10ra-sufficient (Figure 6a, Group 1, CD45.1+Cd4CreIl10ra+/+Il4gfp+) or Il10ra-deficient (Figure 6a, Group 2, CD45.1+Cd4CreIl10rafl/flIl4gfp+) mice to WT mice. Recipient mice were given localized HDM challenges to reactivate transferred Th2 cells (Figure 6a). This system removed the influence of Il10ra-deficient CD4+ dendritic cells, Il10ra-deficient CD8+ cells, and Il10ra-deficient Treg cells16 in vivo. HDM-mediated reactivation of transferred Il10ra-deficient Th2 cells led to an increased influx of cells into the alveolar space (bronchoalveolar lavage), with significantly more eosinophils and macrophages compared with mice that received Il10ra-sufficient Th2 cells (Figure 6b,c). Il10ra-deficient Th2 cells also supported the differentiation and recruitment of host (CD45.2+) T cells into Th2 cells with increased host IL-5+ and IL-13+ Th2 cells in the lungs of recipient mice (Figure 6d–f).

Interleukin-10 receptor (IL-10R) signaling in T helper type 2 (Th2) cells regulates the magnitude of Th2 responses in vivo. (a) Experimental schematic: CD45.1+CD4+TCRβ+Il4gfp+Th2 cells were sorted from spleens and mediastinal lymph nodes (med:LN) of house dust mite (HDM)-sensitized Cd4CreIl10rafl/flIl4gfp or Cd4CreIl10ra+/+Il4gfp mice on day 18. CD45.2+ wild-type (WT) recipient mice were given 0.4–1.0 × 106 Th2 cells intravenously (i.v.) 1 day after the first HDM challenge. Recipient mice were challenged twice more on days 19 and 22 and lungs were harvested on day 23. (b) Total numbers of cells recovered from bronchoalveolar lavage (BAL) of mice receiving no cells, Cd4CreIl10ra+/+Il4gfp+Th2 cells, or Cd4CreIl10rafl/flIl4gfp+Th2 cells. (c) Differential counts on Giemsa-stained BAL cells. (d–f) Numbers of cytokine-expressing CD4+CD44hi recipient (CD45.2+) cells in the lungs. (g–j) CD45.2+ WT recipient mice were given 0.5 × 106 CD45.1+Cd4CreIl10ra+/+Il4gfp and 0.5 × 106 of CD45.1+/–Cd4CreIl10rafl/flIl4gfp cells, as in a, Group (Gp.) 3. On day 23, the percentage of donor WT (CD45.1+) and knockout (KO, CD45.1+CD45.2+) cells in the lung of recipient mice is shown (g, h). The geometric mean fluorescence intensity (gMFI) for Il4gfp (h, i) and granzyme B (h, j) is shown. Data are representative of two separate experiments with Mann–Whitney tests used to determine significance with P⩽0.05 considered significant, marked with an asterisk (*); n= 3–5 mice per group.
Finally, to determine whether Il10ra-deficient Th2 cells survived better than Il10ra-sufficient cells in vivo, we used a similar adoptive transfer system as above (Figure 6a), but cotransferred congenically marked Cd45.1+/– Il10ra-deficient Il4gfp+ Th2 cells and Cd45.1+Il10ra-sufficient Il4gfp+ Th2 cells in equal numbers into recipient Cd45.2+ mice, normalizing any differences in the local inflammatory milieu (Figure 6a, Group 3). Using this system, we observed that Il10ra-deficient Th2 cells survived significantly better in the lung and local lymph node as compared with Il10ra-sufficient Th2 cells (Figure 6g,h). Furthermore, Il10ra-deficient Th2 cells further upregulated Il4gfp, with greater fluorescence intensity, than Il10ra-sufficient Th2 cells (Figure 6h,i) and had less granzyme B expression in the lung, but not the local lymph node (Figure 6h,j), supporting the notion that local IL-10R signaling significantly regulates Th2 cell survival, in part via granzyme B. Taken together, these data demonstrate that IL-10R signaling in Th2 cells is required to control Th2 cell survival in vivo and functions as an important regulatory axis limiting Th2-mediated allergic responses in the lung.
