
Abstract
Acute respiratory distress syndrome (ARDS) causes significant morbidity and mortality each year. There is a paucity of information regarding the mechanisms necessary for ARDS resolution. Foxp3+ regulatory T cells (Foxp3+ Treg cells) have been shown to be an important determinant of resolution in an experimental model of lung injury. We demonstrate that intratracheal delivery of endotoxin (lipopolysaccharide) elicits alveolar epithelial damage from which the epithelium undergoes proliferation and repair. Epithelial proliferation coincided with an increase in Foxp3+ Treg cells in the lung during the course of resolution. To dissect the role that Foxp3+ Treg cells exert on epithelial proliferation, we depleted Foxp3+ Treg cells, which led to decreased alveolar epithelial proliferation and delayed lung injury recovery. Furthermore, antibody-mediated blockade of CD103, an integrin, which binds to epithelial expressed E-cadherin decreased Foxp3+ Treg numbers and decreased rates of epithelial proliferation after injury. In a non-inflammatory model of regenerative alveologenesis, left lung pneumonectomy, we found that Foxp3+ Treg cells enhanced epithelial proliferation. Moreover, Foxp3+ Treg cells co-cultured with primary type II alveolar cells (AT2) directly increased AT2 cell proliferation in a CD103-dependent manner. These studies provide evidence of a new and integral role for Foxp3+ Treg cells in repair of the lung epithelium.
Similar content being viewed by others

Introduction
Acute respiratory distress syndrome (ARDS) is characterized by rapid-onset bilateral pulmonary infiltrates hallmarked by an inflammatory response with neutrophil accumulation, increase in alveolar fluid, and pro-inflammatory cytokine release.1 This syndrome has significant morbidity and mortality, with in-hospital mortality as high as 44%, and accounts for nearly 200,000 hospitalizations and 75,000 deaths each year in the United States.2 Despite years of research, the only treatment for ARDS demonstrated to improve outcomes are supportive.3, 4
Repair of the alveolar epithelium after acute lung injury (ALI) is necessary to restore homeostasis, and current views have proposed that the immune system may have an important role in protecting epithelial surfaces by enhancing barrier function and promoting repair.5, 6 In acute or chronic injury, the failure to regenerate the lung epithelium has a role in such processes as ALI, pneumonia, pulmonary fibrosis, chronic obstructive pulmonary disease, and aging.5 The underlying mechanisms involved in epithelial repair remain largely unknown. Previous work demonstrates a central role for Foxp3+ regulatory T cells (Foxp3+ Treg cells) in the resolution of experimental lung ALI by modulating pro-inflammatory alveolar macrophages and reducing fibroproliferation by decreasing fibrocyte recruitment.7, 8 Moreover, Foxp3+ Treg cells have been shown to increase in the bronchoalveolar lavage (BAL) fluid of patients with ARDS.8
Foxp3+ Treg cells are a distinct population of lymphocytes, which express the transcription factor forkhead homeobox protein-3 (Foxp3).9, 10 This T-cell subset has been demonstrated to suppress or downregulate immune responses in allergic and autoimmune diseases, as well as in cancer biology.11 The mechanisms involved in Foxp3+ Treg cell suppressor activity depend on the context of the response, and include contact-dependent inhibitory cell surface receptors (CTLA-4 and LAG-3), secretion of inhibitory cytokines (interleukin-10 and transforming growth factor-β, competition for growth factors (interleukin-2), and direct lysis (granzymes).12, 13
Prior work has highlighted an important role for Foxp3+ Treg cells in the resolution of experimental lung injury;8, 14 however, pro-resolution mechanisms still remain to be explored. In this study, multicolor flow cytometry was used to identify epithelial populations in the distal lung along with their rates of proliferation during resolution. Using an established model of experimental ALI, intratracheal lipopolysaccharide (IT LPS), we identified a function of Foxp3+ Treg cells in augmenting the proliferation of the epithelium during ALI resolution. In addition, CD103 (an integrin molecule that binds E-cadherin) blockade decreases Foxp3+ Treg cell abundance and alveolar epithelial proliferation during resolution from injury. To determine whether these findings extended to a non-overt inflammatory model of lung growth, a left unilateral pneumonectomy (PNX) model in mice was employed. The left lung is surgically removed eliciting a compensatory response in the remaining right lung, which undergoes a process described as regenerative alveologenesis.15 Foxp3+ Treg cell numbers increased in the alveolar and total lung compartments 7 days post PNX, and mice lacking mature lymphocytes (recombinase-activating gene-1-deficient (Rag-1−/−)) or Foxp3+ Treg-depleted animals (Foxp3DTR) had decreased rates of epithelial proliferation. Furthermore, in vitro co-culture studies demonstrated that proliferation of primary type II alveolar epithelial (AT2) cells was enhanced when cultured with Foxp3+ Treg cells, suggesting a direct effect on lung epithelial proliferation. These studies provide evidence of a new and integral role for Foxp3+ Treg cells in repair of the epithelial cells during inflammatory and non-inflammatory models of lung injury and growth.
Results
Flow cytometry method for identification of alveolar epithelial cells
Multicolor flow cytometry was used to identify specific subpopulations of the alveolar epithelium during and after experimental lung injury from live, single lung cell suspensions obtained similar to previous methods.16, 17, 18 To identify the epithelial cell population in the single-cell suspensions, surface markers CD31 (endothelial lineage) and CD45 (hematopoietic lineage) were used to exclude these two lineages (Lin−) when gated against a pan-epithelial surface marker CD326 (epithelial cell adhesion molecule) in a manner similar to previous reports16, 19 (see Supplementary Figure E1A online). Type I alveolar epithelial (AT1) cells were further delineated in the Lin−CD326+ subpopulation by employing an antibody against a well-characterized marker for AT1 cells, T1α (also known as RT140 or podoplanin).20, 21 Cells gated by this method also contained markers for airway cells such as club cells (formerly called Clara cells) and were identified through the expression of club cell secretory protein (CC10). AT2 cells were identified by surface expression of major histocompatibility class II (MHC II; anti-I-A/I-E antibody), which has previously been reported to be constitutively expressed on AT2 cells.22 Therefore, the Lin−CD326+ population could be further divided into Lin−CD326+MHCII+T1α− (AT2) or Lin−CD326+T1α+ (AT1/club cells) (Supplementary Figure E1A). Additional detail on the method for identification and confirmation of lineage is provided in an online data supplement (Supplementary Figure E1B). We used this scheme to identify changes in alveolar epithelial cell populations in response to experimental lung injury or PNX.
Lung epithelial cells proliferate after LPS-induced ALI
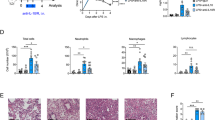
Following administration of IT LPS (3 mg kg−1) in wild-type (WT) (C57BL/6) mice, there is an influx of inflammatory cells and peak injury occurs at day 3 as measured by histology (Figure 1a), BAL protein (Figure 1b), and BAL cell count (Figure 1c). These parameters of lung injury return toward uninjured levels by days 7 to 10 (Figure 1a–c). The receptor for advanced glycation end products (RAGE) is expressed on the basolateral surface of AT1 cells and increased levels in the BAL fluid indicates direct alveolar epithelial damage in lung injury models.23 After IT LPS administration, BAL-soluble RAGE levels increased at day 1, with a return towards baseline levels by day 3 (Figure 1d).

