
Abstract
CD68 is a heavily glycosylated glycoprotein that is highly expressed in macrophages and other mononuclear phagocytes. Traditionally, CD68 is exploited as a valuable cytochemical marker to immunostain monocyte/macrophages in the histochemical analysis of inflamed tissues, tumor tissues, and other immunohistopathological applications. CD68 alone or in combination with other cell markers of tumor-associated macrophages showed a good predictive value as a prognostic marker of survival in cancer patients. Lowression of CD68 was found in the lymphoid cells, non-hematopoietic cells (fibroblasts, endothelial cells, etc), and tumor cells. Cell-specific CD68 expression and differentiated expression levels are determined by the complex interplay between transcription factors, regulatory transcriptional elements, and epigenetic factors. Human CD68 and its mouse ortholog macrosialin belong to the family of LAMP proteins located in the lysosomal membrane and share many structural similarities such as the presence of the LAMP-like domain. Except for a second LAMP-like domain present in LAMPs, CD68/microsialin has a highly glycosylated mucin-like domain involved in ligand binding. CD68 has been shown to bind oxLDL, phosphatidylserine, apoptotic cells and serve as a receptor for malaria sporozoite in liver infection. CD68 is mainly located in the endosomal/lysosomal compartment but can rapidly shuttle to the cell surface. However, the role of CD68 as a scavenger receptor remains to be confirmed. It seems that CD68 is not involved in binding bacterial/viral pathogens, innate, inflammatory or humoral immune responses, although it may potentially be involved in antigen processing/presentation. CD68 could be functionally important in osteoclasts since its deletion leads to reduced bone resorption capacity. The role of CD68 in atherosclerosis is contradictory.
Similar content being viewed by others

Main
CD68 is a heavily glycosylated type I transmembrane glycoprotein, which is mainly associated with endosomal/lysosomal compartment. 1 CD68 shares structural similarities with lysosomal-associated membrane proteins (LAMPs) and belongs to the LAMP family of glycoproteins.2 Along with CD68, that is otherwise called LAMP-4, the LAMP family comprises five lysosome-specific protein members located in the membrane including LAMP-1, LAMP-2, dendritic cell (DC)-LAMP (also known as CD208), and brain and dendritic cell-associated (BAD)-LAMP (also known as LAMP-5). LAMP-1 and LAMP-2 include two LAMP-like domains exposed to the lysosomal lumen and separated by a proline-rich hinge region. The C-terminal portion of LAMPs comprises the transmembrane region followed by a very short cytoplasmic tail. Compared with LAMP-1 and LAMP-2, CD68 contains only a single LAMP-like domain, containing two conserved disulfide bridges, and a mucin-like domain.3 LAMPs and lysosomal integral membrane proteins (LIMPs) are abundantly presented in the lysosomal membrane accounting for about 50% of the membrane protein in lysosomes. The cytosolic tail of LAMPs/LIMPs contains single tyrosine or di-leucine motifs essential for binding adaptor complexes (APS) for further sorting at the trans-Golgi network and targeting to lysosomes.4 LAMP-1 regulates lysosomal exocytosis and fusion with the plasma membrane.5 Lack of LAMP-1/2 leads to the abnormalities in the transport of cytoplasmic peptides for lysosomal degradation6 and abnormal accumulation of cholesterol in lysosomes.7 Human LAMP-2 deficiency causes X-linked inherited pathology (Danon disease) associated with muscle atrophy, cardiomyopathy, and mental retardation due to the intracellular accumulation of glycogen and autophageal material.8 However, so far, the function of LAMPs is yet to be fully investigated.
The function of CD68 is poorly investigated but its preferential location within late endosomes may suggest a role in peptide transport/antigen processing.9 Murine CD68/macrosialin was shown to be mainly located in intracellular pools (85–90%), which exchange at a high rate with CD68 expressed on the surface.10 As estimated, 5000 CD68 molecules can be expressed on the surface of macrophages.11 As was shown in cultured monocyte THP-1 cells, surface CD68 binds oxidized low-density lipoprotein (oxLDL) suggesting a putative role as a receptor for oxLDL.12 This property of CD68 can be disrupted by anti-CD68 monoclonal antibodies. However, so far, the role of CD68 as a receptor for oxLDL has not been confirmed in vivo or in vitro with the purified protein.13
CD68 is well known as a myeloid-specific surface marker, especially abundantly expressed by macrophages.14 However, cytochemical studies showed that non-myeloid cells can also express CD68 but to a lesser extent than myeloid cells.15 In this review, we consider CD68 structure and expression in myeloid and non-myeloid cell types along with its role in normal and pathological conditions.