DISCUSSION
In summary, our data have identified that IL-10 plays a critical role in directly regulating Th2 cells during HDM-induced airway inflammation. We report that mice with T cells deficient in Il10ra have increased allergic airway and lung inflammation and that IL-10 directly regulates Th2 cell differentiation and survival. The specific loss of IL-10R signaling in HDM-elicited Th2 cells resulted in a variety of transcriptional changes, including the significant reduction of granzyme B. In vitro, we demonstrate that IL-10-induced granzyme B in Th2 cells contributes significantly to TCR-induced Th2 cell death, providing a mechanistic explanation of how IL-10R signaling directly regulates Th2-mediated responses.
The heightened allergic response we observed in Cd4CreIl10rafl/fl mice was broadly in line with previous studies that define the importance of IL-10 as a regulatory cytokine in allergic inflammation.2 However, our data identify that IL-10 also functions as a direct regulator of T-cell differentiation and that IL-10 signaling in T cells is critical to prevent severe allergic responses (Figures 1 and 2). Furthermore, we demonstrate that IL-10 is important in regulating Th2 cell differentiation in vivo, and that IL-10RA signaling in differentiated Th2 cells alone is required to prevent allergic responses (Figure 5). Although IL-10 regulation of T-cell responses has classically been attributed to indirect action via APCs,12, 13 our data add IL-10 regulation of Th2 cells to the growing body of literature showing that IL-10 can act directly on T cells.14, 15, 16
In addition to heightened allergic responses in Cd4CreIl10rafl/fl mice, we observed enhanced Th2 cytokine-expressing cells in the lungs and lymph nodes (Figure 2). In vitro, Th2 cells polarized more readily toward the Th2 fate (Figure 2). This is in accordance with previous data showing that IL-5 and IL-4 can be directly regulated by IL-10 in T-cell cultures.17, 18 However, when we purified Il4gfp+Th2 cells from allergic Cd4CreIl10rafl/fl or Cd4CreIl10ra+/+mice, we find similar expression of Il4, Il5, and Il13 when RNA was normalized, suggesting that this regulation occurs on a population level. In this HDM-induced allergy model, Th1 and Th17 responses were unchanged or reduced in Cd4CreIl10rafl/fl mice (Figure 2e,f), suggesting that IL-10-mediated regulation of these responses is either context specific15 or occurs via APCs during airway allergy.
Both STAT3 and pSTAT3 were reduced in Cd4CreIl10rafl/fl Th2 cells (Figure 2h), indicating that IL-10 contributes to STAT3 phosphorylation and total STAT3 levels, possibly via auto regulation of STAT3.25 In addition, Il10rafl/fl Th2 cells secreted less IL-10 in vitro that may contribute to reduced pSTAT3 levels in Th2 cells and explain the enhanced inflammatory response in vivo. Residual pSTAT3 in Il10ra-deficient Th2 cells was sufficient for Th2 differentiation, unlike complete Stat3–/– T cells that fail to differentiate into Th2 cells.26 Taken together, these data demonstrate that IL-10R signaling in T cells regulates the magnitude and severity of airway allergy. In particular, IL-10R signaling via STAT3 regulates Th2 cell differentiation in vitro and in vivo following HDM exposure.