Lung epithelial cells are more proliferative after lipopolysaccharide (LPS)-induced acute lung injury (ALI). Epithelial proliferation levels peak during ALI resolution. Wild-type (WT) mice (n=8–14 per group per time point) were challenged with intratracheal (IT) LPS. (a) Representative lung sections were stained with hematoxylin and eosin to reveal morphological changes at time points after IT LPS. Original magnification, × 40. (b–d) Bronchoalveolar lavage (BAL) total protein (b), BAL total cell counts (c), and BAL soluble receptor for advanced glycation end products (RAGE) measured by enzyme-linked immunosorbent assay (d) were determined in WT mice controls or after treatment with LPS. *P<0.001 versus control by Mann–Whitney test. (e–g) Total lung cell count (e), total CD326+ or E-cadherin+ cell counts (f), and percentage of proliferating (ki-67+) CD326+ or E-cadherin+ cells (g) were determined by flow cytometry in WT mice controls or after treatment with IT LPS. #P<0.05 versus control by Mann–Whitney test. (h) Alveolar regulatory T (Treg) cell (CD4+CD25+Foxp3+) numbers at baseline and after injury in WT mice. (i) Total Foxp3+ Treg numbers (GFP+ cells) in single-cell suspensions of Foxp3gfp mice at baseline and 7 days after treatment with IT water or LPS. (n=7 per group). ^P<0.01 versus control by Mann–Whitney test. P-values determined by Mann–Whitney test.
IT LPS lead to more than a two-fold increase in the total number of lung cells at peak injury (day 3), with a return to baseline by days 7 and 10 (Figure 1e). Inflammatory cells such as neutrophils and macrophages made up a large portion of the influx of cells. The number of Lin−CD326+ epithelial cells significantly decreased at day 3 after IT LPS administration (Figure 1f). Labeling of E-cadherin (CD324), another pan-epithelial marker, demonstrated similar percentages and numbers of epithelial cells as compared with Lin−CD326+ cells, with significant overlap (Figure 1f), irrespective of injury phase (data not shown).24 The percentage of Lin−CD326+ or Lin−E-cadherin+ epithelial cells staining for ki-67, a marker of proliferation, increased from a baseline (for Lin−CD326+) of 1.37±0.39% (mean±s.d.; n=11) at control conditions to an average peak proliferation rate of 15.90±6.80% (mean±s.d.; n=10) 7 days after LPS-induced injury (Figure 1g). The predominant proliferating alveolar epithelial cell type was AT2 as measured by Lin−CD326+MHCII+T1α− ki-67+ cells (Supplementary Figure E2A-B). These studies indicate that IT LPS induces epithelial loss and damage at peak lung injury and subsequent proliferation of alveolar epithelial cells during time points of ALI resolution.
Alveolar and interstitial Foxp3+ Treg cells increase after ALI
The timing of peak alveolar Foxp3+ Treg cells correlates with maximal epithelial proliferation (Figure 1h). To further understand the potential contribution of Foxp3+ Treg cells in the epithelial response after injury, we performed flow cytometry on both the alveolar compartment (BAL) and single-cell lung suspensions (whole lung) to determine potential changes in Foxp3+ Treg cell numbers after injury. We used the Foxp3gfp reporter mouse that expresses a N-terminal green fluorescent protein (GFP)–Foxp3 fusion protein to identify Foxp3+ Treg cells in our ALI model.25 Foxp3gfp and WT mice had similar increases in the numbers of Foxp3+ Treg cells and rates of epithelial proliferation 7 days post IT LPS administration (Supplementary Figure E3A-D). We also detected a significant increase in Foxp3+ Treg cells in the lung compartment after LPS injury (Figure 1i) in Foxp3gfp mice despite no difference in total lung cell numbers between control and mice 7 days post injury. (Supplementary Figure E3C). These observations show that Foxp3+ Treg cell numbers increase in the lung during experimental ALI resolution.
Epithelial proliferation during ALI resolution is impaired in Foxp3+ Treg cell-depleted mice
To determine whether Foxp3+ Treg cells affect epithelial proliferation during resolution from LPS-induced injury we used a transgenic Foxp3DTR mouse. Foxp3DTR mice express the human diphtheria toxin receptor (DTR) along with GFP, whose genes have been inserted into the 3′- untranslated region of the Foxp3 locus.26 These mice allow specific elimination of Foxp3+ Treg cells in vivo through intraperitoneal (i.p.) administration of diphtheria toxin (DT).26, 27
We examined the role of Foxp3+ Treg cells in lung cell numbers and rates of proliferation after LPS-induced injury and DT administration in Foxp3DTR mice (Figure 2a). Foxp3+ Treg-depleted mice have persistent histological lung injury 7 days after IT LPS administration (Figure 2b) with increased BAL total protein (Supplementary Figure E4A). The total number of lung cells was not statistically different 7 day post IT LPS administration (Figure 2c). Foxp3+ Treg-depleted mice had decreased total CD326+ cells compared with similarly treated Foxp3gfp mice (Figure 2d, P=0.0535, column 3 versus column 4). Injured Foxp3+ Treg-depleted mice had lower absolute numbers of proliferating CD326+ cells (Figure 2e) along with a lower percentage of proliferating CD326+ cells compared with their controls (Foxp3gfp mice administered both LPS and DT) (Figure 2f). Similar results were detected with E-cadherin (data not shown). Greater than 90% Foxp3+ Treg-cell depletion was obtained as evaluated by flow cytometry of the spleen (Supplementary Figure E4B). I.p. administration of DT alone in the absence of IT LPS had no effect on the rate of epithelial proliferation (Figure 2e and f) or markers of lung inflammation as measured by BAL protein (Supplementary Figure E4A).26 In summary, injured mice lacking Foxp3+ Treg cells have both impairment in resolution of lung injury and decreased rates of epithelial proliferation.

Epithelial proliferation during acute lung injury (ALI) resolution is impaired in regulatory T (Treg) cell-depleted mice. Treg-depleted mice have decreased levels of epithelial proliferation. (a) Foxp3gfp or Foxp3DTR mice (n=5–9 per group) were challenged with intratracheal lipopolysaccharide (IT LPS) (day 0) and/or diphtheria toxin (DT; 50 or 10 μg kg–1) administered at day −2, −1, 1, 3, and 5 and collected at day 7 post LPS. (b) Representative lung sections were stained with hematoxylin and eosin to reveal morphologic changes 7 days after IT LPS±DT. Original magnification, × 40. (c–f) Total lung cell count (c), total CD326+ cell numbers (d), proliferating epithelial cell (CD326+ki-67+) numbers (e), and percentage of proliferating CD326+ cells compared with total CD326+ cells (f) were determined by flow cytometry. P-values determined by Mann–Whitney test.