CD68 Structure and expression
The 2.6-kb-long human CD68 gene is located on chromosome 17p13 and comprises six exons.1 The 1.9-kb-long murine macrosialin/CD68 gene is located on chromosome 11 also containing six exons.16 The mouse gene lacks a classical TATA-box but contains other transcriptional regulatory sites consistent with preferential monocyte/macrophage gene expression. In the human CD68 gene, intron 1 serves as a macrophage-specific transcriptional enhancer. Intron 1 of the mouse gene lacks this capacity thereby suggesting different organization of transcriptional regulatory regions in the human and murine CD68 genes.17
Role of Transcription Regulatory Sites and Transcription Factors in Cell-Specific Expression of CD68
In the bone marrow, expression of CD68 along with other macrophage-specific genes is induced by macrophage/granulocyte-macrophage colony-stimulating factors (M-CSF/GM-CSF) in common myeloid progenitors during diversification between lymphoid and myeloid lineages.18, 19 In mouse, CD68+lin− plasmacytoid monocytes highly expressing CD68 but lacking other myeloid surface (CD14−, CD33−, CD13−, CD11b−, CD11c−) and lysosomal (myeloperoxidase and lysozyme) markers have been observed. These cells represent primary precursors of dendritic cells (DCs) since they may differentiate into typical DCs upon stimulation with IL-3 and CD40 ligand.20
CD68 is highly expressed in cells of the mononuclear phagocyte lineage including macrophages, microglia, osteoclasts, and myeloid dendritic cells (DCs).18 Recently, Iqbal et al21 reported construction of human promoter CD68 green fluorescent protein (GFP) in mice that provides a possibility to observe trafficking of CD68/GFP-positive monocytes and their differentiation to macrophages in vivo. PU.1, an ETS-domain transcription factor that activates gene expression during myeloid and B-lymphoid cell development along with c-Jun drive expression of the murine CD68 gene and C-Jun is required for maximal expression of CD68 in monocytic cells.22 The murine CD68 promoter region contains several regulatory elements between −7.0 and −2.5 kb from the transcriptional start site, which have strong enhancer activity in macrophages and repressor activity in non-myeloid cells. E26 transformation-specific related transcription factor (Elf-1) and Friend leukemia integration 1 transcription factor (Fli-1), also cooperates with PU-1 in priming CD68 expression.23 The proximal -150-bp sequence of the mouse CD68 promoter exhibits high level promoter activity in macrophages. Ets factors (Elf-1 and Fli-1) associate with the -106-bp site while PU.1 binds to the -89-bp site that represents a composite element also recognized by interferon-regulatory factors (IRFs).23 In hematopoietic cells, IRF-4 and IRF-8 are specifically expressed driving development of myeloid and lymphoid lineages. Both IRFs are able to form heterocomplexes with PU.1.24 However, binding of PU.1/IRF-4 or PU/IRF-8 to the -89 site results in the inhibition of CD68 expression in vitro. Chromatin immunoprecipitation data revealed that neither IRF-4 nor IRF-8 associate with the CD68 -89 site in macrophages in vivo but IRF-4 is associated with this site in B cells causing inhibition of CD68 expression.23
In addition to high expression in the mononuclear phagocyte lineage, considerably lower levels of CD68 are expressed in lymphoid cells such as CD19+ B lymphocytes and CD4+ T lymphocytes. CD68 expression was also found in granulocytes such as basophils (taken from patients with myeloproliferative disorders) and intestinal neutrophils derived from inflamed sites of patients with inflammatory bowel disease. CD68-positive neutrophils were almost absent in the normal mucosal tissue.25 The analysis of the distribution and phosphorylation state of polymerase II across the CD68 gene showed that chromatin architecture greatly influences transcription of CD68 in permissive (ie, monocytoid) and non-permissive (ie, lymphoid) cell lines by promoting CD68 expression in monocytoid cells and inhibiting transcription in lymphoid cells.26 Indeed, chromatin architecture is likely to permit inhibitory binding of IRF-4 to the CD68 promoter in lymphoid cells.
Low expression of CD68 was found in several non-hematopoietic cell types including human umbilical cord mesenchymal stem cells,27 fibroblasts, endothelial cells, and various tumor cell lines,15 as well as intimal smooth muscle cells of human arteries.28 Indeed, finding of CD68 in many non-myeloid cell types questions the mononuclear phagocyte lineage specificity of this surface marker even though CD68 expression is much stronger in myeloid cells. The analysis of CD68 transcripts in different cell types showed that fibroblasts preferentially produce the longest mRNA transcript while monocytes synthesize equal amounts of the long and short transcript while DCs and macrophages mainly express the shortest mRNA.15 Indeed, in fibroblasts and myeloid cells, CD68 transcription is initiated from different sites. The analysis of the CD68 promoter revealed that fibroblast CD68 transcription is started at -89 site containing the consensus Ets/PU.1/interferon responsive element (IRE) sequence while in macrophages and DCs transcription is predominantly initiated at the site located approximately 40 bp downstream.15 In fact, myeloid and non-myeloid cells utilize two distinct alternative promoters, which can determine (along with the influence of chromatin architecture in the promoter region) the marked difference in CD68 expression levels between these cell types.
Protein Structure of Human CD68 and Mouse Macrosialin
Human CD68 comprises 354 amino acids (a.a.), with the 21-a.a. long signal peptide, mucin-like domain (a.a. 23–140; contains two 30-a.a. long repeats), proline-rich hinge, LAMP-like domain, transmembrane domain (a.a. 320–344), and C-terminal cytosolic tail.2 The murine macrosialin/CD68 is closely related to human CD68 sharing 72.0% identity and 80.6% similarity in amino acid sequence.1 Like in human CD68, the extracellular domain of murine macrosialin has a bipartite structure divided by the proline hinge. In human CD68 and mouse macrosialin, the membrane-proximal LAMP-like domain is compact and includes four regularly spaced cysteines (36–37 residues apart). The cysteine positions are highly conserved in LAMP proteins and contribute to the formation of disulfide bridges between the first and second and between the third and fourth cysteines.29, 30 However, compared with LAMPs, CD68 and macrosialin contain only one LAMP-like domain.
Another LAMP member, DC-LAMP, is also abundantly expressed in DCs and also has a single LAMP-like domain.31 Compared with human CD68, human DC-LAMP is longer as it comprises 416 a.a. In addition, human DC-LAMP contains seven N-glycosylation sites and 34 O-glycosylation sites. Human CD68 has a total of 29 and nine O- and N-glycosylation sites respectively. While CD68 is rather an endosomal protein, DC-LAMP is located in lysosomes.31 In fact, DC-LAMP is a nonspecific marker of DC maturation because it appears before the translocation of the major histocompatibility (MHC) class II molecules to the cell surface, after which it accumulates into perinuclear lysosomes. This may suggest that DC-LAMP might change the lysosome function after the transfer of peptide-MHC class II molecules to the surface of DCs, a sign of the DC maturation. DC-LAMP is involved in the control of MHC class II antigen presentation in DCs through the regulation of MHC II-peptide complex formation and expression on the plasma membrane.32 Probably, the involvement in the control of the presentation/degradation of some antigens can be related to CD68 due to the close structural similarity with DC-LAMP. However, this needs to be verified.