The kinetics and regulation of Il10 and Il10ra expression in T cells has been the focus of many studies. We observed that both Il10 and Il10ra were coexpressed greatest in tissue-derived Th2 cells as compared with lymph node-derived Th2 cells (Figure 3). IL-4, GATA-3, and STAT6 are required for IL-10 production from Th2 cells,27, 28, 29 but the precise signaling events that induce Il10 expression in Th2 cells in vivo are unclear. Our data suggest that either local inflammatory or tissue-derived signals stimulate Il10 expression in Th2 cells. For example, tissue-derived Th2 cells respond in an antigen receptor-independent, innate-like manner, producing IL-13 in response to IL-33.30 Whether local IL-33 also regulates IL-10 expression in Th2 cells is currently unclear. IL-10 signaling in Th2 cells was required for IL-10 production, as tissue-derived or in vitro generated Il10ra-deficient Th2 cells had lower Il10 expression than Il10ra-sufficient Th2 cells. Whether an IL-10 autocrine regulatory loop supports IL-10 secretion in Th2 cells or whether B cell- and T cell-derived IL-10, which also potently suppresses innate and adaptive immune responses associated with airway allergy,31, 32 regulates Th2 cell-derived IL-10 is currently unclear. An autocrine IL-10 loop is supported by the coexpression of Il10 and Il10ra in tissue-derived Th2 cells, and is in line with observations in Treg cells.16
IL-10R signaling also regulated granzyme B expression in Th2 cells. Although it was previously unclear how granzyme B was regulated in Th2 cells, it has been shown that IL-10 can induce granzyme B expression in CD8+ T cells,33 and we now show here that IL-10 can regulate granzyme B in CD4+ T cells. Similarly, it has been known for several years that CD4+Th2 cells,34, 35 CD4+ cytotoxic Th1-like cells,36 and CD4+ Treg cells37 express granzyme B with functional studies, using granzyme B-deficient mice or granzyme B inhibitors, clearly demonstrating that granzyme B is required for TCR-induced Th2 cell death.24, 35 Taken together with these previous observations, we now provide evidence that IL-10R signaling in Th2 cells provides an upstream mechanism, upregulating granzyme B in Th2 cells for TCR-induced Th2 cell death. This pathway provides an explanation for the elevated Th2 cell-mediated airway inflammation observed in mice with, or given, Il10ra-deficient Th2 cells. In particular, Th2 cells in the lung, which had the highest expression of Il10ra and produced the highest amount of Il10, appeared to be more tightly regulated by IL-10 than Th2 cells in the local lymph node (Figure 6). These observations lead to the hypothesis that IL-10 may function to directly regulate T cells in the tissue as compared with IL-10 acting on APCs in the lymph node; however, this hypothesis requires testing.
Overall, these data provide new insight into IL-10-mediated regulation of allergic airway inflammation and identify that pathogenic Th2 cells are an important direct target of IL-10. Thus, therapeutic use of IL-1020, 38 to treat airway inflammatory conditions may directly impact antigen-reactive pathogenic T cells, expanding the broad immunosuppressive properties of IL-10.
METHODS
Animals. All mice were bred and maintained under specific pathogen-free conditions at The Francis Crick Institute (formerly the MRC National Institute for Medical Research, London, UK). Cd4CreIl10rafl/fl Il4gfp mice were developed by crossing CD4Cre mice with Il10rafl/fl19 and Il4gfp mice.22 Cd4CreIl10ra+/+Il4gfp mice were used as controls. WT B6/J CD45.1 mice were bred in-house. Il4gfpFoxp3rfpIl10Bit mice were generated by crossing 4get,39 FIR,40 and IL-10BiT reporter mice.23 In vivo experiments were performed using female mice; all mice were on the C57Bl/6 background. All animal experiments were carried out following UK Home Office regulations (project license 80/2506) and were approved by The Francis Crick Institute Ethical Review Panel.
Cell sorting and flow cytometry. Cell sorting was performed using a FACS Aria II (BD Biosciences, San Jose, CA), MoFlo XDP (Beckman Coulter, Brea, CA), or Influx (BD Biosciences) cell sorter. For lungs, tissue was first digested for 45 min at 37 °C in 1 mg ml−1 Liberase (Roche, Basel, Switzerland) with 0.024 mg ml−1 DNase I (Roche) in Iscove’s modified Dulbecco’s medium (Gibco, Thermo Fisher, Waltham, MA) and filtered through a 40 μM cell strainer. To prepare cells for sorting, CD4+ cells were first positively selected using MACS CD4 beads and magnetic columns (Miltenyi Biotec, Bergisch Gladbach, Germany) and were then stained for 25 min with antibodies in phosphate-buffered saline (PBS) with 1% fetal