Epithelial proliferation after ALI is abrogated with blockade of CD103
One mechanism by which lymphocytes may localize and interact with epithelial surfaces is through their expression of the integrin αEβ7.28, 29, 30 Integrin αE (CD103) binds integrin β7 to form a heterodimeric molecule whose only known ligand is E-cadherin expressed on epithelial cells.29, 30 The percentage of Foxp3+ Treg cells expressing CD103 increased 3 days after IT LPS administration when compared with baseline levels (Figure 3a). To examine the role of CD103, we use antibody-mediated blockade of CD103 delivered to WT mice (i.p.) on days 0, 1, 3, and 5 after IT LPS administration (Figure 3b) and assessed epithelial proliferation at day 7. CD103 blockade significantly decreased Foxp3+ Treg cells and decreased both the percentage and number of proliferating epithelial cells (CD326+ ki67+) in the lung at 7 days after injury when compared with isotype antibody-treated animals (Figure 3c–e). Lung Foxp3+ Treg cell total numbers directly correlated with greater epithelial cell proliferation (Figure 3f). There was no statistical difference in injury parameters in mice administered CD103 blockade versus antibody control (see Supplementary Figures E5A-E). These results support a role for Foxp3+ Treg CD103 in modulating lung epithelial proliferation and further show a strong correlation between Foxp3+ Treg numbers in the lung and increased rates of epithelial proliferation after injury.

Regulatory T (Treg) cell number and epithelial proliferation after acute lung injury (ALI) is abrogated with antibody blockade of CD103. CD103 blockade decreased lung Foxp3+ Treg number and subsequent epithelial proliferation after ALI. (a) Wild-type (WT) mice (n=10–12 per group per time point) were challenged with intratracheal lipopolysaccharide (IT LPS) and the number of Foxp3+ Treg cells in the lung cell suspensions were identified by flow cytometry along with the percentage of Foxp3+ cells that co-expressed CD103 (n=7–9 per group per time point). *P<0.05; ^P<0.005 versus control by Mann–Whitney test. (b–f) WT mice (n=9 per group) were challenged with IT LPS along with administration of either an anti-CD103 antibody (150 μg) or isotype control (150 μg rat IgG) at days 0, 1, 3, and 5 post IT LPS administration (b). The number of Foxp3+ cells in the lung cell suspensions were determined by flow cytometry (c) along with the total number (d) and percentage (e) of proliferating CD326+ cells (CD326+ki-67+ cells) cells. (f) Total lung Foxp3+ cell number positively correlated with CD326+ki-67+ epithelial cell number (Pearson’s r=0.8730, R2=0.7621, P<0.0001 using linear regression). #P=0.0939; *P<0.05 by Mann—Whitney test.
Transfer of CD103−/− Foxp3+ Treg cells into lymphopenic mice fails to augment epithelial proliferation
CD103 expression is not limited to Foxp3+ Treg cells but is also found on other lymphocyte subsets such as intraepithelial lymphocytes and Thy-1+ dendritic epidermal T cells along with subsets of dendritic cells.28, 30 To determine the role of CD103 on Foxp3+ Treg cells in mediating epithelial proliferation we isolated CD4+CD25− or CD4+CD25+ cells from the spleens of WT or CD103−/− mice and performed adoptive transfers (AT) into Rag-1−/− mice exposed to IT LPS, followed by collection at 7 days.8 We have shown that lymphocyte-deficient Rag-1−/− mice have impaired lung injury resolution despite similar initial and peak parameters of lung injury following administration of IT LPS compared with WT mice.8
Surviving Rag-1−/− mice had decreased numbers of CD326+ cells when compared with WT mice despite a similar number of total lung cells (Supplementary Figure E6A-B). AT of WT Treg cells (CD4+CD25+ lymphocytes) into Rag-1−/− mice restored the percentage of proliferating CD326+ ki67+ cells back to WT levels. In contrast, AT of WT CD4+CD25− or CD103−/−CD4+CD25+ cells back to Rag-1−/− mice did not increase CD326+ epithelial proliferation (Figure 4a). Among the AT groups, Rag-1−/− mice that received WT CD4+CD25+ cells had significantly more Foxp3+ cells than Rag-1−/− mice that received CD103−/− CD4+CD25+ cells (Figure 4b). Furthermore, AT of WT CD4+CD25+ cells but not CD4+CD25− or CD103−/− CD4+CD25+ cells into Rag-1−/− mice led to restoration of barrier permeability as measured by BAL total protein levels (Supplementary Figure E6C). No significant difference was detected in BAL total cell count between AT of WT CD4+CD25+ or CD4+CD25+CD103−/− cells into Rag-1−/− mice (Supplementary Figure E6D); however, AT of WT CD4+CD25+ cells but not CD4+CD25− or CD4+CD25+CD103−/− cells into Rag-1−/− mice led to restoration of histopathologic resolution 7 days after LPS-induced injury (Supplementary Figure E6E). These results reinforce the role for Foxp3+ Treg cells and its integrin CD103 in the proliferation of alveolar epithelial cells after injury and demonstrate a partial complementation in inflammatory resolution of the alveolar epithelium after acute injury.

Adoptive transfer (AT) of regulatory T cells (Tregs) augments epithelial proliferation after acute lung injury (ALI) in Rag-1−/− mice. AT of wild-type (WT) Tregs augments epithelial proliferation after ALI in Rag-1−/− mice. Rag-1−/− mice were challenged with intratracheal lipopolysaccharide (IT LPS) and 1 h afterward received 1 × 106 WT CD4+CD25+, WT CD4+CD25− or CD103−/−CD4+CD25+ cells and lungs collected at day 7 post LPS administration. (a) Percentage of proliferating CD326+ki-67+ epithelial cells at day 7 post LPS in WT mice or Rag-1−/− mice after infusion of designated lymphocytes subsets (n=6–12 per group). (b) Total number of Foxp3+ cells in the lungs of WT mice or Rag-1−/− mice after infusion of designated lymphocytes subsets (n=4–7 per group). P-values determined by Student’s t-test.
Foxp3+ Treg cells increase in the right lung after left lung PNX
We sought to determine whether Foxp3+ Treg cells can induce augmentation of epithelial proliferation independent on their ability to dampen alveolar inflammation. We employed a left lung PNX murine model characterized by contralateral lung growth without significant inflammation.15, 31
After PNX of the left lung, we observed an increase in the right lung cellularity by histology compared with sham surgery (L thoracotomy) (Figure 5a). Total Foxp3+ Treg cells increased in the BAL and in the right lung of contralateral PNX-treated mice compared with control or sham surgery of WT (Figure 5b and c). Peak levels of epithelial proliferation occurred at day 7 post PNX (data not shown). The increase in Foxp3+ Treg cells also correlated with the peak level of CD326+ epithelial proliferation (Figure 5f; column 1 and 2). There was no difference in markers of inflammation, including BAL protein or BAL cell count between WT mice undergoing left PNX versus sham surgery (data not shown). These observations indicate that Foxp3+ Treg cell numbers increase in the remaining lung post PNX with a concomitant increase in epithelial proliferation.