The principal scheme of the domain organization of murine macrosialin and human proteins LAMP-1, LAMP-2, DC-LAMP, and CD68 is provided in Figure 1. It is predicted that all LAMP family proteins contain LAMP domains of similar structure (Figure 2).31, 33 The DC-LAMP domain consists of two β-sheets.33 The domain's N- and C-termini are located on the 'front' β-sheet, which consists of six β-strands. The β-strands S1 and S3 are short and arranged in tandem, antiparallel to strand S2. Two more antiparallel β-strands, S10 and S9, are followed by the short C-terminal β-strand S11, which is arranged in parallel to S9.33

A scheme shows the general features of the domain organization of murine macrosialin and human proteins LAMP-1, LAMP-2, DC-LAMP, and CD68 and depicts the principal differences between the family members. N-glycans and O-glycans are indicated by red and green Ys, respectively. The structure of the LAMP domain is well known;31, 33 the domain folds into a compact β-sheet sandwich. It is essential to note that the LAMP domain does not reach far into the vesicle lumen but is a compactly folded unit directly adjacent to the membrane. As shown in the scheme, each molecule contains the short C-terminal cytoplasmic tail and the transmembrane domain. The main portion of the molecule is located in the lumen of lysosomes/endosomes. The luminal portion of DC-LAMP, CD68, and macrosialin contain the LAMP-like domain and the mucin-like domain linked by a proline-rich hinge. LAMP-1 and LAMP-2 have two LAMP-like domains separated by the proline-rich hinge. The length of the amino acid (a.a) sequence of each protein is shown in brackets. Please note that the LAMP-like and mucin-like domains of DC-LAMP are located very close to the membrane, not being projected too far to the lumen space. The linker between the two LAMP domains in LAMP-1 and LAMP-2 was predicted to adopt rather an elongated conformation and it is not folded as drawn in the scheme. The actual structural organization of the DC-LAMP domain can be seen in Figure 2. Please note also that mucins are known to be very proline rich, so that it is questionable if a mucin-like domain and a proline-rich hinge domain of CD68, DC-LAMP, and macrosialin should be distinguished.

The β-strand arrangement of the DC-LAMP domain. The ‘front’ and ‘back’ β-sheets are drawn in red and blue, respectively. The cysteines that form the first disulfide bond (S–S) are labeled. The β-strands and loops are identified as S1 to S11 and L1 to L3, respectively. The topology of the β-sheets is drawn schematically in the lower part with sheets opened out. (This figure is reproduced from Wilke et al,33 with permission from BioMed Central).
The C-terminal 10-residue tail (RRRFSAYQAL in human CD68 and RRRQSTYQPL in murine macrosialin), is highly conserved in LAMPs indicating lysosomal-specific targeting. While some lysosomal membrane proteins are delivered to lysosomes indirectly (through endocytosis), others such as CD68 reach endosomes/lysosomes primarily from the trans-Golgi network in clathrin-coated vesicles.34 The mucin-like domain is enriched with serine and threonine residues (up to 43% of a total domain a.a. content), a characteristic of mucin-like molecules with high rate of O-glycosylation.35
CD68 Glycosylation
CD68 is highly glycosylated. Both human and murine molecules contain nine putative sites for N-glycosylation four of which are located in the LAMP domain. The number of O-glycosylation sites is much higher (up to 29 in human CD68 and 26 in mouse macrosialin) (Figure 1). All O-glycosylation sites are resided in the mucin-like domain.1 In murine macrosialin, the molecular mass of the polypeptide backbone is only 35 kDa while the entire mass of mature glycoprotein varies from 90 to 120 kDa. The largest contribution to the mass of the mature macrosialin comes from both O- and N-linked sugars (40% and 20–25%, respectively), with only about one-third coming from the polypeptide backbone.36
The differential glycosylation of CD68 is extensively regulated during macrophage activation that results in acquiring numerous O-linked terminal sialic acid residues and N-linked polylactosaminoglycans.37 In rat resident peritoneal macrophages, basal expression of CD68 is low but can be greatly upregulated in response to inflammatory stimuli. Accordingly, CD68 glycosylation is extensively remodeled during proinflammatory activation giving rise to both O-linked and N-linked glycosylation.36 In mouse CD68, phagocytosis increases the complexity and length of O-linked chains that results in increase of ligand-binding capacity (ie, binding of plant agglutinins) by peritoneal macrophages.38 For example, phagocytosis of zymosan, a fungal surface glucan, increases the specific incorporation of D-[2-3H]mannose, D-[6-3H]galactose, N-acetyl-D-[1-3H]glucosamine and L-[5,6-3H]fucose into the polysaccharide portion of CD68, thereby inducing the differential glycosylation of this receptor.38
The glycosylation rate and qualitative composition significantly influences structural and functional properties of glycoproteins. For example, increased glycosylation of the IgG constant fragment (Fc) greatly modulates binding to cellular Fc receptors and complement activation by IgG switching IgG activity from proinflammatory to anti-inflammatory.39, 40 Glycosylation contributes to correct antigen recognition by a T-cell receptor in an immunological synapse.41 For LAMP-2, increased N-linked polylactosamine glycosylation of a newly synthesized protein delays its passage through the Golgi apparatus. The residence time in the Golgi apparatus inversely correlates with the glycosylation rate of LAMP-2.42 N-linked glycosylation preserves LAMP-1 and LAMP-2 from degradation since deglycosylation leads to rapid removal of both proteins.43 However, it remains unclear in which cellular compartment this occurs and which proteases are involved. Similarly, N-glycosylation could have a protective role to prevent the degradation of de novo synthesized CD68 during its lysosomal translocation from the Golgi network. In cytotoxic cells, glycosylated LAMP-1 was shown to protect cytotoxic lymphocytes from self-destruction during the release of cytotoxic granules (CGs) because CG-associated LAMPs appear on the cell surface after CG exocytosis.44 Accordingly, CD68 upregulation seen in activated neutrophils could also contribute to protection against self-damage during CG release. Finally, elevated CD68 glycosylation should strengthen its ligand-binding properties.
CD68 Function: does CD68 contribute to atherosclerosis and foam cell formation?
Macrophages, due to their primary involvement in the clearance of pathogens, cytotoxic molecules, and dead cells, have developed a repertoire of various scavenger receptors (SRs) with the capability to sense a broad spectrum of ligands.45 CD68 was also considered as a member of the family of SRs as a scavenger receptor type D (SCARD)46 because (i) CD68 can be significantly upregulated in macrophages responding to inflammatory stimuli;23, 36 (ii) CD68 is able to bind modified LDL, phosphatidylserine, and apoptotic cells;12, 47, 48 (iii) CD68 can rapidly shuttle between the plasma membrane and endosomes.10 Indeed, the intrinsic ability of CD68 to bind and internalize oxLDL provided an option to consider its putative role in atherogenesis and intracellular lipid accumulation.