calf serum FCS. To prepare for sorting, stained cells were diluted in phenol-red free Iscove’s modified Dulbecco’s medium (Gibco) (with 1% fetal calf serum, 2 mM EDTA (Invitrogen, Carlsbad, CA), 100 U ml−1 penicillin, and 100 μg ml−1 streptomycin (Gibco), 8 mM L-glutamine (Gibco), and 0.05 mM 2-mercaptoethanol (Gibco)). PI was used to determine cell viability. Intracellular cytokine staining was performed following 6 h of restimulation with 50 ng ml−1 phorbol 12-myristate 13-acetate (Promega, Madison, WI) and 1 μg ml−1 ionomycin (Sigma, St Louis, MO). BD Golgi Stop and BD Golgi Plug (diluted 1:1,000, BD Biosciences) were included for 6 h for ex vivo experiments and for the last 3 h of restimulation for in vitro experiments. Following surface stain, cells were fixed with 2% formaldehyde in PBS (Sigma) and then incubated with antibodies in Permeabilization buffer (eBioscience, San Diego, CA) for a further 30 min. Cells were analyzed using a BD LSRII (BD Biosciences) and data were analyzed using FlowJo software (Version 7.6.5, Treestar, Ashland, OR). Antibodies used included: CD4 (efluor450 and PE-Cy7, RM4-5, eBioscience), CD25 (Fitc, 7D4, BD Pharmin, San Jose, CA), CD44 (Fitc, Percpcy5.5, and APC, IM7, eBioscience), CD45.1 (PE-Cy7 and APC, A20, eBioscience), IFNγ (PE, XMG1.2, BD Pharmin), IL-4 (PE, 11B11, eBioscience), IL-5 (APC, TRFK5, BD Pharmingen, San Jose, CA), IL-13 (eFluor660, eBio13A, eBioscience), TCRβ (APC, H57-597, eBioscience), IL-17A (PECy7, eBio17B7, eBioscience), granzyme B (Alexa-647 or PE, GB11, Biolegend, BD Pharmingen), and phospho-STAT3 (Y705) (LUVNKLA, BD Pharmingen). Staining was performed in the presence of FcR Blocking Reagent (Miltenyi Biotec). In analysis experiments, viability was determined using the Molecular Probes Live/Dead Fixable Blue Dead Cell Stain Kit (Life Technologies, Carlsbad, CA).
HDM-induced airway allergy. Mice were sensitized intraperitoneally. with 10 μg HDM (Greer, Lenoir, NC) or PBS and 2 mg Imject Alum (Thermo Scientific) and challenged intratracheally with 10 μg HDM or PBS. Bronchoalveolar lavage using 1.5 ml PBS was performed to recover cells for Cytospin and differential counts, following staining with Accustain Wright-Giemsa stain, modified (Sigma). Then, 2–3 lung lobes were harvested for flow staining experiments. For histology, lungs were fixed in 4% formaldehyde (VWR, Radnor, PA) followed by 70% ethanol and then prepared for staining with Alcian blue/periodic acid–Schiff and hematoxylin and eosin. Slides were scored on a 0–4 scale for mucus and inflammation, as previously described,6 with modifications. Briefly, for mucus score: the entire lung section was graded for presence of Goblet cells, 1; having more than 3 affected bronchioles, 2; having the entire ring stained in more than 3 bronchioles, 3; and presence of mucus plugs, 4. This score was averaged with an average score for five individual airways graded as: presence of Goblet cells, 1; 50% of bronchiole stained, 2; 100% of bronchiole stained, 3; and mucus plug, 4. Similarly, inflammation was graded as an average of the entire lung section score with the average scores of five individual airways. For entire lung section: presence of perivascular inflammation, 1; inflammation around 3 or more bronchioles, 2; dense inflammatory foci 3 cells deep, 3; and loss of lung architecture, 4. For each individual airway: presence of inflammation, 1; 50% inflammation around bronchiole, 2; 100% inflammation around bronchiole, 3; and dense inflammation bridging two bronchioles, 4. Half scores were given if between grades.
Adoptive cell transfer. CD4+TCRβ+Il4gfp+ Th2 cells were sorted from spleens and thoracic lymph nodes of Cd4CreIl10ra+/+Il4gfp (Group 1) or Cd4CreIl10rafl/flIl4gfp (Group 2) or in a 50:50 mix Cd4CreIl10ra+/+Il4gfp and Cd4CreIl10rafl/flIl4gfp cells from HDM-sensitized mice (HDM and alum delivered intraperitoneally at day 0 and day 14; sort on day 18). 0.5–1.0 × 106 Th2 cells were given intravenously to WT B6/J CD45.2 recipients.
Enzyme-linked immunosorbent assay. Total IgE enzyme-linked immunosorbent assay was performed by coating with Purified Rat Anti-Mouse IgE (R35-72, BD Pharmingen) at 2 μg ml−1 overnight, followed by overnight incubation with serum and standard (Purified Mouse IgE, k isotype Standard, BD Pharmingen), and detection with Biotin Rat Anti-Mouse IgE at 1 μg ml−1 (R35-118, BD Pharmingen), streptavidin horseradish peroxidase at 1:1,000 (BD Pharmingen), and ABTS One Component horseradish peroxidase Microwell Substrate (SurModics, Eden Prairie, MN). IL-10 was measured in culture supernatant using paired capture and detection antibodies (BD Pharmingen).