Regulatory T cells (Tregs) augment epithelial proliferation in mice after pneumonectomy (PNX). Wild-type (WT), Rag-1−/−, Foxp3gfp, and Foxp3DTR mice underwent left lung pneumonectomy. Foxp3gfp and Foxp3DTR mice undergoing PNX were also administered diphtheria toxin (DT) at the time of PNX and at days 2, 4, and 6 post PNX. (a) Representative lung sections were stained with hematoxylin and eosin to reveal morphological changes 7 days post procedure in WT sham, WT PNX, and Foxp3DTR mice (PNX and DT). Original magnification, × 40 and × 100. (b and c) WT mice (n=5 per group) alveolar (b) and lung (c) Foxp3+ Treg number at baseline and at 7 days post sham or PNX. *P<0.05 versus control/sham by 1 way analysis of variance. (d–f) Total lung cell count (d), CD326+ cell numbers (e), and percentage of proliferating epithelial cells (CD326+ki-67+) number (f) were determined by flow cytometry 7 days after sham procedure or PNX (n=6–13 per group). P-values determined by Mann–Whitney test.
Foxp3+ Treg cells augment epithelial proliferation after PNX
To further examine whether Foxp3+ Treg cells can modulate epithelial proliferation in a non-inflammatory model of lung growth, Rag-1−/−, Foxp3gfp, and Foxp3DTR mice underwent left lung PNX. Subsets of Foxp3gfp and Foxp3DTR mice were also administered DT at the time of PNX and again at days 2, 4, and 6 post PNX. The increase in the number of cells seen by histology (Figure 5a) was also determined in the prepared single-cell lung suspensions. No statistical difference in total lung cells was detected between WT sham and WT PNX (Figure 5d; column 1 and 2). There was an increase in total lung cells between Rag-1−/− sham and Rag-1−/− PNX, and between Foxp3DTR and Foxp3gfp mice undergoing PNX (Figure 5d). There was a significant difference in total CD326+ cell number between Foxp3gfp and Foxp3DTR mice undergoing PNX and administered DT (Figure 5e; columns 5 and 6). Mice lacking lymphocytes (Rag-1−/−) or depleted of Foxp3+ Treg cells (Foxp3DTR) but not controls (Foxp3gfp) had a 50% reduction in epithelial proliferation when compared with the WT mice 7 days after PNX (Figure 5f). Similar to the role of Foxp3+ Treg in the IT LPS inflammatory model, Foxp3+ Treg cells enhanced epithelial proliferation in a non-inflammatory model of alveolar growth.
Foxp3+ Treg cells enhance proliferation of AT2 cells in vitro
To determine a potential direct effect of Foxp3+ Treg cells in modifying epithelial proliferation, AT2 cells were grown in co-culture experiments with specific lymphocytes subsets (1:5 ratio of lymphocytes: AT2). This ratio was chosen as previous reports have estimated that there are ∼10–20 intraepithelial lymphocytes per 100 bronchial epithelial cells.32 AT2 cells co-cultured for 24 h with CD4+CD25+ (Foxp3+ Treg) cells had increased rates of proliferation when compared with those cultured with CD4+CD25− lymphocytes (Figure 6). The augmentation of proliferation by CD4+CD25+ cells continued when the two cells types were separated by a transwell insert, demonstrating that the effect has contact independence (Figure 6). When CD103-blocking antibody was added to the co-cultures of AT2 and Foxp3+ Treg cells, epithelial proliferation rates decreased to control levels (Figure 6). When we extended the co-cultures out to 72–96 h, a greater number of AT2 cells cultured with CD4+CD25+ cells had continued expression of surfactant protein-C as demonstrated by expression of GFP through a surfactant protein-C promoter, and decreased levels of T1α expression by imaging (Supplementary Figure E7). These data demonstrate that Foxp3+ Treg cells directly increase primary AT2 proliferation and this effect is in part CD103 dependent.

Regulatory T cells (Tregs) enhance proliferation of type II alveolar cells (AT2) in vitro. Primary AT2 cells were isolated by sorting GFP+ cells from SP-CGFP mice and co-cultured with lymphocytes subsets (wild-type (WT) CD4+CD25+ and WT CD4+CD25−) at a lymphocyte: AT2 ratio of 1:5, n≥5 for each condition. Fold increase (compared with media alone) in proliferation (CD326+ki-67+) was determined after 24 h of co-culture of AT2 cells with either CD4+CD25−, CD4+CD25+, CD4+CD25+ plus addition of CD103 blocking antibody, or CD4+CD25+ separated from AT2 cells through a Transwell insert. P-values determined by Mann–Whitney test.
Discussion
In this report we identify that Foxp3+ Treg cells augment the rate of epithelial proliferation in an experimental model of lung injury and also in a compensatory lung growth model (PNX). Lymphocyte and, specifically, Foxp3+ Treg cell effects on alveolar epithelial proliferation after ALI have not been previously described. Prior work has demonstrated an integral role for Foxp3+ Treg cells during LPS-induced lung inflammation resolution and that these cells exhibit suppressive function mainly on pro-inflammatory alveolar macrophages and neutrophils.8, 14, 33 In this study we use both a depletive method (Foxp3DTR mice) and an additive method (AT of lymphocyte subsets into Rag-1−/− mice) to determine the effect of Foxp3+ Treg cells on proliferation after injury. Furthermore, our PNX findings reinforce that Foxp3+ Treg cells exert an effect on the lung epithelium in a non-inflammatory context. Herein, we also show that the number of Foxp3+ Treg cells increases in the whole lung after both IT LPS and PNX. The extent to which the increase in total number represents proliferation, retention, recruitment, or a combination of all three is still unknown; however, results from this study suggest retention through Foxp3+ Treg cells expression of CD103 may have a role, and prior reports suggest proliferation and recruitment.8 Probably, all three mechanisms have a role and the interplay with other immune cells such as macrophages and dendritic cells promoting development of Foxp3+ Treg cells are likely involved.34
The LPS model was chosen, as it has been widely characterized and elicits reproducible alveolar epithelial damage.23, 35 One limitation of the LPS model is that in the absence of lymphocytes (Rag-1−/− mice) or specifically Foxp3+ Treg cells (Foxp3DTR mice depleted of Foxp3+ cells) there is a sustained inflammatory response that may directly or indirectly dampen the epithelial reparative process. Persistent injury in the absence of Foxp3+ Treg cells led us to explore our findings into the PNX model to better determine Foxp3+ Treg cell effects on lung epithelial proliferation independent of sustained inflammation.
We identified specific lung epithelial subpopulations during resolution of lung injury by using multicolor flow cytometry. The cytometric scheme presented allows for identification of alveolar epithelial cells and builds on the previous literature.16, 17, 19 In this report we use MHC II molecules to identify AT2 cells within the CD326+ population. MHC II has been previously described to be constitutively expressed on AT2 cells.22, 36, 37 Lee et al.19, 36 published the construction of a transgenic mouse that expresses GPF through an SP-C promoter and demonstrated that CD74 can be used to identify AT2 cells. CD74 or invariant chain is part of the MHC II system.36 Additional work is needed to better identify subsets; however, this methodology allows for identification without the need for a transgenic report strain for identification of a specific epithelial population.