The possible involvement of CD68 in atherosclerosis was shown in in vitro and in vivo experiments. In cultured mouse resident peritoneal macrophages, exposure to oxLDL and minimally oxidized LDL stimulated expression of macrosialin.49 THP-1 monocytic cells exposed to oxLDL had upregulated expression of scavenger receptors CD36, CLA-1, and CD68 (but not LOX-1 and SR-A1) and increased oxLDL uptake.50 In vivo studies that assessed a role of CD68 in atherosclerosis were performed in APA hamsters that develop atheromatous plaques in streptozotocin-induced diabetes. In such a model of diabetic atherosclerosis, rabbits develop early lesions (fatty streaks) by 6 weeks after streptozotocin administration along with hyperlipidemia.51 In diabetic hamsters, Yamanouchi et al52 observed expression of SR-AI, CD68, and receptor for advanced glycation endproducts (RAGE) mRNA in foam cells accumulated in early atherosclerotic lesions. Indeed, these observations suggested a likely role of CD68 in foam cell formation.
De Beer et al53 observed marked upregulation of hepatic macrosialin expression in atheroprone C57BL/6 and atheroresistant C3H/HeJ mice fed an atherogenic high-fat diet. However, overproduction of transgenic macrosialin in COS-7 and CHO cells led to significant surface expression but no binding of oxLDL to macrosialin. Inhibition of macrosialin did not affect the binding of oxLDL to macrophages.53 Song et al54 reported that CD68-deficient mononuclear phagocytes showed a potent lipid uptake thereby indicating no significant role of murine CD68 in lipid internalization. Indeed, more recent studies disputed the involvement of CD68 in atherogenic foam cell formation.
It is possible that CD68 has a significant role in diabetes-associated cardiovascular pathology but this should be evaluated in diabetic patients. It would also be interesting to characterize the consequences of CD68 deletion in Apolipoprotein E (ApoE)- or LDL receptor-deficient mice, ie, established mouse model of atherosclerosis.
CD68: A role in inflammation and immunity
CD68 is routinely used as a histochemical/cytochemical marker of inflammation associated with the involvement of monocytes/macrophages.55 CD68 was also used to identify cells of macrophage lineage such as tissue histiocytes, multinucleated giant cells, Kupffer cells, and osteoclasts.56 CD68 may be used in combination with transcription factor markers, pSTAT1, RBP-J, and CMAF to discriminate M1 and M2 macrophage polarization in vivo.57 Ultrastructural studies of human atherosclerotic lesions demonstrated that CD68 is mostly located in lysosomes (Figure 3).58, 59 In contrast to macrophages,59 CD68 is not expressed by dendritic cells (DCs)14, 60 and this helps to immunohistochemically distinguish these two cell types, both of which have antigen-presenting capabilities. Several studies have shown significant upregulation of CD68 expression in macrophages in response to inflammatory stimuli such as exposure to oxLDL12, 49, 50 and chronic stimulation with bacterial lipopolysaccharide (LPS) or inflammatory cytokine interferon-γ (IFN-γ). In microglial cells (brain tissue-resident macrophages), CD68 expression was stimulated by LPS and IFN-γ in a Toll-like receptor 4 (TLR4)-dependent manner.61 CD68 was also upregulated in inflammatory neutrophils from patients with inflammatory bowel disease.25 However, no CD68-specific functional effects were investigated upon CD68 upregulation in inflammatory myeloid cells.

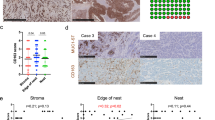
Electron microscopic immunocytochemical demonstration of the presence and distribution of CD68 antigen in primary and secondary lysosomes of macrophage cells located in atherosclerotic lesions of the human aorta (a–d). Immunogold technique. Scale bars, 200 nm (a–d). (Adapted from Bobryshev et al59 with permission from John Wiley and Sons).
Song et al54 comprehensively assessed immune and inflammatory properties of CD68-deficient murine myeloid cells. Like wild-type macrophages, CD68-lacking mouse macrophages exhibited normal phagocytic activity towards killed bacteria Staphylococcus aureus and failed to show defective innate response to infection by Listeria monocytogenes, Legionella pneumophila, and vesicular stomatitis virus. CD68-deficient macrophages showed inflammatory responses equivalent to those of wild-type macrophages. Finally, CD68-deficient DCs demonstrated normal capacity to present antigens and induce humoral immune responses.54 Altogether, the findings of Song et al54 suggest no apparent role for CD68 in inflammatory responses of innate and humoral immunity.
However, recently, Cha et al62 reported finding a peptide P39 by a phage-display library screen. This peptide is able to bind to CD68, a potential receptor for malaria sporozoite, and inhibit entry of the sporozoite to Kupffer cells (liver tissue resident macrophages). Thus, this suggests a role of CD68 as a major gateway for hepatic malaria infection.
Interestingly, Song et al54 observed that CD68-deficient mononuclear phagocytes showed an increased antigen presentation to CD4+ T cells suggesting that CD68 might negatively regulate either antigen uptake and loading or MHC class II trafficking. The role of CD68 in antigen processing is unknown. However, there are some data suggesting the involvement of LAMPs that are structurally close to CD68 in related activities. It has been suggested that LAMP-1/2 regulates phagosome–lysosome fusion and the formation of phagolysosomes.63 Immuno-electron microscopy has shown that LAMP-1 and LAMP-2 are concentrated in adapter protein 3 (AP-3) positive membrane domains associated with budding profiles that emerge from a tubular endosomal compartment.64 During DC maturation, LAMP-2 controls transfer of MHC class II molecules from cytoplasmic stores (ie, from multivesicular bodies where MHC class II molecules are loaded by an antigen) to CD68-containing tubules from which antigen-loaded MHC molecules are delivered to the surface to present an antigen.9, 65 Interestingly, Lin et al66 showed constitutive expression of CD68 along with other key components of the antigen processing/presenting system in small intestinal epithelial cells involved in constitutive processing and presentation of intestinal antigens. All these components were located in vesicular-like structures. This observation suggests a putative involvement of CD68 in antigen processing/presentation.
In summary, the role of CD68 in inflammation and immunity is still mysterious. A study performed by Cha et al62 showed that CD68 can serve as a receptor for binding malaria sporozoite, which promotes subsequent infection of resident hepatic macrophages. Data obtained by Song et al54 indicate a lack of any involvement of macrosialin/CD68 in inflammatory responses to bacterial/viral pathogens and innate/humoral immunity. However, Song et al54 did propose a potential role of CD68 in antigen processing and presentation. Indeed, since CD68 is able to bind and internalize oxLDL, it would be interesting to investigate whether CD68 is involved in the processing and presentation of oxLDL-derived antigenic peptides. If so, this may suggest involvement of CD68 in the production of anti-oxLDL antibodies in atherosclerosis.