In vitro T-cell polarization and Th2 cell survival assay. Naïve CD4+ T cells were sorted from spleens as CD4+TCRβ+CD44–CD25–Il4gfp–PI– and cultured in vitro for 3 days with 10 ng ml−1 IL-4 (R&D, Minneapolis, MN), 5 ng ml−1 IL-2 (R&D), and 10 μg ml−1 anti-IFNγ (XMG1.2, BioXcell, Minneapolis, MN) on flat-bottom 96-well plates coated with 1 μg ml−1 anti-CD3 (17A2, BioXcell) and 10 μg ml−1 anti-CD28 (37.51, BioXcell). On day 3, cells were split into round-bottom plates and supplemented with media containing 5 ng ml−1 IL-2 (R&D). On day 7, cells were counted, washed, and repolarized for another week following the same timings as week 1. Th2 cell survival assays were performed as previously described.24 Briefly, we generated Th2 cells for 7 days, as above. On day 7, Il4gfp+PI– Th2 cells, or simply PI– cells when using Il10–/– cells, were sort purified and stimulated on flat-bottom 96-well plates coated with 10 μg ml−1 of anti-CD3 (17A2, BioXcell) for 24 h. Cultures were supplemented with recombinant IL-10(10 ng ml−1) or a granzyme B inhibitor (10 μM, z-AAD-CH2Cl, Cambridge BioScience, Cambridge, UK) where indicated. PI incorporation, granzyme B expression, and total cell counts were performed after 24 h of culture.
Western blotting. Sorted naive or day 3 polarized Th2 cells were lysed in 1 × RIPA buffer (500 mM Tris HCl, pH 7.5, 150 mM NaCl, 2 mM EDTA, 0.1% SDS, 0.5% deoxycholate, and 1% Nonidet-P40) containing protein inhibitors as per the manufacturer’s instructions (Roche), 50 mM NaF, 1 mM Sodium ortho-vanadate, 100 nM Okadaic acid, and 2 mM sodium pyrophosphate tertrabasic. Protein concentration was determined using the Pierce BCA Protein Assay Kit (Life Technologies), lysates were resolved on a 10% Criterion TGX (Bio-Rad, Hercules, CA) or NuPage 4-12% Bis-Tris Gel (Life Technologies), and separated proteins were transferred onto Trans-Blot Turbo PVDF transfer (Bio-Rad) membranes. Membranes were blocked with 0.1% PBST (PBS with Tween 20) containing 5% milk (Sigma) and incubated with primary antibodies from Cell Signalling Technology (Danvers, MA): pSTAT3 (9145), pSTAT6 (9361), STAT3 (12640), and STAT6 (9362), followed by secondary (Rabbit IgG; GE Healthcare, Little Chalfont, UK) antibody in 0.1% PBST (Sigma) containing 5% milk (Sigma). Actin antibody was obtained from Santa Cruz Biotechnologies (C-11; Dallas, TX); secondary antibody for Actin was Rabbit Anti-Goat IgG (H+L) horseradish peroxidase (Southern Biotech, Birmingham, AL). Membranes were washed in 0.1% PBST and specific bound antibodies were visualized by chemiluminescence (Amersham ECL (GE Healthcare), Immobilon; (Merck Millipore, Billerica, MA)).
Quantitative real-time reverse transcription-PCR. RNA was extracted using the RNeasy Mini Kit (Qiagen, Hilden, Germany). Complementary DNA was reverse transcribed from RNA using QuantiTect Reverse Transcription Kit (Qiagen) according to the manufacturer’s instructions. Quantitative real-time reverse transcription-PCR analysis was performed using Power SYBR Green PCR master mix (Applied Biosystems, Foster City, CA) on an ABI Prism 7900HT Sequence Detection System (Applied Biosystems). Relative quantities of mRNA were determined by the comparative threshold cycle method as described by Applied Biosystems for the ABI Prism 7700/7900HT Sequence Detection Systems using the following primers: Hprt forward (Fwd): 5′-GCCCTTGACTATAATGAGTACTTCAGG-3′ and reverse (Rvs): 5′-TTCAACTTGCGCTCATCTTAGG-3′; Il10r1 Fwd: 5′-CCCATTCCTCGTCACGATCT-3′ and Rvs: 5′-TTTCCAGTGGAGGATGTGCT-3′; Il10 Fwd: 5′-ATGCTGCCTGCTCTTACTGACTG-3' and Rvs: 5′-CCCAAGTAACCCTTAAAGTCCTGC-3′; Muc5ac Fwd: 5′-CAGGACTCTCTGAAATCGTACCA-3′ and Rvs 5′-AAGGCTCGTACCACAGGGA-3′.