Through flow cytometry we detected a significant decrease in total CD326+ or E-cadherin+ epithelial cells at peak levels of injury after IT LPS administration. This decrease in cell number could be due to damage elicited by LPS as confirmed by elevated levels of soluble RAGE after injury.23 Other possibilities could include loss of cellular expression of these markers through enzymatic cleavage, degradation, proliferation, or expansion of a specific progenitor population with altered or lack of expression of these epithelial markers at specific times during the reparative process. Peak proliferation of the epithelium occurs 7 days after IT LPS administration, which also corresponded to the peaked increase in Foxp3+ Treg cells. Depletion of Foxp3+ Treg cells throughout the injury course correlated with lower numbers of total CD326+ epithelial cells, likely due to persistent inflammation and cellular damage and the remaining CD326+ epithelial cells were less proliferative when compared with controls.
The mechanisms underlying the pro-proliferative effects on epithelium regeneration are complex and involve progenitor populations, signaling pathways, and interaction with the extracellular matrix.38 The paradigm for alveolar epithelial repair has been that AT1 cells are susceptible to damage and AT2 cells migrate into and proliferate at sites of injury.39 AT2 cells undergo hyperplasia and then differentiate into AT1 cells to reform an intact functional epithelial barrier.40 This report supports that the presence of Foxp3+ Treg cells modulates proliferation of the epithelium after injury. Interestingly, earlier studies in cultured human fetal colon tissue demonstrated that pokeweed mitogen or anti-CD3+ activated lamina propria CD3+ cells underwent induced expression of CD25+, and this T-cell activation increased the rate of epithelial proliferation in the crypts of Lieberkühn.41 This study was performed before identification of the transcription factor Foxp3, but the results support that immunological factors can exert effects on epithelial turnover and differentiation.41, 42 To our knowledge this is the first report demonstrating a role for Foxp3+ Treg cells exerting an effect on epithelial proliferation in general and specifically the lung. A previous study found that intraepithelial γδ T cells (dendritic epidermal T cells) induced epithelial cell proliferation in a murine keratinocyte cell line (Balb/MK) through production of keratinocyte growth factor.43 Foxp3+ Treg cells may also exhibit similar effects. The effects may be organ or tissue specific, and the increased rates of epithelial turnover in certain tissues such as the gastrointestinal tract or skin may not be as influenced by Foxp3+ Treg cells as the lung epithelium, and recent work demonstrate that Foxp3+CD4+ Treg cells promote muscle repair after injury.44 Future work examining Foxp3+ Treg cells’ effects in other models of injury (naphthalene, bleomycin, or live bacterial), and in respect to progenitor populations may provide a better understanding of the Foxp3+ Treg cell mechanisms involved in epithelial repair, and are the subject of ongoing investigation.
Foxp3+ Treg cells have been previously demonstrated to migrate to sites of infection, inflammation, and tumor microenvironments.45 The αE (CD103) chain is part of the integrin αEβ7, which is well described and may function to localize certain immune cells such as dendritic cells and lymphocyte subsets to epithelial surfaces through binding to E-cadherin.29 Little is known about the immune regulation of CD103 since it was first described, and CD103 expression is found largely on lymphocytes at epithelial sites.46, 47 Furthermore, CD103 expression is induced by transforming growth factor-β1 and may induce integrin αEβ7 expression on T cells at epithelial surfaces along with inducing Foxp3+ Treg cell development.48 The increased percentage of CD103+ Foxp3+ Treg cells detected at days 3–4 days post IT LPS administration may be due in part to increased levels of transforming growth factor-β1 during injury.8
We detected a decrease in Foxp3+ Treg cells in the lung interstitium in injured animals treated with anti-CD103 blockade, which in turn affected the rate of epithelial proliferation. Moreover, when correlating the number of Foxp3+ Treg cells to the number of proliferating CD326+ cells in these conditions we found a strong linear correlation. Moreover, in contrast to CD103+ Treg cells, adoptively transferring CD103−/− Treg cells into Rag-1−/− mice did not augment epithelial proliferation. CD103 blockade did not impact lung injury resolution. Several possibilities could explain these findings. First, it is possible that greater blockade is needed to see an effect on injury resolution as the number of lung Foxp3+ Treg cells still increased after CD103 blockade but not to the levels of isotype antibody control. The decrease Foxp3+ Treg cell number after LPS-induced injury by CD103 blockade may allow for changes in detection of epithelial proliferation but may be enough to adequately suppress the ability of Foxp3+ Treg cells to dampen alveolar inflammation. Furthermore, mechanisms underlying Foxp3+ Treg modulation of epithelial repair (e.g. CD103) may be distinct from pathways used by Foxp3+ Tregs to modulate inflammation or abrogate macrophage/neutrophil responses. Nevertheless, the AT experiments with Rag-1−/− mice transferred CD103−/− Foxp3+ Treg cells, and our co-culture experiments with Foxp3+ Treg and AT2 cells in the presence of anti-CD103 antibodies support the a role for this integrin’s contribution in Foxp3+ Treg cell-mediated epithelial repair.
The in vitro co-culture studies demonstrate that there is a contact-independent effect along with the results demonstrating that blocking CD103 abrogates Foxp3+ Treg cell effects on AT2 epithelial cell proliferation in vitro. These findings strongly indicate an interaction between Foxp3+ Treg cells and AT2 cells in culture, and this interaction in part is driven by CD103 as demonstrated by the antibody-mediated blockade effects. Several potential mechanisms from these experiments can be considered and may not be mutually exclusive. First, CD103’s role could be ascribed to retain Foxp3+ Treg cells to epithelial surfaces. The potential mechanism for decreased epithelial proliferation with antibody-mediated blockade in vivo is a decrease in the number of epithelial-localized Foxp3+ Treg cells, which then exerts a CD103-independent effect on epithelial proliferation detached from whether Foxp3+ Treg cells specifically express CD103 or not. This potential mechanism is supported in part by the AT experiments where Rag-1−/− mice were transferred CD103−/− Foxp3+ Treg cells (Figure 4). This subset of ATs had a lower percentage of epithelial proliferation and a lower number of Foxp3+ cells in the lung compared to the WT Treg AT subset. However, the in vitro co-culture experiment where anti-CD103 antibodies were added to the co-culture of Foxp3+ Treg cells and AT2 cells already in direct contact demonstrated a lower level of AT2 proliferation compared with IgG isotype controls. This strongly supports a role for Foxp3+ Treg cell-expressed CD103 in AT2 proliferation (Figure 6). Another potential mechanism is blocking CD103 may downregulate Foxp3+ Treg cell expression of a pro-proliferative soluble mediator ,which enhances epithelial proliferation (such as a cytokine or growth factor). Foxp3+ Treg cells may demonstrate similar ability to induce epithelial cell proliferation through production of growth factors such as keratinocyte growth factor similar to that shown by intraepithelial γδ T cells,43, 49 and may help explain a contact-independent mechanism for epithelial proliferation. This is supported by the contact-independent experiment where Foxp3+ Treg cells still exert an effect on AT2 proliferation across a transwell membrane. A third possibility is that CD103 antibody-mediated blockade may downregulate or inactivate cell-to-cell CD103–E-cadherin interactions that may be important to enhance downstream pro-proliferative signaling in epithelial cells. This mechanism could be supported by the in vivo antibody-mediated blockade data (Figure 3), and also by the antibody-mediated blockade in vitro experiments (Figure 6). The transwell in vitro experiments however suggest that this may not be the major mechanism.