CD68 In osteoclasts
Osteoclasts represent a bone-specific population of tissue-resident macrophages involved in bone maintenance, repair, and remodeling. They remove and dissolve protein/mineral aggregates in a process called bone resorption.67 Imbalance in skeletal turnover when bone resorption exceeds bone formation leads to osteoporosis, a disease associated with decreased density and increased fragility of bone.
M-CSF and receptor activator of NF-κB ligand (RANKL; an osteoclast differentiation factor) were shown to induce CD68 expression in osteoclasts. Mice lacking CD68 have skeletal abnormalities associated with increased trabecular bone. CD68-deficient osteoclasts had reduced bone-resorbing capacity.68 Further studies are needed to determine the nature of functional defects observed in osteoclasts deficient for CD68.
CD68 In cancer
CD68 is broadly used as cancer-associated diagnostic and prognostic marker. It is used to identify neoplasms with macrophage lineage and also may be expressed by tumor cells from other lineages. Finding of CD68 expression by tumor cells is not surprising since metastatic tumor cells widely express immune markers to escape macrophage-mediated phagocytosis and cell-damaging effects from cytotoxic CD8+ T cells during invasion of a normal, non-tumor tissue environment. Macrophage molecular traits seem to be the most frequent immune antigens used by emigrated tumor cells including expression of various antigens such as CD14, CD47, CD68, MAC387, CD163, and DAP12.69, 70 Indeed, overexpression of macrophage antigens in tumor tissue may indicate a prometastatic state and may be associated with poor prognosis.71, 72 The molecular mechanisms leading to the expression of immune antigens by tumor cells remain still incompletely understood. The intercellular genetic exchange through cell fusion or release of exosomes may have a role in induction of immune markers in cancer cells.
Of course, the host body fights against the tumor and sends various immune cells to the tumor tissue including DCs and macrophages to induce and conduct the antitumor immune response. Macrophages are the most common type of tumor-infiltrating immune cells.73 However, the tumor microenvironment is anti-inflammatory and immunosuppressive. This microenvironment switches the macrophage phenotype from a proinflammatory state (M1) to an anti-inflammatory state (M2).74 Such CD68-positive macrophages are called tumor-associated macrophages (TAMs). TAMs acquire immunosuppressive properties and inhibit cytotoxic activity of CD8+ T cells in tumors.75 A population of CD68-positive immature myeloid cells (called myeloid-derived suppressor cells (MDSCs) that inhibit proliferation and cytokine release by T cells was found to be present in glioma76 and squamous cell carcinoma tumors.77 These cells can be produced by the tumor microenvironment from circulating monocytes.78
TAMs, MDCDs, and other tumor-associated immune cells compose the leukocytic infiltrate, of which TAMs are the major contributors. Indeed, higher expression of CD68 and other macrophage markers in the tumor stroma usually correlates with higher tumor grade, higher lymph node metastasis, and other malignant characteristics that characterize tumor progression and aggressiveness.79, 80, 81 Except for evaluating tumorigenesis, TAM-associated CD68 expression levels can serve as a useful predictive marker for prognosis of survival of cancer patients.82, 83, 84 In the phase III LYM-3001 study, Coiffier et al85 showed a significant predictive value of a combination of low CD68 tumor expression and a genetic marker (proteasome subunit-β type-1 (PSMB1) PA11A G allele) for a longer progression-free survival of follicular lymphoma patients treated with bortezomib (proteasome inhibitor) and rituximab (anti-CD20 monoclonal antibody) vs treatment with rituximab alone. Indeed, these results suggest a potential role for assessment of CD68 expression in tumors to evaluate the efficiency and outcome prognosis of anticancer therapy.
However, there is a significant inconsistency in the assessment of CD68 immunostaining predictive power. For example, Steidl et al86 suggested CD68 as a surrogate marker for macrophages. The authors86 showed in the independent cohort of patients with classical Hodgkin lymphoma (cHL; a B-cell lymphoproliferative disorder) that the presence of less than 5% of CD68+ TAMs in the nodes correlated with longer progression-free survival after primary therapy and lower recurrence rates after autologous transplantation. However, these results were not clearly confirmed.87, 88 Harris et al89 suggested that CD163 staining is superior vs CD68 in the identification of TAMs in neoplastic nodes owing to lower background staining and less nonspecific staining of background inflammatory cells and Hodgkin cells. In addition, Klein et al90 showed better predictive value using CD163 as a marker of TAMs for survival of cHL patients. Differences in the composition of the case series, technical variability, differences in clones used to produce the antibody, and different cut-offs for expression of TAM-associated markers could, in part, explain the discrepancy in results. Indeed, technical standardization as well as prospective validation in an independent series of cHL patients, with similar treatment protocols and comparable clinical stages, would be required before CD68 immunohistochemical staining is undertaken.
Conclusion
The function of CD68 in inflammation and carcinogenesis is insufficiently studied despite its routine employment as an immunochemical marker of macrophages and other mononuclear phagocytes. CD68 expression remains a valuable tool for the assessment of tumor grade, malignancy, progression, and metastatic potential.
Known natural CD68 ligands are limited to oxLDL, phosphatidylserine, apoptotic cells (due to the presence of phosphatidylserine in the plasma membrane), and notably the malaria sporozoite. The latter is important because it provides evidence for the involvement of CD68 in the life/infectious cycle of malaria plasmodium, a protozoan parasite. It seems that CD68 is not able to recognize bacterial and viral products. The heavily glycosylated extracellular domain of CD68 or its mouse ortholog macrosialin has a crucial role in ligand binding. However, a functional role of two a.a. tandemly repeated sequences located in the mucin-like domain is unknown. It would be interesting to know a role of this tandem in binding ligands, for example, oxLDL. It would also be helpful to extend the number of natural ligands capable of binding to CD68. This could provide new insights into CD68 function.