RNA-Seq. RNA was isolated from cells using RNeasy Mini Kit according to the manufacturer’s instructions (Qiagen). RNA-Seq libraries were constructed using the TruSeq RNA Sample Preparation Kit V2 according to the manufacturer’s instructions (Illumina, San Diego, CA). Libraries were sequenced using the HiSeq 2500 System (Illumina).The raw Illumina reads were analyzed as follows. First, the data quality was analyzed using FastQC (www.bioinformatics.babraham.ac.uk/projects/fastqc). Low-quality bases were trimmed using Trimmomatic,41 and the read pairs that passed the trimming quality filters were aligned to mm10 (Ensembl version 75) using Tophat2.42 Counts were determined using htseq_count.43 Normalization and statistical analysis was performed using edgeR.44 Normalization and statistical analysis was performed using edgeR44 with statistically significant genes (P<0.05) determined using a modified t-test with Benjamini–Hochberg multiple testing correction and a false discovery rate of <0.01.
Statistical analysis. Data sets were compared by Mann–Whitney test using GraphPad Prism (V.5.0, San Diego, CA). Differences were considered significant at *P⩽0.05.
References
Moore, K.W., O'Garra, A., de Waal Malefyt, R., Vieira, P. & Mosmann, T.R. Interleukin-10. Annu. Rev. Immunol. 11, 165–190 (1993).
Hawrylowicz, C.M. Regulatory T cells and IL-10 in allergic inflammation. J. Exp. Med. 202, 1459–1463 (2005).
Borish, L., Aarons, A., Rumbyrt, J., Cvietusa, P., Negri, J. & Wenzel, S. Interleukin-10 regulation in normal subjects and patients with asthma. J. Allergy Clin. Immunol. 97, 1288–1296 (1996).
John, M. et al. Inhaled corticosteroids increase interleukin-10 but reduce macrophage inflammatory protein-1alpha, granulocyte-macrophage colony-stimulating factor, and interferon-gamma release from alveolar macrophages in asthma. Am. J. Respir. Crit. Care Med. 157, 256–262 (1998).
Lim, S., Crawley, E., Woo, P. & Barnes, P.J. Haplotype associated with low interleukin-10 production in patients with severe asthma. Lancet 352, 113 (1998).
Wilson, M.S. et al. IL-13Ralpha2 and IL-10 coordinately suppress airway inflammation, airway-hyperreactivity, and fibrosis in mice. J. Clin. Invest. 117, 2941–2951 (2007).
Grunig, G., Corry, D.B., Leach, M.W., Seymour, B.W., Kurup, V.P. & Rennick, D.M. Interleukin-10 is a natural suppressor of cytokine production and inflammation in a murine model of allergic bronchopulmonary aspergillosis. J. Exp. Med. 185, 1089–1099 (1997).
Tournoy, K.G., Kips, J.C. & Pauwels, R.A. Endogenous interleukin-10 suppresses allergen-induced airway inflammation and nonspecific airway responsiveness. Clin. Exp. Allergy 30, 775–783 (2000).
Radinger, M., Johansson, A.K., Sitkauskiene, B., Sjostrand, M. & Lotvall, J. Eotaxin-2 regulates newly produced and CD34 airway eosinophils after allergen exposure. J. Allergy Clin. Immunol. 113, 1109–1116 (2004).
Akdis, C.A., Blesken, T., Akdis, M., Wuthrich, B. & Blaser, K. Role of interleukin 10 in specific immunotherapy. J. Clin. Invest. 102, 98–106 (1998).