In whole, the in vivo CD103 antibody-mediated blockade results suggest that retention of Foxp3+ Treg cells during time points of ALI resolution are directly correlated to increased levels of epithelial proliferation (see Figure 3f), i.e., more Foxp3+ Treg cells localized to the epithelium during resolution the greater the percentage of epithelial proliferation. In vivo and in vitro antibody-mediated blockade may affect this interaction through one or more several possible mechanisms listed above. The transwell contact-independent experiments performed suggest that not all of the pro-proliferative effects of Foxp3+ Treg cells on AT2 cells in vitro are mediated through direct Foxp3+ Treg cells–alveolar epithelial communication, and possibly other mechanisms besides CD103 may be important. Determining the extent of the contact-dependent and/or contact-independent factors are future efforts of the authors.
The co-culture data suggests that the interaction of AT2 cells and Foxp3+ Treg cells may also influence surfactant production and differentiation of AT2 cells after injury as detected by differences in SP-C–GFP expression versus T1α expression by imaging. This interaction could extend to other subsets of epithelial cells in the lung, such as Club or AT1 cells. Growth of purified AT2 cells in vitro have been complicated by their ability to proliferate along with the findings that AT2 cells trans-differentiate to a AT1-like phenotype in the absence of specific growth factors.50, 51, 52 The role of AT1 cells in injury repair is unknown and there have been reports suggesting that AT1 undergo proliferation in vitro;53 however, the current thought is that AT1 cells are terminally differentiated and do not undergo proliferation.39 If AT1 cells do proliferate after injury then further work investigating the interactions between AT1 and Foxp3+ Treg cells should be studied.
In a reciprocal manner, there have been reports that AT2 cells induce Foxp3+ Treg cells in an antigen-dependent manner, and the presence of surfactant protein A has been demonstrated to enhance Foxp3+ Treg cell numbers after LPS-induced injury through a transforming growth factor-β-dependent mechanism.22, 54
Lastly, the roles for Foxp3+ Treg cells in mediating the response to injury that indirectly impact AT2 proliferation cannot be excluded and is a future area of investigation. We cannot exclude that a contribution of Foxp3+ Treg cells contribute through modulation of a third cell type such as dendritic cells or macrophages. For instance, Foxp3+ Treg cells decrease tumor necrosis factor-α production by macrophages in a contact-dependent manner.8 Furthermore, others have demonstrated in the PNX model that primary capillary endothelial cells increase MMP14, which in turn exposes epidermal growth factor receptor and elicits growth of epithelial cells after unilateral PNX.15 The interactions of pulmonary capillary endothelial cells and Foxp3+ Treg cells may also be important to the effect of proliferation by Foxp3+ Treg cells in PNX.15 However, the in vitro co-culture experiments lend support to a direct interaction and effect of Foxp3+ Treg cells on the lung epithelium.
In summary, alveolar epithelial repair is a dynamic process and further studies are needed to clarify the role of the cell types involved in the reparative process after ALI. Moreover, recent studies have demonstrated that Foxp3+ Treg cells may have additional functions in non-lymphoid tissues.55 These studies suggest an important role for Foxp3+ Treg cells in alveolar epithelial repair after LPS-induced ALI. Furthermore, Foxp3+ Treg cells also promote CD326+ epithelial cell proliferation in a non-inflammatory compensatory lung growth model. The integrin CD103 has a role in retaining Foxp3+ Treg cells in the lung after injury and blocking this interaction abrogates the Foxp3+ Treg cell proliferative effects on the lung epithelium. This work highlights an undescribed role Foxp3+ Treg cells in alveolar epithelial proliferation. A better understanding of the interactions of Foxp3+ Treg cells and alveolar epithelium during ALI resolution may provide valuable insight into the processes of resolution after injury and potentially uncover mechanisms that enhance endogenous lung repair.
Methods
Mice. C57BL/6 WT, Rag-1−/−, and αE−/− (CD103−/−) mice (6–8 weeks of age) were purchased from The Jackson Laboratory (Bar Harbor, ME) and housed in a pathogen-free facility. SP-CGFP mice were a gift from Dr John K. Heath (University of Birmingham). Foxp3gfp and Foxp3DTR mice were gifts of Dr Alexander Y. Rudensky (Sloan-Kettering Institute). Procedures were approved and conducted under protocols by the Johns Hopkins Animal Care and Use Committee.
LPS administration. Mice were anesthetized and LPS instilled as previously described.8 Additional detail is provided in an online data supplement.
Unilateral left lung PNX. Nine- to 12-week-old C57BL/6, Rag-1−/−, Foxp3gfp, and Foxp3DTR mice underwent left PNX, and lung tissue obtained at 3, 7, or 14 day post procedure for endpoints as previously described.15
Diphtheria toxin administration. Diphtheria toxin (List Biologicals Laboratories, Campbell, CA) was suspended in phosphate-buffered saline. Stock solutions were thawed once and mice were injected i.p. with 50 or 10 μg kg−1 of diphtheria toxin similar as previously described.26
In vivo blockade of CD103. WT animals were given 0.15 mg/dose/mouse of i.p. injections of a blocking CD103 rat polyclonal antibody (M290; BioXcell, West Lebanon, NH) or isotype (rat IgG, Sigma-Aldrich, St Louis, MO) on days 0, 1, 3 and 5 after IT LPS challenge.
Analysis of BAL fluid. BAL was obtained by cannulating the trachea with a 20-gauge catheter, and lungs lavaged and fluid obtained as previously described.8 Additional detail is provided in an online data supplement.
Lung morphology. Lungs from animals were inflated to 25 cm H2O with 1% of low-melting agarose (Invitrogen, Carlsbad, CA) for histological evaluation by hematoxylin and eosin staining.8
Preparation of lung single-cell suspensions for multi-color flow cytometry. After obtaining the BAL fluid for experiments the lungs were infused with 1 ml dispase (BD Bioscience, East Rutherford, NJ) and elastase 3 U ml−1 (Worthington Biochemical, Lakewood, NJ) before 1% (wt/vol) low-melting agarose (Invitrogen) infusion similar to previous descriptions.7, 16 Lungs were minced and filtered as described8, 16 and additional detail is provided in an online data supplement.
Isolation of CD4+ CD25+ T cells and CD4+ CD25− T cells, and AT. Spleens were removed and prepared for single-cell suspensions. CD4+ T cells were isolated from the resulting splenocytes using magnetic bead separation as previously described.8
BAL RAGE levels. BAL fluid was used in enzyme-linked immunosorbent assays to determine levels of soluble RAGE (R&D Systems, Minneapolis, MN) as previously described.23
Isolation of AT2 cells and co-culture experiments. Single-cell suspension was obtained as per the flow cytometry method above and labeled for surface stains before sorting performed with FACSAria and FACSDiva software (Becton Dickinson, BD Bioscience) as previously described.7 Additional detail is provided in an online data supplement.