The contribution of CD68 to immunity remains to be investigated. It seems that CD68 does not have a central role in regulating innate and adaptive immune responses. However, some data indicates that CD68 could potentially be involved in the processing/recognition of antigenic peptides/moieties derived from oxLDL suggesting a possible role in atherogenesis that so far has proven elusive.
References
Holness CL, Simmons DL . Molecular cloning of CD68, a human macrophage marker related to lysosomal glycoproteins. Blood 1993;81:1607–1613.
Holness CL, da Silva RP, Fawcett J et al, Macrosialin, a mouse macrophage-restricted glycoprotein, is a member of the lamp/lgp family. J Biol Chem 1993;268:9661–9666.
Fukuda M . Lysosomal membrane glycoproteins. Structure, biosynthesis, and intracellular trafficking. J Biol Chem 1991;266:21327–21330.
Winchester BG . Lysosomal membrane proteins. Eur J Paediatr Neurol 2001;5 (Suppl A):11–19.
Yogalingam G, Bonten EJ, van de Vlekkert D et al, Neuraminidase 1 is a negative regulator of lysosomal exocytosis. Dev Cell 2008;15:74–86.
Demirel Ö, Jan I, Wolters D et al, The lysosomal polypeptide transporter TAPL is stabilized by interaction with LAMP-1 and LAMP-2. J Cell Sci 2012;125 (Pt 18):4230–4240.
Eskelinen EL, Schmidt CK, Neu S et al, Disturbed cholesterol traffic but normal proteolytic function in LAMP-1/LAMP-2 double-deficient fibroblasts. Mol Biol Cell 2004;15:3132–3145.
Nishino I, Fu J, Tanji K et al, Primary LAMP-2 deficiency causes X-linked vacuolar cardiomyopathy and myopathy (Danon disease). Nature 2000;406:906–910.
Barois N, de Saint-Vis B, Lebecque S et al, MHC class II compartments in human dendritic cells undergo profound structural changes upon activation. Traffic 2002;3:894–905.
Kurushima H, Ramprasad M, Kondratenko N et al, Surface expression and rapid internalization of macrosialin (mouse CD68) on elicited mouse peritoneal macrophages. J Leukoc Biol 2000;67:104–108.
van der Kooij MA, von der Mark EM, Kruijt JK et al, Human monocyte-derived macrophages express an approximately 120-kD Ox-LDL binding protein with strong identity to CD68. Arterioscler Thromb Vasc Biol 1997;17:3107–3116.
Ramprasad MP, Terpstra V, Kondratenko N et al, Cell surface expression of mouse macrosialin and human CD68 and their role as macrophage receptors for oxidized low density lipoprotein. Proc Natl Acad Sci USA 1996;93:14833–14838.
Steinberg D, Witztum JL . Oxidized low-density lipoprotein and atherosclerosis. Arterioscler Thromb Vasc Biol 2010;30:2311–2316.
Betjes MG, Haks MC, Tuk CW et al, Monoclonal antibody EBM11 (anti-CD68) discriminates between dendritic cells and macrophages after short-term culture. Immunobiology 1991;183:79–87.
Gottfried E, Kunz-Schughart LA, Weber A et al, Expression of CD68 in non-myeloid cell types. Scand J Immunol 2008;67:453–463.
Jiang Z, Shih DM, Xia YR et al, Structure, organization, and chromosomal mapping of the gene encoding macrosialin, a macrophage-restricted protein. Genomics 1998;50:199–205.
Greaves DR, Quinn CM, Seldin MF et al, Functional comparison of the murine macrosialin and human CD68 promoters in macrophage and nonmacrophage cell lines. Genomics 1998;54:165–168.
Greaves DR, Gordon S . Macrophage-specific gene expression: current paradigms and future challenges. Int J Hematol 2002;76:6–15.
Kondo M . Lymphoid and myeloid lineage commitment in multipotent hematopoietic progenitors. Immunol Rev 2010;238:37–46.
Strobl H, Scheinecker C, Riedl E et al, Identification of CD68+lin- peripheral blood cells with dendritic precursor characteristics. J Immunol 1998;161:740–748.
Iqbal AJ, McNeill E, Kapellos TS et al, Human CD68 promoter GFP transgenic mice allow analysis of monocyte to macrophage differentiation in vivo. Blood 2014;124:e33–e44.
Li AC, Guidez FR, Collier JG et al, The macrosialin promoter directs high levels of transcriptional activity in macrophages dependent on combinatorial interactions between PU.1 and c-Jun. J Biol Chem 1998;273:5389–5399.
O'Reilly D, Quinn CM, El-Shanawany T et al, Multiple Ets factors and interferon regulatory factor-4 modulate CD68 expression in a cell type-specific manner. J Biol Chem 2003;278:21909–21919.
Günthner R, Anders HJ . Interferon-regulatory factors determine macrophage phenotype polarization. Mediators Inflamm 2013;2013:731023.
Amanzada A, Malik IA, Blaschke M et al, Identification of CD68(+) neutrophil granulocytes in in vitro model of acute inflammation and inflammatory bowel disease. Int J Clin Exp Pathol 2013;6:561–570.
O'Reilly D, Greaves DR . Cell-type-specific expression of the human CD68 gene is associated with changes in Pol II phosphorylation and short-range intrachromosomal gene looping. Genomics 2007;90:407–415.
La Rocca G, Anzalone R, Farina F . The expression of CD68 in human umbilical cord mesenchymal stem cells: new evidences of presence in non-myeloid cell types. Scand J Immunol 2009;70:161–162.
Andreeva ER, Pugach IM, Orekhov AN . Subendothelial smooth muscle cells of human aorta express macrophage antigen in situ and in vitro. Atherosclerosis 1997;135:19–27.
Carlsson SR, Fukuda M . Structure of human lysosomal membrane glycoprotein 1. Assignment of disulfide bonds and visualization of its domain arrangement. J Biol Chem 1989;264:20526–20531.
Arterburn LM, Earles BJ, August JT . The disulfide structure of mouse lysosome-associated membrane protein 1. J Biol Chem 1990;265:7419–7423.
de Saint-Vis B, Vincent J, Vandenabeele S et al, A novel lysosome-associated membrane glycoprotein, DC-LAMP, induced upon DC maturation, is transiently expressed in MHC class II compartment. Immunity 1998;9:325–336.
Villadangos JA, Schnorrer P, Wilson NS . Control of MHC class II antigen presentation in dendritic cells: a balance between creative and destructive forces. Immunol Rev 2005;207:191–205.