Jutel, M. et al. IL-10 and TGF-beta cooperate in the regulatory T cell response to mucosal allergens in normal immunity and specific immunotherapy. Eur. J. Immunol. 33, 1205–1214 (2003).
de Waal Malefyt, R. et al. Interleukin 10 (IL-10) and viral IL-10 strongly reduce antigen-specific human T cell proliferation by diminishing the antigen-presenting capacity of monocytes via downregulation of class II major histocompatibility complex expression. J. Exp. Med. 174, 915–924 (1991).
Fiorentino, D.F. et al. IL-10 acts on the antigen-presenting cell to inhibit cytokine production by Th1 cells. J. Immunol. 146, 3444–3451 (1991).
Kamanaka, M. et al. Memory/effector (CD45RB(lo)) CD4 T cells are controlled directly by IL-10 and cause IL-22-dependent intestinal pathology. J. Exp. Med. 208, 1027–1040 (2011).
Huber, S. et al. Th17 cells express interleukin-10 receptor and are controlled by Foxp3(-) and Foxp3+ regulatory CD4+ T cells in an interleukin-10-dependent manner. Immunity 34, 554–565 (2011).
Chaudhry, A. et al. Interleukin-10 signaling in regulatory T cells is required for suppression of Th17 cell-mediated inflammation. Immunity 34, 566–578 (2011).
Schandene, L. et al. B7/CD28-dependent IL-5 production by human resting T cells is inhibited by IL-10. J. Immunol. 152, 4368–4374 (1994).
Barrat, F.J. et al. In vitro generation of interleukin 10-producing regulatory CD4(+) T cells is induced by immunosuppressive drugs and inhibited by T helper type 1 (Th1)- and Th2-inducing cytokines. J. Exp. Med. 195, 603–616 (2002).
Pils, M.C. et al. Monocytes/macrophages and/or neutrophils are the target of IL-10 in the LPS endotoxemia model. Eur. J. Immunol. 40, 443–448 (2010).
Wang, S.B., Deng, Y.Q., Ren, J., Xiao, B.K., Liu, Z. & Tao, Z.Z. Exogenous interleukin-10 alleviates allergic inflammation but inhibits local interleukin-10 expression in a mouse allergic rhinitis model. BMC Immunol. 15, 9 (2014).
Finbloom, D.S. & Winestock, K.D. IL-10 induces the tyrosine phosphorylation of tyk2 and Jak1 and the differential assembly of STAT1 alpha and STAT3 complexes in human T cells and monocytes. J. Immunol. 155, 1079–1090 (1995).
Mohrs, M., Shinkai, K., Mohrs, K. & Locksley, R.M. Analysis of type 2 immunity in vivo with a bicistronic IL-4 reporter. Immunity 15, 303–311 (2001).
Maynard, C.L. et al. Regulatory T cells expressing interleukin 10 develop from Foxp3+ and Foxp3- precursor cells in the absence of interleukin 10. Nat. Immunol. 8, 931–941 (2007).
Devadas, S. et al. Granzyme B is critical for T cell receptor-induced cell death of type 2 helper T cells. Immunity 25, 237–247 (2006).
Ichiba, M., Nakajima, K., Yamanaka, Y., Kiuchi, N. & Hirano, T. Autoregulation of the Stat3 gene through cooperation with a cAMP-responsive element-binding protein. J. Biol. Chem. 273, 6132–6138 (1998).
Stritesky, G.L. et al. The transcription factor STAT3 is required for T helper 2 cell development. Immunity 34, 39–49 (2011).
Zhu, J. et al. Conditional deletion of Gata3 shows its essential function in T(H)1-T(H)2 responses. Nat. Immunol. 5, 1157–1165 (2004).
Shoemaker, J., Saraiva, M. & O'Garra, A. GATA-3 directly remodels the IL-10 locus independently of IL-4 in CD4+ T cells. J. Immunol. 176, 3470–3479 (2006).
Chang, H.D. et al. Expression of IL-10 in Th memory lymphocytes is conditional on IL-12 or IL-4, unless the IL-10 gene is imprinted by GATA-3. Eur. J. Immunol. 37, 807–817 (2007).
Guo, L., Huang, Y., Chen, X., Hu-Li, J., Urban, J.F. Jr. & Paul, W.E. Innate immunological function of T2 cells in vivo. Nat. Immunol. 16, 1051–9 (2015).
Bohm, L. et al. IL-10 and regulatory T cells cooperate in allergen-specific immunotherapy to ameliorate allergic asthma. J. Immunol. 194, 887–897 (2015).