Immunoblot analysis. Whole lung samples were prepared and used for immunoblot analysis as previously described.56 Additional detail is provided in an online data supplement.
Immunofluorescence. AT2 cell were grown on chamber slides (Lab-Tek, Waltham, MA) and at time points of interest the media was removed and the cells were washed with phosphate-buffered saline. Next, APC-conjugated anti-mouse podoplanin (Biolegend, San Diego, CA) was added to phosphate-buffered saline with 3% albumin and incubated at 37 °C for 30 min. The slides were then washed with phosphate-buffered saline and then slides covered with Fluoromount (Sigma-Aldrich), sealed, and visualized by immunofluorescence with a Leica SP2 Confocal Microscope (Buffalo Grove, IL).
Statistical analysis. Markers of injury were compared using Mann–Whitney rank sum test. Pair-wise comparisons were made by using either Student’s two-tailed unpaired t-test or Mann–Whitney rank sum test. Data are expressed as the mean±s.e.m. where applicable. Statistical analysis was performed using GraphPad Prism 5 software (La Jolla, CA). Statistical difference was accepted at P<0.05.
References
Ware, L.B. & Matthay, M.A. The acute respiratory distress syndrome. N. Engl. J. Med. 342, 1334–1349 (2000).
Ranieri, V.M. et al. Acute respiratory distress syndrome: the Berlin Definition. JAMA 307, 2526–2533 (2012).
Brower, R.G. et al. Ventilation with lower tidal volumes as compared with traditional tidal volumes for acute lung injury and the acute respiratory distress syndrome. The Acute Respiratory Distress Syndrome Network. N. Engl. J. Med. 342, 1301–1308 (2000).
Guerin, C. et al. Prone positioning in severe acute respiratory distress syndrome. N. Engl. J. Med. 368, 2159–2168 (2013).
Mizgerd, J.P. Respiratory infection and the impact of pulmonary immunity on lung health and disease. Am. J. Respir. Crit. Care Med. 186, 824–829 (2012).
Bhattacharya, J. & Matthay, M.A. Regulation and repair of the alveolar-capillary barrier in acute lung injury. Annu. Rev. Physiol. 75, 593–615 (2013).
Garibaldi, B.T. et al. Regulatory T cells reduce acute lung injury fibroproliferation by decreasing fibrocyte recruitment. Am. J. Respir. Cell. Mol. Biol. 48, 35–43 (2013).
D'Alessio, F.R. et al. CD4+CD25+Foxp3+ Tregs resolve experimental lung injury in mice and are present in humans with acute lung injury. J. Clin. Invest. 119, 2898–2913 (2009).
Fontenot, J.D., Gavin, M.A. & Rudensky, A.Y. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat. Immunol. 4, 330–336 (2003).
Abbas, A.K. et al. Regulatory T cells: recommendations to simplify the nomenclature. Nat. Immunol. 14, 307–308 (2013).
Shevach, E.M. CD4+ CD25+ suppressor T cells: more questions than answers. Nat. Rev. Immunol. 2, 389–400 (2002).
Vignali, D.A., Collison, L.W. & Workman, C.J. How regulatory T cells work. Nat. Rev. Immunol. 8, 523–532 (2008).
Miyara, M. & Sakaguchi, S. Natural regulatory T cells: mechanisms of suppression. Trends Mol. Med. 13, 108–116 (2007).
Ehrentraut, H. et al. CD73+ regulatory T cells contribute to adenosine-mediated resolution of acute lung injury. FASEB J. 27, 2207–2219 (2013).
Ding, B.S. et al. Endothelial-derived angiocrine signals induce and sustain regenerative lung alveolarization. Cell 147, 539–553 (2011).
Kim, C.F. et al. Identification of bronchioalveolar stem cells in normal lung and lung cancer. Cell 121, 823–835 (2005).
Fujino, N. et al. A novel method for isolating individual cellular components from the adult human distal lung. Am. J. Respir. Cell. Mol. Biol. 46, 422–430 (2012).
Giangreco, A., Shen, H., Reynolds, S.D. & Stripp, B.R. Molecular phenotype of airway side population cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 286, L624–L630 (2004).
Lee, J.H. et al. SPC H2B-GFP mice reveal heterogeneity of surfactant protein C-expressing lung cells. Am. J. Respir. Cell. Mol. Biol. 48, 288–289 (2012).
Yamamoto, K. et al. Type I alveolar epithelial cells mount innate immune responses during pneumococcal pneumonia. J. Immunol. 189, 2450–2459 (2012).
Dobbs, L.G., Williams, M.C. & Gonzalez, R. Monoclonal antibodies specific to apical surfaces of rat alveolar type I cells bind to surfaces of cultured, but not freshly isolated, type II cells. Biochim. Biophys. Acta 970, 146–156 (1988).
Lo, B., Hansen, S., Evans, K., Heath, J.K. & Wright, J.R. Alveolar epithelial type II cells induce T cell tolerance to specific antigen. J. Immunol. 180, 881–888 (2008).
Su, X., Looney, M.R., Gupta, N. & Matthay, M.A. Receptor for advanced glycation end-products (RAGE) is an indicator of direct lung injury in models of experimental lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 297, L1–L5 (2009).
van Roy, F. & Berx, G. The cell-cell adhesion molecule E-cadherin. Cell Mol. Life Sci. 65, 3756–3788 (2008).
Fontenot, J.D. et al. Regulatory T cell lineage specification by the forkhead transcription factor foxp3. Immunity 22, 329–341 (2005).
Kim, J.M., Rasmussen, J.P. & Rudensky, A.Y. Regulatory T cells prevent catastrophic autoimmunity throughout the lifespan of mice. Nat. Immunol. 8, 191–197 (2007).
Boehm, F. et al. Deletion of Foxp3+ regulatory T cells in genetically targeted mice supports development of intestinal inflammation. BMC Gastroenterol. 12, 97 (2012).
Lehmann, J. et al. Expression of the integrin alpha Ebeta 7 identifies unique subsets of CD25+ as well as CD25− regulatory T cells. Proc. Natl Acad. Sci. USA 99, 13031–13036 (2002).
Cepek, K.L. et al. Adhesion between epithelial cells and T lymphocytes mediated by E-cadherin and the alpha E beta 7 integrin. Nature 372, 190–193 (1994).
Schon, M.P. et al. Mucosal T lymphocyte numbers are selectively reduced in integrin alpha E (CD103)-deficient mice. J. Immunol. 162, 6641–6649 (1999).
Hsia, C.C. Signals and mechanisms of compensatory lung growth. J. Appl. Physiol. 97, 1992–1998 (2004).
Smyth, L.J., Kirby, J.A. & Cunningham, A.C. Role of the mucosal integrin alpha(E)(CD103)beta(7) in tissue-restricted cytotoxicity. Clin. Exp. Immunol. 149, 162–170 (2007).