Wilke S, Krausze J, Büssow K . Crystal structure of the conserved domain of the DC lysosomal associated membrane protein: implications for the lysosomal glycocalyx. BMC Biol 2012;10:62.
Hunziker W, Geuze HJ . Intracellular trafficking of lysosomal membrane proteins. Bioessays 1996;18:379–389.
Fukuda M . Leukosialin, a major O-glycan-containing sialoglycoprotein defining leukocyte differentiation and malignancy. Glycobiology 1991;1:347–356.
Rabinowitz SS, Gordon S . Macrosialin, a macrophage-restricted membrane sialoprotein differentially glycosylated in response to inflammatory stimuli. J Exp Med 1991;174:827–836.
Rabinowitz S, Gordon S . Differential expression of membrane sialoglycoproteins in exudate and resident mouse peritoneal macrophages. J Cell Sci 1989;93 (Pt 4):623–630.
da Silva RP, Gordon S . Phagocytosis stimulates alternative glycosylation of macrosialin (mouse CD68), a macrophage-specific endosomal protein. Biochem J 1999;338 (Pt 3):687–694.
Böhm S, Schwab I, Lux A et al, The role of sialic acid as a modulator of the anti-inflammatory activity of IgG. Semin Immunopathol 2012;34:443–453.
Quast I, Lünemann JD . Fc glycan-modulated immunoglobulin G effector functions. J Clin Immunol 2014;34 (Suppl 1):S51–S55.
Rudd PM, Wormald MR, Stanfield RL et al, Roles for glycosylation of cell surface receptors involved in cellular immune recognition. J Mol Biol 1999;293:351–366.
Nabi IR, Dennis JW . The extent of polylactosamine glycosylation of MDCK LAMP-2 is determined by its Golgi residence time. Glycobiology 1998;8:947–953.
Kundra R, Kornfeld S . Asparagine-linked oligosaccharides protect Lamp-1 and Lamp-2 from intracellular proteolysis. J Biol Chem 1999;274:31039–31046.
Cohnen A, Chiang SC, Stojanovic A et al, Surface CD107a/LAMP-1 protects natural killer cells from degranulation-associated damage. Blood 2013;122:1411–1418.
Penberthy KK, Ravichandran KS . Apoptotic cell recognition receptors and scavenger receptors. Immunol Rev 2016;269:44–59.
Moore KJ, Freeman MW . Scavenger receptors in atherosclerosis: beyond lipid uptake. Arterioscler Thromb Vasc Biol 2006;26:1702–1711.
Ottnad E, Parthasarathy S, Sambrano GR et al, A macrophage receptor for oxidized low density lipoprotein distinct from the receptor for acetyl low density lipoprotein: partial purification and role in recognition of oxidatively damaged cells. Proc Natl Acad Sci USA 1995;92:1391–1395.
Sambrano GR, Steinberg D . Recognition of oxidatively damaged and apoptotic cells by an oxidized low density lipoprotein receptor on mouse peritoneal macrophages: role of membrane phosphatidylserine. Proc Natl Acad Sci USA 1995;92:1396–1400.
Yoshida H, Quehenberger O, Kondratenko N et al, Minimally oxidized low-density lipoprotein increases expression of scavenger receptor A, CD36, and macrosialin in resident mouse peritoneal macrophages. Arterioscler Thromb Vasc Biol 1998;18:794–802.
Tsukamoto K, Kinoshita M, Kojima K et al, Synergically increased expression of CD36, CLA-1 and CD68, but not of SR-A and LOX-1, with the progression to foam cells from macrophages. J Atheroscler Thromb 2002;9:57–64.
Yamanouchi J, Takatori A, Itagaki S et al, APA hamster model for diabetic atherosclerosis. 2. Analysis of lipids and lipoproteins. Exp Anim 2000;49:267–274.
Yamanouchi J, Takatori A, Nishida E et al, Expression of lipoprotein receptors in the aortic walls of diabetic APA hamsters. Exp Anim 2002;51:33–41.
de Beer MC, Zhao Z, Webb NR et al, Lack of a direct role for macrosialin in oxidized LDL metabolism. J Lipid Res 2003;44:674–685.
Song L, Lee C, Schindler C . Deletion of the murine scavenger receptor CD68. J Lipid Res 2011;52:1542–1550.
Ferenbach D, Hughes J . Macrophages and dendritic cells: what is the difference? Kidney Int 2008;74:5–7.
Brooks E, Simmons-Arnold L, Naud S et al, Multinucleated giant cells’ incidence, immune markers, and significance: a study of 172 cases of papillary thyroid carcinoma. Head Neck Pathol 2009;3:95–99.
Barros MHM, Hauck F, Dreyer JH et al, Macrophage polarization: an immunohistochemical approach for identifying M1 and M2 macrophages. PLoS One 2013;8:1–11.
Bobryshev YV . Monocyte recruitment and foam cell formation in atherosclerosis. Micron 2006;37:208–222.
Bobryshev YV, Shchelkunova TA, Morozov IA et al, Changes of lysosomes in the earliest stages of the development of atherosclerosis. J Cell Mol Med 2013;17:626–635.
Bobryshev YV . Dendritic cells and their role in atherogenesis. Lab Invest 2010;90:970–984.
Papageorgiou IE, Lewen A, Galow LV et al, TLR4-activated microglia require IFN-γ to induce severe neuronal dysfunction and death in situ. Proc Natl Acad Sci USA 2016;113:212–217.
Cha SJ, Park K, Srinivasan P et al, CD68 acts as a major gateway for malaria sporozoite liver infection. J Exp Med 2015;212:1391–1403.
Huynh KK, Eskelinen EL, Scott CC et al, LAMP proteins are required for fusion of lysosomes with phagosomes. EMBO J 2007;26:313–324.
Peden AA, Oorschot V, Hesser BA et al, Localization of the AP-3 adaptor complex defines a novel endosomal exit site for lysosomal membrane proteins. J Cell Biol 2004;164:1065–1076.
Chow A, Toomre D, Garrett W et al, Dendritic cell maturation triggers retrograde MHC class II transport from lysosomes to the plasma membrane. Nature 2002;418:988–994.