Stanic, B. et al. IL-10-overexpressing B cells regulate innate and adaptive immune responses. J. Allergy Clin. Immunol. 135, 771–780.e8 (2015).
Mumm, J.B. et al. IL-10 elicits IFNgamma-dependent tumor immune surveillance. Cancer Cell 20, 781–796 (2011).
Altin, J.A., Goodnow, C.C. & Cook, M.C. IL-10+ CTLA-4+ Th2 inhibitory cells form in a Foxp3-independent, IL-2-dependent manner from Th2 effectors during chronic inflammation. J. Immunol. 188, 5478–5488 (2012).
Xu, X. et al. Activation-induced T helper cell death contributes to Th1/Th2 polarization following murine Schistosoma japonicum infection. J. Biomed. Biotechnol. 2010, 202397 (2010).
van de Berg, P.J., van Leeuwen, E.M., ten Berge, I.J. & van Lier, R. Cytotoxic human CD4(+) T cells. Curr. Opin. Immunol. 20, 339–343 (2008).
McHugh, R.S. et al. CD4(+)CD25(+) immunoregulatory T cells: gene expression analysis reveals a functional role for the glucocorticoid-induced TNF receptor. Immunity 16, 311–323 (2002).
Steidler, L. et al. Treatment of murine colitis by Lactococcus lactis secreting interleukin-10. Science 289, 1352–1355 (2000).
Mohrs, K., Wakil, A.E., Killeen, N., Locksley, R.M. & Mohrs, M. A two-step process for cytokine production revealed by IL-4 dual-reporter mice. Immunity 23, 419–429 (2005).
Wan, Y.S.Y. & Flavell, R.A. Identifying Foxp3-expressing suppressor T cells with a bicistronic reporter. Proc. Natl. Acad. Sci. USA 102, 5126–5131 (2005).
Bolger, A.M., Lohse, M. & Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120 (2014).
Kim, D., Pertea, G., Trapnell, C., Pimentel, H., Kelley, R. & Salzberg, S.L. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 14, R36 (2013).
Anders, S., Theodor Pyl, P. & Huber, W. HTSeq — A Python framework to work with high-throughput sequencing data. Bioinformatics 131, 166–169 (2015).
Robinson, M.D., McCarthy, D.J. & Smyth, G.K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2010).
Acknowledgements
We are indebted to Anne O’Garra for providing critical comments and advice. We also thank The Francis Crick Institute Flow Cytometry Facility, specifically Bhavik Patel, Graham Preece, and Wayne Turnbull, for the provision of cell sorting services in the production of this work. We are indebted to The Francis Crick Institute Procedural Service Section for production of GA lines and Biological Services, especially Trisha Norton, Keith Williams, and Adebambo Adekoya for animal husbandry and technical support. We thank the Systems Biology Department, in particular Abdul Sessay, Harsha Jani, Deb Jackson, and Leena Bhaw-Rosun, for help with RNA-Seq processing. We also thank Radma Mahmood and Radhika Anand for preparation of histology samples. The majority of this work was supported by the Medical Research Council (MRC File Reference number MC_UP_A253_1028) before we became The Francis Crick Institute on 1 April 2015.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declared no conflict of interest.
Additional information
SUPPLEMENTARY MATERIAL is linked to the online version of the paper
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Coomes, S., Kannan, Y., Pelly, V. et al. CD4+ Th2 cells are directly regulated by IL-10 during allergic airway inflammation. Mucosal Immunol 10, 150–161 (2017). https://doi.org/10.1038/mi.2016.47
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mi.2016.47
This article is cited by
-
Anti-CD20 treatment attenuates Th2 cell responses: implications for the role of lung follicular mature B cells in the asthmatic mice
Inflammation Research (2024)
-
Mesenchymal stem cells overexpressing interleukin-10 prevent allergic airway inflammation
Stem Cell Research & Therapy (2023)
-
B cell-derived IL-10 promotes the resolution of lipopolysaccharide-induced acute lung injury
Cell Death & Disease (2023)
-
B-cell-derived IL-10 promotes allergic sensitization in asthma regulated by Bcl-3
Cellular & Molecular Immunology (2023)
-
Interleukin-10 multitasking in allergic airway inflammation
Cellular & Molecular Immunology (2023)