Wang, L. et al. BLT1-dependent alveolar recruitment of CD4(+)CD25(+) Foxp3(+) regulatory T cells is important for resolution of acute lung injury. Am. J. Respir. Crit. Care Med. 186, 989–998 (2012).
Soroosh, P. et al. Lung-resident tissue macrophages generate Foxp3+ regulatory T cells and promote airway tolerance. J. Exp. Med. 210, 775–788 (2013).
Rojas, M., Woods, C.R., Mora, A.L., Xu, J. & Brigham, K.L. Endotoxin-induced lung injury in mice: structural, functional, and biochemical responses. Am. J. Physiol. Lung Cell. Mol. Physiol. 288, L333–L341 (2005).
Marsh, L.M. et al. Surface expression of CD74 by type II alveolar epithelial cells: a potential mechanism for macrophage migration inhibitory factor-induced epithelial repair. Am. J. Physiol. Lung Cell. Mol. Physiol. 296, L442–L452 (2009).
Cunningham, A.C. et al. Constitutive expression of MHC and adhesion molecules by alveolar epithelial cells (type II pneumocytes) isolated from human lung and comparison with immunocytochemical findings. J. Cell. Sci. 107 (Pt 2), 443–449 (1994).
Rock, J. & Konigshoff, M. Endogenous lung regeneration: potential and limitations. Am. J. Respir. Crit. Care Med. 186, 1213–1219 (2012).
Evans, M.J., Cabral, L.J., Stephens, R.J. & Freeman, G. Transformation of alveolar type 2 cells to type 1 cells following exposure to NO2. Exp. Mol. Pathol. 22, 142–150 (1975).
Fehrenbach, H. Alveolar epithelial type II cell: defender of the alveolus revisited. Respir. Res. 2, 33–46 (2001).
Evans, C.M., Phillips, A.D., Walker-Smith, J.A. & MacDonald, T.T. Activation of lamina propria T cells induces crypt epithelial proliferation and goblet cell depletion in cultured human fetal colon. Gut 33, 230–235 (1992).
Castro, G.A. Immunological regulation of epithelial function. Am. J. Physiol. 243, G321–G329 (1982).
Boismenu, R. & Havran, W.L. Modulation of epithelial cell growth by intraepithelial gamma delta T cells. Science 266, 1253–1255 (1994).
Burzyn, D. et al. A special population of regulatory T cells potentiates muscle repair. Cell 155, 1282–1295 (2013).
Belkaid, Y. & Rouse, B.T. Natural regulatory T cells in infectious disease. Nat. Immunol. 6, 353–360 (2005).
Cerf-Bensussan, N. et al. A monoclonal antibody (HML-1) defining a novel membrane molecule present on human intestinal lymphocytes. Eur. J. Immunol. 17, 1279–1285 (1987).
Kilshaw, P.J. & Murant, S.J. Expression and regulation of beta 7(beta p) integrins on mouse lymphocytes: relevance to the mucosal immune system. Eur. J. Immunol. 21, 2591–2597 (1991).
Weiner, H.L. Induction and mechanism of action of transforming growth factor-beta-secreting Th3 regulatory cells. Immunol. Rev. 182, 207–214 (2001).
Boismenu, R. & Havran, W.L. Gammadelta T cells in host defense and epithelial cell biology. Clin. Immunol. Immunopathol. 86, 121–133 (1998).
Leslie, C.C., McCormick-Shannon, K., Mason, R.J. & Shannon, J.M. Proliferation of rat alveolar epithelial cells in low density primary culture. Am. J. Respir. Cell. Mol. Biol. 9, 64–72 (1993).
Sugahara, K., Mason, R.J. & Shannon, J.M. Effects of soluble factors and extracellular matrix on DNA synthesis and surfactant gene expression in primary cultures of rat alveolar type II cells. Cell Tissue Res. 291, 295–303 (1998).
Chen, Z. et al. Identification of two novel markers for alveolar epithelial type I and II cells. Biochem. Biophys. Res. Commun. 319, 774–780 (2004).
Gonzalez, R.F., Allen, L. & Dobbs, L.G. Rat alveolar type I cells proliferate, express OCT-4, and exhibit phenotypic plasticity in vitro. Am. J. Physiol. Lung Cell. Mol. Physiol. 297, L1045–L1055 (2009).
Mukherjee, S. et al. Surfactant protein A modulates induction of regulatory T cells via TGF-beta. J. Immunol. 188, 4376–4384 (2012).
Burzyn, D., Benoist, C. & Mathis, D. Regulatory T cells in nonlymphoid tissues. Nat. Immunol. 14, 1007–1013 (2013).
Mock, J.R. et al. Haemophilus ducreyi targets Src family protein tyrosine kinases to inhibit phagocytic signaling. Infect. Immun. 73, 7808–7816 (2005).
Acknowledgements
We thank Andre Robinson and James Watkins for expert assistance with tissue processing for histological studies. We thank Dr Mark Soloski, Joe Chrest, and Raffaello Cimbro for their assistance in the Johns Hopkins Bayview Flow Cytometry Core. We thank Dr Enid Neptune for use of the SP-CGFP mice, which were a gift from Dr John Heath and Dr Jo Rae Wright. We thank Dr Alexander Rudensky for use of the Foxp3gfp and Foxp3DTR mice strains. This study was supported by the NIH NRSA F32HL110561 (J.R.M.), NIH R00HL103973 (F.R.D.), NIH R01HL089346 (L.S.K.), and Johns Hopkins Bayview Scholars Program (L.S.K.).
Author contributions
J.R.M., B.T.G., N.R.A., B.D.S., V.S., L.S.K., and F.R.D.: conceived and designed experiments; J.R.M., B.T.G., J.J., N.L., B.D.S., E.C., R.R., and D.C.F.: performed experiments and analysis; B.T.G. and B.D.S.: provided assistance with statistical analysis; J.R.M., N.R.A., B.D.S., W.M., E.M.W., L.S.K., and F.R.D.: wrote the manuscript and provided creative input.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declared no conflict of interest.
Additional information
SUPPLEMENTARY MATERIAL is linked to the online version of the paper
Supplementary information
Rights and permissions
About this article

Cite this article
Mock, J., Garibaldi, B., Aggarwal, N. et al. Foxp3+ regulatory T cells promote lung epithelial proliferation. Mucosal Immunol 7, 1440–1451 (2014). https://doi.org/10.1038/mi.2014.33
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mi.2014.33
This article is cited by
-
Dexmedetomidine alleviates acute lung injury by promoting Tregs differentiation via activation of AMPK/SIRT1 pathway
Inflammopharmacology (2023)
-
The role of T cells in age-related diseases
Nature Reviews Immunology (2022)
-
Type II alveolar cell MHCII improves respiratory viral disease outcomes while exhibiting limited antigen presentation
Nature Communications (2021)
-
Tissue regulatory T cells: regulatory chameleons
Nature Reviews Immunology (2021)
-
Regulatory role of Gpr84 in the switch of alveolar macrophages from CD11blo to CD11bhi status during lung injury process
Mucosal Immunology (2020)