Lin XP, Almqvist N, Telemo E . Human small intestinal epithelial cells constitutively express the key elements for antigen processing and the production of exosomes. Blood Cells Mol Dis 2005;35:122–128.
Teitelbaum SL . Bone resorption by osteoclasts. Science 2000;289:1504–1508.
Ashley JW, Shi Z, Zhao H et al, Genetic ablation of CD68 results in mice with increased bone and dysfunctional osteoclasts. PLoS One 2011;6:e25838.
Shabo I, Svanvik J . Expression of macrophage antigens by tumor cells. Adv Exp Med Biol 2011;714:141–150.
Steinert G, Schölch S, Niemietz T et al, Immune escape and survival mechanisms in circulating tumor cells of colorectal cancer. Cancer Res 2014;74:1694–1704.
Jakovic LR, Mihaljevic BS, Perunicic Jovanovic MD et al, The prognostic relevance of tumor associated macrophages in advanced stage classical Hodgkin lymphoma. Leuk Lymphoma 2011;52:1913–1919.
Shabo I, Olsson H, Stål O et al, Breast cancer expression of DAP12 is associated with skeletal and liver metastases and poor survival. Clin Breast Cancer 2013;13:371–377.
De Palma M, Lewis CE . Macrophage regulation of tumor responses to anticancer therapies. Cancer Cell 2013;23:277–286.
Mantovani A, Schioppa T, Porta C et al, Role of tumor-associated macrophages in tumor progression and invasion. Cancer Metastasis Rev 2006;25:315–322.
Kitamura T, BZ1 Qian, Pollard JW . Immune cell promotion of metastasis. Nat Rev Immunol 2015;15:73–86.
Prosniak M, Harshyne LA, Andrews DW et al, Glioma grade is associated with the accumulation and activity of cells bearing M2 monocyte markers. Clin Cancer Res 2013;19:3776–3786.
Nguyen N, Bellile E, Thomas D et al, Tumor infiltrating lymphocytes and survival in patients with head and neck squamous cell carcinoma. Head Neck 2016;38:1074–1084.
Kim JH, Oh SH, Kim EJ et al, The role of myofibroblasts in upregulation of S100A8 and S100A9 and the differentiation of myeloid cells in the colorectal cancer microenvironment. Biochem Biophys Res Commun 2012;423:60–66.
Mazur G, Haloń A, Wróbel T et al, Macrophage/histiocytic antigen CD68 expression in neoplastic and reactive lymph nodes. Rocz Akad Med Bialymst 2004;49 (Suppl 1):73–75.
Strojnik T, Kavalar R, Zajc I et al, Prognostic impact of CD68 and kallikrein 6 in human glioma. Anticancer Res 2009;29:3269–3279.
Ni YH, Ding L, Huang XF et al, Microlocalization of CD68+ tumor-associated macrophages in tumor stroma correlated with poor clinical outcomes in oral squamous cell carcinoma patients. Tumour Biol 2015;36:5291–5298.
Tzankov A, Matter MS, Dirnhofer S . Refined prognostic role of CD68-positive tumor macrophages in the context of the cellular micromilieu of classical Hodgkin lymphoma. Pathobiology 2010;77:301–308.
Kong LQ, Zhu XD, Xu HX et al, The clinical significance of the CD163+ and CD68+ macrophages in patients with hepatocellular carcinoma. PLoS One 2013;8:e59771.
Sjödahl G, Lövgren K, Lauss M et al, Infiltration of CD3+ and CD68+ cells in bladder cancer is subtype specific and affects the outcome of patients with muscle-invasive tumors. Urol Oncol 2014;32:791–797.
Coiffier B, Li W, Henitz ED et al, Prespecified candidate biomarkers identify follicular lymphoma patients who achieved longer progression-free survival with bortezomib-rituximab versus rituximab. Clin Cancer Res 2013;19:2551–2561.
Steidl C, Farinha P, Gascoyne RD . Macrophages predict treatment outcome in Hodgkin's lymphoma. Haematologica 2011;96:186–189.
Agostinelli C, Gallamini A, Rigacci L et al, The prognostic value of biologic markers in classical Hodgkin lymphoma (cHL) patients. Ann Oncol 2011;22:153–154.
Azambuja D, Natkunam Y, Biasoli I et al, Lack of association of tumor-associated macrophages with clinical outcome in patients with classical Hodgkin's lymphoma. Ann Oncol 2012;23:736–742.
Harris JA, Jain S, Ren Q et al, CD163 versus CD68 in tumor associated macrophages of classical Hodgkin lymphoma. Diagn Pathol 2012;7:12.
Klein JL, Nguyen TT, Bien-Willner GA et al, CD163 immunohistochemistry is superior to CD68 in predicting outcome in classical Hodgkin lymphoma. Am J Clin Pathol 2014;141:381–387.
Acknowledgements
This work was supported by the Russian Science Foundation (grant #14-15-00112), Russian Federation.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
CD68 is not restricted to serving as a cytochemical marker of monocyte/macrophages. CD68 is involved in binding several ligands, such as oxidized low density lipoproteins, phosphatidylserine, apoptotic cells and malaria sporozoite. However, CD68 it is not involved in binding bacterial/viral pathogens, innate, inflammatory or humoral immune responses.
Rights and permissions
About this article

Cite this article
Chistiakov, D., Killingsworth, M., Myasoedova, V. et al. CD68/macrosialin: not just a histochemical marker. Lab Invest 97, 4–13 (2017). https://doi.org/10.1038/labinvest.2016.116
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/labinvest.2016.116
This article is cited by
-
Dendritic cells under allergic condition enhance the activation of pruritogen-responsive neurons via inducing itch receptors in a co-culture study
BMC Immunology (2024)
-
Nanotechnology for microglial targeting and inhibition of neuroinflammation underlying Alzheimer’s pathology
Translational Neurodegeneration (2024)
-
TYROBP/DAP12 knockout in Huntington’s disease Q175 mice cell-autonomously decreases microglial expression of disease-associated genes and non-cell-autonomously mitigates astrogliosis and motor deterioration
Journal of Neuroinflammation (2024)
-
Single-cell RNA sequencing analysis of vestibular schwannoma reveals functionally distinct macrophage subsets
British Journal of Cancer (2024)
-
Identification of a leucine-mediated threshold effect governing macrophage mTOR signalling and cardiovascular risk
Nature Metabolism (2024)
